Мобильные генетические элементы – один из факторов, делающих нас людьми

Поиск генетических отличий человека от его ближайших родственников – шимпанзе – является одной из интереснейших задач современной биологии. На настоящий момент представляется совершенно очевидным, что мобильные генетические элементы отнюдь не являются ненужным «мусором» генома на фоне «важных» белоккодирующих последовательностей. Напротив, показано, что мобильные генетические элементы, радикальным образом влияя на работу генов, играют одну из ключевых ролей в эволюции организмов, в том числе, и на эволюцию человека. Подробный анализ специфических для человека мобильные генетические элементы, возможно, позволит выявить множество генов, имеющих уникальный для человека характер экспрессии. Генов, отличающих, таким образом, человека от животных.
Что отличает нас от животных? Поэт, романтик и философ скажут, что человека от животных отличает наличие души. Специалист в области высшей нервной деятельности вслед за Павловым заметит, что только у человека есть вторая сигнальная система. Нейробиолог добавит, рассказав нам много об особенностях строения мозга человека: и про увеличение ассоциативных полей неокортекса, и про увеличение височных долей, про центры речи и про многое другое. Антрополог, изучающий кости ископаемых предков Homo sapiens, обязательно скажет, что только у нас есть подбородочный выступ в черепе, нет надбровных дуг, несоразмерно большой мозговой отдел черепа и т.д. (Существует даже красивая теория, что одно из отличий человека от животных – это умение смеяться и плакать.) И все они будут правы. Выходит, искать отличия человека от животных можно в самых разных областях науки, да и жизни вообще. Однако, поскольку все признаки организма определяются его геномом, все эти отличия можно свести к отличиям в геноме. И поиск генетической основы, ответственной за очевидные различия в фенотипах человека и животных, особенно, его ближайших родственников, представляется одной из интереснейших задач современной биологии. Показано, что значительный вклад в структуру и функционирование генома, и, соответственно, в его видоспецифичность, вносят мобильные генетические элементы (МГЭ).
В обсуждаемой статье, посвященной роли МГЭ в функционировании генома человека, К. К. Баскаев и А. А. Буздин из Института биоорганической химии им. акад. М. М. Шемякина и Ю. А. Овчинникова РАН приводят краткий обзор основных генетических отличий человека от его ближайших нынеживущих родственников – шимпанзе, представленных в настоящее время двумя видами: обыкновенный шимпанзе (Pan troglodytes) и карликовый шимпанзе, или бонобо (P. paniscus). Авторы выделяют четыре основные группы генетических различий человека от шимпанзе.
1. Различная хромосомная организация, включающая утраты частей хромосом (делеции), различные вставки (инсерции), удвоения частей хромосом (дупликации) и обмен участками между негомологичными хромосомами (транслокации). Таких хромосомные перестройки происходят сравнительно часто, они имели место и на филогенетической линии, ведущей к человеку, и на линии шимпанзе. Посчитано, что инсерции и делеции, по которым человек отличается от шимпанзе, включают около 150 млн. п.о. Гораздо реже происходят более крупномасштабные события, такие как изменения числа хромосом. На человеческой линии произошло объединение двух предковых хромосом с образованием человеческой 2 хромосомы. У шимпанзе эта хромосома соответствует 12 и 13 хромосомам. Еще одно существенное отличие – перестройки в теломерных и центромерных областях, а так же в некоторых участках Y-хромосомы
2. Вариации в числе копий, положении в геноме и активности общих для человека и шимпанзе последовательностей.
3. Различия в белоккодирующих регионах, результатом которых являются различия в первичной структуре некоторых белков.
4. Видоспецифичные инсерции МГЭ.
Различия между ДНК человека и шимпанзе составляют в среднем около 1,24% и всего примерно 0,5% в белоккодирующих участках. Это значит, что если из всего генома человека наугад выбрать 100 нуклеотидов, то из них, скорее всего, только 1,24 будут не такими, как у шимпанзе. Тем не менее, никто не спутает человека с шимпанзе. Дело в том, что геном организован так, что один маленький эффект – экспрессия белоккодирующего гена - регулируется сложнейшей огромной сетью регуляторных генов. И маленькое изменение в этой пирамиде может радикальным образом увеличить или уменьшить экспрессию какого-то белка.
