Симбиоз — основа растительной жизни
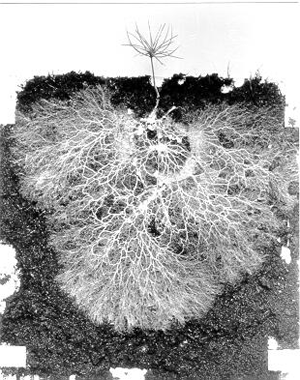
Высшие растения жили в симбиозе с грибами и бактериями в течение всей своей истории. Выход растений на сушу во многом был обусловлен симбиозом с грибами (микориза). Генетические системы, обеспечивающие взаимодействие растений с микоризными грибами, в дальнейшем многократно менялись в связи с вовлечением в симбиоз новых грибов и бактерий. Первичная функция корней состояла в обеспечении симбиоза с почвенными микроорганизмами, и лишь потом корни приобрели способность самостоятельно усваивать питательные вещества из почвы.
Статья Н. А. Проворова представляет собой большой аналитический обзор, содержащий так много важных фактов и идей, что его переложение в виде краткой популярной заметки представляется делом довольно неблагодарным. Однако это необходимо сделать, поскольку мы не можем публиковать полные тексты статей (напомним, что организация, именуемая «МАИК Наука — Интерпериодика», владеет монопольным правом на торговлю этими текстами).
Ранее на нашем сайте уже был опубликован пересказ статьи Н. А. Проворова и Е. А. Долгих (см.: От биохимического сотрудничества — к общему геному; там же есть подборка ссылок по новейшим открытиям в области изучения симбиотических систем).
В своей новой статье Н. А. Проворов показывает, что, несмотря на огромное разнообразие растительно-микробных симбиозов, большинство из них, по-видимому, имеют единое эволюционное происхождение.
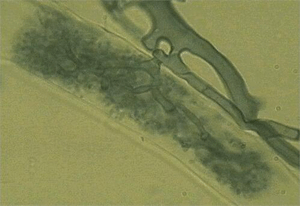
К концу XX века стало ясно, что внешне различные формы растительно-микробных симбиозов основаны на очень сходных генетических, клеточных и молекулярных механизмах. Их изучение привело большинство специалистов к выводу о том, что все наблюдаемое многообразие симбиозов растений с почвенными грибами и бактериями произошло от одной самой древней, первичной формы такого симбиоза — арбускулярной микоризы (АМ). Грибы, участвующие в АМ, проникают внутрь растительных клеток, образуя там особые внутриклеточные струкутуры — арбускулы (см.: Изменение гена, необходимого для симбиоза растений с грибами, привело к формированию симбиоза с азотфиксирующими бактериями. «Элементы», 12.03.08).
1. Происхождение арбускулярной микоризы и происхождение наземных растений. Уже самые древние и примитивные наземные растения — псилофиты — жили в симбиозе с грибами и имели АМ (см.: W. Remy, T. N. Taylor, H. Hass, H. Kerp. Four hundred-million-year-old vesicular arbuscular mycorrhizae // PNAS. 1994. V. 91. P. 11841–11843). У псилофитов еще не было настоящих корней. Их подземную часть представляли ризоиды, которые могли служить для закрепления растения в грунте, но не для питания. Поэтому для первых наземных растений симбиоз с грибами, по-видимому, был абсолютно необходим. АМ характерна и для большинства современных растений (а те, у которых ее нет, скорее всего, происходят от предков, имевших АМ).
На этом основании еще в 1970-х годах была предложена гипотеза о том, что выход растений на сушу состоялся именно благодаря симбиозу с АМ-грибами (Pirozinski, Malloch, 1975). Эта гипотеза впоследствии блестяще подтвердилась не только палеонтологическими данными, но и молекулярно-филогенетическими: анализ генов 18S рРНК показал, что АМ-грибы происходят от общего предка, жившего 400–00 млн лет назад, т.е. как раз в то время, когда на суше появились первые растения.
По-видимому, «уже на заре эволюции наземных растений у них сложилась способность регулировать жизнедеятельность микроорганизмов, колонизирующих подземные органы». Генные системы АМ довольно универсальны (это подверждается низкой специфичностью АМ-грибов по отношению к растениям), и впоследствии они многократно перестраивались для организации различных симбиозов в различных группах растений.
