Происхождение цветковых растений остается загадочным

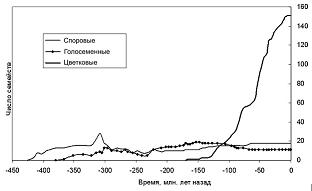
Цветковые, или покрытосеменные – сравнительно молодая группа высших растений, появившаяся в меловом периоде, стремительно размножившаяся в середине – конце мела и с тех пор занимающая господствующее положение на суше. Проблеме происхождения цветковых посвящено огромное количество публикаций, однако в этом вопросе по-прежнему много неясного. Общепризнано, что цветковые произошли от голосеменных, но от кого именно и каким образом?
Д.Д.Соколов и А.К.Тимонин с кафедры высших растений биофака МГУ подробно рассмотрели основные теории происхождения покрытосеменных в обзорной статье, опубликованной во втором номере Журнала общей биологии за 2007 г.
Авторы обращают внимание на парадоксальный факт: цветы покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Поэтому и сформулировать, что такое цветок, весьма трудно. Тем не менее ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных.
Впрочем, если не рассматривать мужские цветки, у остальных все-таки есть общая черта, отличающая цветок покрытосеменных от фруктификаций голосеменных. У цветковых плодолистики свернуты и образуют, во-первых, рыльце, улавливающее пыльцу, во вторых – замкнутую полость (завязь), в которой развиваются семязачатки. У некоторых голосеменных есть нечто похожее – замкнутые капсулы с семязачатками, однако у них нет завязи (пыльцу улавливают сами семязачатки при помощи длинных отростков, высовывающихся из узкого отверстия капсулы). К тому же исходный листочек, несущий семязачатки (мегаспорофилл, у цветковых ему соответствует плодолистик) свернут у голосеменных нижней (абаксиальной) стороной внутрь, а у цветковых – наоборот, верхней (адаксиальной) стороной внутрь. Соответственно и семязачатки сидят на разных сторонах листочка.
Эти различия представляются настолько радикальными, что большинство ботаников не видят возможности постепенного перехода из одного состояния в другое и предполагают сальтационное превращение (одним скачком). Это вполне возможно, если учесть, что развитие фруктификаций у растений управляется небольшим числом генов-регуляторов, мутации в которых действительно могут приводить к скачкообразным изменениям плана строения. Аналогичная ситуация наблюдается и у животных, план строения которых в основном определяется небольшим набором генов-регуляторов (HOX-гены; см.: «Новое в науке о знаменитых Hox-генах, регуляторах развития»).
В начале XX века была популярна теория происхождения цветковых от беннеттитов – вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) – в отличие от других голосеменных, имеющих однополые фруктификации. Из стробилов беннеттитов, действительно напоминающих цветы, ботаники и пытались вывести типичный для покрытосеменных обоеполый цветок. Но тут возникла проблема: у беннеттитов не обнаружилось никаких структур, из которых можно было бы вывести плодолистики. У них вообще нет мегаспорофиллов, а семязачатки сидят прямо на оси стробила. Проблема казалась неразрешимой.
В 80-х годах XX века крупнейший российский палеоботаник С.В.Мейен выдвинул гипотезу, разрешающую это противоречие. Он предположил, что у каких-то беннеттитовых сальтационным путем произошла перестройка программы развития фруктификации. В результате ключевые элементы программы развития мужских элементов стробилы (микроспорофиллов, у цветковых это тычинки) стали реализовываться при развитии женских элементов. Это привело к появлению химерных образований – микроспорофиллов, несущих семязачатки вместо пыльников1 . Именно из таких «мутантных» микроспорофиллов и образовались плодолистики. У беннеттитовых микроспорофиллы несут пыльники как раз на своей верхней стороне, в отличие от других голосеменных. Поэтому такая мутация сразу привела к появлению «правильных» прототипов для формирования плодолистиков, с семязачатками на нужной стороне.