Особое внимание в обзоре было уделено анализу вклада специфических для человека МГЭ в структуру и функционирование генома человека.
Напомню о принципе строения, функционирования и вкладе МГЭ в геном. Тем, кто все это знает, следующие три абзаца можно пропустить. МГЭ – это фрагмент двунитевой молекулы ДНК, часть генома организма-хозяина, способная к самовоспроизведению, независимо от воспроизведения остального генома.

Этот белок умеет «вырезать» МГЭ из молекулы ДНК, делать ступенчатые разрывы в сайте встраивания в молекуле ДНК-мишени и вшивать вырезанный МГЭ в «ступенчато разорванный» сайт встраивания. После вшивания, ДНК-полимераза и ДНК-лигаза заполняют появившиеся в результате ступенчатых разрывов «липкие концы» ДНК, и образуются концевые прямые повторы - отличительный признак МГЭ, уже прошедшего транспозицию (рис. 3).
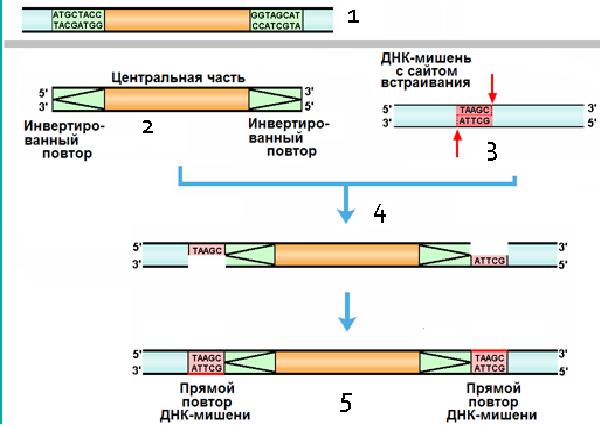
У эукариот, помимо описанных выше IS-элементов, существует два класса транспозонов: транспозоны I класса, присущие только эукариотам, и транспозоны II класса, общие для прокариот и эукариот. Транспозоны II класса, или ДНК-транспозоны, отличаются от описанных IS-элементов лишь тем, что они включает не только ген транспозазы, но и другие, посторонние, гены. Бывают составные ДНК-транспозоны и несоставные, или комплексные, ДНК-транспозоны. Составные ДНК-транспозоны представляют собой два идентичных типичных IS-элемента, окружающих центральную часть, содержащую посторонние гены, и вся эта конструкция вырезается, переносится и вшивается как единое целое. Комплексные ДНК-транспозоны по сути являются IS-элементами, отличие от типичных IS-элементов в том, что у этих транспозонов в центральной части присутствуют посторонние гены. Транспозоны II класса, или ретротранспозоны, присущи только эукариотам. И это, наряду с эндогенными ретровирусами (см. ниже), единственный способный к транспозиции класс МГЭ, найденный у млекопитающих и человека. МГЭ других классов утратили способность автономно воспроизводиться и перемещаться по геному из-за накопившихся мутаций. Главное отличие ретротранспозонов от ДНК-транспозонов в том, что их цикл репродукции включает стадию промежуточной молекулы РНК и стадию обратной транскрипции. Соответственно, в составе ретротранспозонов, как правило, имеется участок, кодирующий обратную транскриптазу, или ревертазу. На ретротранспозоны очень похожи т.н. эндогенные ретровирусы эукариот – ретровирусы, прочно интегрированные в геном клетки-хозяина и утратившие способность к образованию вирионов. Т.е. эндогенные ретровирусы можно рассматривать как МГЭ, имеющие вирусное происхождение, но на настоящей стадии являющиеся в большей степени частью организма-хозяина, чем частью вирусного паразита.
Результатом вставок МГЭ является отключение генов в результате непосредственной вставки МГЭ в их белоккодирующие части и изменение их активности в результате вставки МГЭ в регуляторные участки. Так, например, у человека не работает ген CMP, кодирующий фермент гидроксилазу сиаловой кислоты. Это фермент превращает N-ацетилнейраминовую кислоту в N-гликолилнейраминовую кислоту, которую человек, в отличие от шимпанзе, синтезировать не умеет. Причина отключения гена – вставка в его белоккодирующую часть специфического для человека ретротранспозона из семейства Alu. Другой результат транспозиции – это рекомбинация генома (рис. 4).