Как растения, так и грибы, по-видимому, могли «подготовиться» к долгой совместной жизни задолго до выхода растений на сушу. Возможно, предки высших растений уже в водной среде вступали в симбиозы с различными водными грибами, как это делают сегодня зеленые и красные водоросли. Грибы, вышедшие на сушу намного раньше растений, могли вступать здесь в симбиоз с цианобактериями. Гриб Geosiphon, считающийся наиболее вероятным предком АМ-грибов, вступает в симбиоз с цианобактериями Nostoc, которые не только фотосинтезируют, но и фиксируют атмосферный азот. Это позволяет грибу жить на крайне бедных субстратах. Симбиозы такого типа могли быть широко распространены на суше до ее освоения растениями. Таким образом, еще до выхода растений на сушу почвенные грибы могли выработать эффективные системы для усвоения органики, производимой фотосинтезирующими симбионтами, а также для снабжения этих симбионтов фосфатами, поглощаемыми грибом из почвы.
В отличие от цианобактерий, растения могут снабжать симбиотические грибы гораздо большим количеством органики. Недостаток азота в симбиотической системе мог быть компенсирован симбиозом АМ-грибов с другими азотфиксирующими микробами.
2. Для чего нужны корни? Н. А. Проворов предполагает, что «поддержание микоризных грибов могло быть более древней функцией корней, чем самостоятельное усвоение питательных веществ из почвы». В дальнейшем корни «научились» самостоятельно всасывать минеральные вещества из почвы, и зависимость от микоризных грибов у некоторых растений стала слабее. Часть цветковых травянистых растений, а также некоторые культурные растения вообще утратили микоризу (у последних это во многом было связано с переходом растений на питание минеральными удобрениями). Однако большинство растений так и не перешли к полностью самостоятельному корневому питанию, а некоторые орхидные вообще утратили фотосинтез и стали получать весь углерод от симбиотических грибов. Иные из этих удивительных растений (орхидея Galeola) достигают гигантских размеров и имеют широкие ареалы. Эти орхидеи фактически паразитируют на других растительно-грибных симбиозах (грибы, от которых орхидеи получают органику, сами получают ее от других растений). Таким образом, «в ходе коэволюции растений с грибами потоки питательных веществ, которыми они обмениваются, могли достаточно легко менять свое направление». Растения, утратившие фотосинтез и перешедшие к паразитизму на грибах — симбионтах других растений известны и за пределами семейства орхидных (они есть среди двудольных, папоротников, мхов и печеночников).
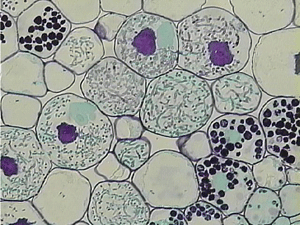
3. Эволюция грибов — симбионтов растений. Симбиотические отношения растений с грибами очень разнообразны и вовсе не исчерпываются арбускулярной микоризой. Раньше считалось, что многие грибы, находящиеся с растениями в мутуалистических (взаимовыгодных) отношениях в прошлом были паразитами. Однако современные данные, в том числе молекулярно-филогенетические реконструкции, не подтверждают этого. По-видимому, переход от паразитизма к мутуализму был явлением весьма редким. Гораздо чаще симбионтами растений становились грибы-сапрофиты (питающиеся мертвой органикой). Многократно происходили и обратные переходы от мутуализма с растениями к свободному существованию в качестве сапрофитов. Многие адаптации, связанные с мутуализмом, возможно, вырабатывались независимо (конвергентно) у разных грибов-сапрофитов при переходе к мутуализму с растениями. Однако «эволюцию симбиотических признаков в разных группах высших грибов нельзя считать полностью независимой, поскольку такие их фундаментальные свойства, как образование мицелия и осмотрофное питание, могли возникать в процессе адаптации к развитию в тканях растений». Иными словами, вполне возможно, что эволюция основных групп высших грибов (аскомицетов и базидиомицетов) с самого начала была теснейшим образом связана с растениями. «По некоторым данным, до 90% аскомицетов и базидиомицетов имеют в своих жизненных циклах стадии симбиозов с растениями, образование которых можно считать одним из древнейших свойств мицелиальных грибов». По-видимому, не только наземные растения, но и основные группы грибов сумели стать столь разнообразными и процветающими в первую очередь благодаря симбиозу и коэволюции.