Ничего невероятного в таком сценарии нет, исходя из того, что известно об общих принципах генетической регуляции развития. Гипотеза Мейена исключительно изящно и просто объясняет противоречие, казавшееся неразрешимым. Ее нельзя проверить классическими методами палеоботаники и морфологии (поскольку никаких переходных форм гипотеза Мейена не предполагает). Однако ее можно проверить методами сравнительно-генетического анализа.
Генетическая регуляция развития фруктификаций у семенных растений стала активно изучаться в начале 90-х годов XX века. Оказалось, что у всех цветковых главным регулятором развития цветка является ген LFY. У голосеменных есть аналогичный ген (Leaf family), который экпрессируется (работает) на ранних стадиях развития мужских фруктификаций. Кроме того, у голосеменных есть еще один сходный ген (Needle family), отсутствующий у цветковых, который участвует в формировании женских фруктификаций. У большинства голосеменных, напомним, фруктификации раздельнополы. При появлении цветковых, таким образом, «женский ген» был утрачен, а «мужской» стал руководить развитием всего цветка, с его мужскими и женскими частями.
На этом основании Фролик и Паркер (Frolich, Parker, 2000) выдвинули «преимущественно мужскую теорию» происхождения цветка. Они предположили, что в результате утраты гена из семейства Needle у предков цветковых женские фруктификации перестали формироваться, а семязачатки стали развиваться на верхней стороне плоских микроспорофиллов. Казалось бы, это в общем то же самое, что предполагал С.В.Мейен. Авторы «преимущественно мужской теории», однако, подчеркивают, что Мейен предполагал перенос признаков мужских структур на женские, а они говорят о переносе органов с места на место (семязачатков на микроспорофиллы). Но это, конечно, больше похоже на игру в слова, а не на существенное различие двух теорий. Различие на самом деле в другом, а именно в вопросе о предковой группе цветковых. От кого они все-таки произошли – от беннеттитов с обоеполыми фруктификациями, или от других голосеменных с разнополыми?
Эволюционные реконструкции, основанные на морфологии, подтверждали родство покрытосеменных и беннеттитовых (а также родственных беннеттитам гнетовых, которые дожили до наших дней). Однако реконструкции, основанные на молекулярных данных, показали иное. Они показали, что все дожившие до современности голосеменные представляют собой монофилетичную группу (т.е. группу, происходящую от одного общего предка и включающую всех без исключения потомков этого предка). Это значит, что последний общий предок всех современных голосеменных, в том числе гнетовых (а стало быть и беннеттитов тоже) не был предком покрытосеменных. Таким образом, предка цветковых нужно искать среди других вымерших голосеменных, более древних, точнее, обособившихся раньше, чем беннеттитовые. Фролик и Паркер предложили свою кандидатуру – користоспермовые семенные папоротники (Corystospermataceae). У них, правда, микроспорангии (пыльники) располагались на нижней, а не на верхней стороне микроспорофиллов.
Д.Д.Соколов и А.К.Тимонин также приводят доводы в пользу того, что строение семязачатков користоспермовых не позволяет вывести из них семязачатки покрытосеменных. «Подходящие» семязачатки есть у другой группы голосеменных – у кейтониевых, но зато у них нет ничего похожего на плоские микроспорофиллы, из которых можно было бы вывести плодолистики.
Авторы предполагают, что причина всех этих противоречий, вероятно, кроется в недостаточной надежности эволюционных реконструкций, основанных на молекулярных данных. По их мнению, голосеменные растения плохо подходят для молекулярно-филогенетического анализа, поскольку большинство семейств голосеменных давно вымерло. Мы, таким образом, имеем недостаточно репрезентативную выборку таксонов, что, как хорошо известно, может приводить к ошибочным молекулярным филогениям. В конечном счете, по мнению авторов, на сегодняшний день нет оснований для безоговорочного отрицания идеи Мейена о происхождении цветковых от беннеттитовых. Эту идею вполне можно сохранить в качестве рабочей гипотезы.
1Да простят меня ботаники за нестрогость терминологии, но упоминать про «синангии микроспорангиев» в популярной заметке было бы жестоко по отношению к читателю :)
Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Популярные синопсисы