Итак, вернемся к специфическим для человека вставкам МГЭ. Как писалось выше, геном человека отличается от генома шимпанзе на 1,24%. Из этих отличий 78% приходится на МГЭ-независимые делеции и дупликации, 19% - на простые замены и лишь 3% - на вставки ретроэлементов – единственных активных (не выключенных накопившимися мутациями) МГЭ человека. Авторы обсуждаемой статьи выделяют четыре группы «человеческих» ретроэлементов: HERV-K (от Human endogenous retroviruses, эндогенные ретровирусы человека), L1, Alu и SVA, на которые приходится, соответственно, 5%, 51%, 24% и 20% от всех «человеческих» ретроэлементов. В обзоре перечислены основные известные на сегодняшний день виды влияний специфических для человека МГЭ на функционирование генома человека.
1. МГЭ – субстраты рекомбинации. Основные механизмы МГЭ-зависимой рекомбинации приведены на рис. 4.
2. МГЭ – энхансеры транскрипции. Действительно, известно много случаев, когда МГЭ увеличивает уровень экспрессии гена. Так, например, ретроэлемент семейства Alu является частью энхансерной области (т.е. области, повышающей активность) человеческого гена CD8.
3. МГЭ – транскрипционные промоторы. Оказывается, около 25% человеческих промоторов [0] (сайтов связывания ДНК-зависимой РНК-полимеразы) содержат в своей последовательности ретроэлементы.
4. МГЭ – поставщики альтернативных сайтов сплайсинга. Здесь ключевая роль принадлежит ретротранспозонам Alu. По краям белоккодирующего участка гена всегда есть нетранслируемые участки, в которых присутствие Alu может оказывать существенное влияние на то, что происходит с мРНК после транскрипции. Механизмы этого влияния могут быть разными: влияние на трансляцию, стабильность мРНК и альтернативный сплайсинг. Т.е. МГЭ могут влиять на продукт гена уже после его транскрипции. Примером такого влияния может служить специфическая для человека вставка Alu в ген, связанный с врождённой мышечной дистрофией.
5. МГЭ – доноры сигнала полиаденилирования. Напомню, что полиаденилирование – один из важных этапов созревания мРНК перед трансляцией, состоящий в присоединении 100-200 остатков аденина к определенной сигнальной последовательности – сигналу полиаденилирования. МГЭ человека кодируют свои гены, и эти гены имеют свои сигналы полиаденилирования. Значит, включение МГЭ в гены человека создает альтернативные сайты полиаденилирования.
6. МГЭ – антисмысловые регуляторы транскрипции . Антисмысловые РНК – это РНК, комплементарные мРНК. Показано, что МГЭ, присутствующие в интронах генов, как правило, находятся в антисмысловой ориентации относительно направления транскрипции этого гена. Таким образом, промоторы МГЭ могут управлять транскрипцией РНК, комплементарной участкам мРНК соответствующего гена. Получившаяся антисмысловая РНК, комплементарно связываясь с мРНК, может подавлять ее сплайсинг и трансляцию.
Не будет лишним еще раз отметить, что все перечисленные механизмы влияния МГЭ обнаружены у человека. И для каждого механизма есть примеры, отсутствующие у шимпанзе. Таким образом, по мнению авторов обсуждаемой статьи, специфические для человека МГЭ можно рассматривать в качестве важного кандидата на роль агента антропогенеза. Если выйти за рамки эволюции человека и подойти к проблеме влияния МГЭ на эволюцию в целом, то можно увидеть, что это влияние неожиданно велико. Показано, что МГЭ играли ключевую роль в эволюции млекопитающих («Прочтение генома опоссума доказало ключевую роль транспозонов в эволюции млекопитающих»). Учитывая, что на настоящий момент детально изучено лишь 2% всех специфических для человека МГЭ, можно надеяться, пишут авторы, что дальнейший полногеномный анализ МГЭ человека позволит выявить множество генов, имеющих уникальный для человека характер экспрессии. Возможно, это позволит нам приблизиться к извечному вопросу человечества как группы организмов, способных к научному изучению самих себя: «Что же делает нас людьми?».
Последние выпуски
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
- Том 86, № 4. Июль-август 2025
- Том 86, № 3. Май-июнь 2025
Популярные синопсисы