4. Симбиоз с азотфиксирующими бактериями. Генетические системы, обеспечивающие возможность симбиоза растений с азотфиксирующими бактериями, по-видимому, являются результатом модификации генетических систем АМ (см.: Изменение гена, необходимого для симбиоза растений с грибами, привело к формированию симбиоза с азотфиксирующими бактериями. «Элементы», 12.03.08). Кроме того, Н. А. Проворов предполагает, что «в ходе эволюции АМ гломусовые грибы служили для растений донорами азотфиксирующих симбионтов». Гломусовые грибы, участвующие в АМ, часто вступают в симбиоз с азотфиксирующими бактериями. «Учитывая поистине планетарный масштаб происходящего при развитии АМ смешивания грибной и растительной цитоплазмы, логично предположить, что отбор мог подхватывать даже очень редко возникающие эндосимбионты грибов, способные сохранять жизнеспособность в цитоплазме растений».
В статье подробно разбираются возможные пути эволюции симбиоза растений с азотфиксирующими бактериями — ризобиями и актинобактериями. Отмечается, что большую роль в развитии этих сибмиозов сыграли преадаптации, развившиеся в ходе эволюции АМ, однако для налаживания взаимоотношений с новыми симбионтами были рекрутированы и многие гены и генные комплексы растений, которые ранее выполняли другие функции.
По-видимому, «приобретение двудольными способности к азотфиксирующим клубеньковым симбиозам было связано с последовательным замещением различных типов микроорганизмов, которые могут заселять межклеточные и субклеточные симбиотические компартменты в кортексе корня. При этом прокариотические азотфиксаторы использовали анцестральную программу размещения («хостинга») микросимбионтов, которая возникла при коэволюции древнейших наземных растений с АМ-грибами и претерпевала закономерные усложнения, происходившие параллельно в различных семействах».
Начальным этапом этого процесса, возможно, было замещение АМ-грибов азотфиксирующими актинобактериями Frankia. Эти бактерии внешне очень похожи на грибы (поэтому их раньше называли актиномицетами). Как и грибы, они образуют мицелий. Поначалу азотфиксирующая активность новых симбионтов была низкой, но потом растения выработали средства для ее интенсификации (в частности, более интенсивно стали откачиваться в надземную часть растения азотистые соединения, которые ингибируют азотфиксацию).
Симбиоз с Frankia создал предпосылки для вступления растений в симбиоз и с другими азотфиксаторами, которые могли вытеснять актинобактерий из программы развития симбиоза, в частности, благодаря своему более быстрому росту. Бактерии-конкуренты научились быстрее, чем Frankia, активировать у растений программу «хостинга», т.е. стимулировать растение к приему симбионтов. Проникновению Frankia в корень предшествует долгий (в несколько суток) период накопления актинобактерий у поверхности корней, тогда как ризобии проходят этот предварительный этап всего за несколько часов.
Однако замена «грибоподобных» актинобактерий другими бактериальными симбионтами (ризобиями) была сопряжена с опасностью, поскольку это открывало дорогу в организм растения множеству других бактерий, в том числе патогенных. Возможно, именно поэтому симбиоз с ризобиями сложился лишь у бобовых и некоторых вязовых (это могло быть связано с особенностями защитных систем этих растений).
Замещение микоризных грибов азотфиксаторами вовсе не означало отказ от микоризы. Напротив, у подавляющего большинства бобовых и «актиноризных» растений имееется также и АМ. При этом азотфиксирующие симбионты снабжают растение азотом, а грибы — фосфором. Однако растения, живущие в симбиозе с ризобиями, не образуют актиноризу, потому что нет смысла поддерживать два дублирующих друг друга азотфиксирующих симбиоза (на их поддержание растению приходится тратить много энергии).
Таким образом, развитие разнообразных растительно-грибных и растительно-бактериальных симбиозов представляет собой единый эволюционный континуум.
См. также:
От биохимического сотрудничества — к общему геному. По статье: Н. А. Проворов, Е. А. Долгих Метаболическая интеграция организмов в системах симбиоза // Журнал общей биологии. 2006. Т. 67. С. 403–422.
Изменение гена, необходимого для симбиоза растений с грибами, привело к формированию симбиоза с азотфиксирующими бактериями. «Элементы», 12.03.08.
Remy et al., 1994. Four hundred-million-year-old vesicular arbuscular mycorrhizae.
Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Популярные синопсисы