О том, как встреча после разлуки завершает видообразование

Между популяциями, произошедшими от общего предка и развивающимися далее в географической изоляции, накапливаются различия. Эти различия могут стать причиной генетической несовместимости при последующей встрече популяций: их гибридное потомство будет слабым и хилым, если вообще сможет появиться на свет. Чтобы отличить «своих» от «чужих», избежать пустой траты времени и энергии на взаимные ухаживания и спаривания, бабочки голубянки вновь сошедшихся популяций вынуждены были ускоренно обзавестись различной внешностью – они изменили цвет крыльев. Мы наблюдаем ситуацию, в которой выгодно усиление репродуктивной изоляции, оно является целью, а не побочным продуктом дивергенции.
Произошедшие от одного предка сестринские популяции, изолированные друг от друга географически, неизбежно приобретают специфические особенности, связанные с генетическими изменениями. Популяции накапливают эти генетические перестройки или мутации, не проверяя их на взаимную совместимость. В результате накопленные мутации одной популяции, вполне мирно уживающиеся с ее старыми генами, могут оказаться слегка вредоносными или абсолютно несовместимыми в сочетании с новыми мутациями популяции сестринской (см.: Генетическая несовместимость нарастает по параболе. «Элементы», 26.09.10). При последующем контакте этих популяций (коли таковой происходит) они либо снова сольются воедино, либо продолжат существовать самостоятельно как два отдельных вида (впрочем, одна из них, более успешная, вполне может вытеснить другую). Встает вопрос: если разграничение на виды все-таки происходит, то в какой именно момент?
Считается, что возможны два сценария становления репродуктивной изоляции между такими встретившимися популяциями (см.: Видообразование). В первом случае предполагается, что уже в аллопатрии, при раздельном проживании, предрешено, быть или не быть им далее единым видом. Репродуктивная изоляция возникает сама по себе, как неизбежный побочный продукт дивергенции, как прямое естественное следствие географического разобщения – и только.
Второй случай предполагает, что встретившиеся особи еще не окончательно утратили способность к скрещиванию. Однако если их гибриды вследствие генетической несовместимости будут иметь пониженную приспособленность или окажутся стерильными, то выиграют те особи, которые сумеют избежать скрещивания с «чужаками». Любые мутации, приводящие к увеличению различий между представителями двух популяций и заставляющие особей предпочитать в виде половых партнеров своих соплеменников, получат преимущество. Произойдет «усиление» (reinforcement) механизмов репродуктивной изоляции, в результате которого представители разных популяций вовсе перестанут скрещиваться друг с другом. В этом случае отбор «целенаправленно» создает репродуктивную изоляцию ради нее самой – и видообразование завершается.
Селективную выгоду изолирующих механизмов, направленных на предотвращение гибридизации, первым отметил А. Уоллес в 1889 году. В 1937-40 гг. интерес к эффекту усиления, как к одному из путей видообразования, возродил Ф.Г. Добржанский.
Выигрыш от презиготической, т.е. предотвращающей оплодотворение, изоляции в случае пониженной жизнеспособности гибридов – очевиден. Экспериментально неоднократно было продемонстрировано формирование сексуальных предпочтений особей в сторону себе подобных после периода изоляции, сопряженного с изменениями условий обитания (см.: Опыты по «искусственному видообразованию…»). Однако тот факт, что отбор, направленный на изоляцию, не представляет собой универсального процесса, а также трудность в определении времени образования изолирующих морфологических и поведенческих признаков – возникли они еще в аллопатрии, или уже в симпатрии – долгое время не позволяли обнаружить бесспорных эмпирических подтверждений такому отбору.
В.А. Лухтанов из СПбГУ противопоставляет две гипотезы: аллопатрического видообразования, как единственно возможного (поддержанную Аланом Темплтоном) и «усиления» видообразования при вторичной симпатрии (названную автором правилом Добржанского). В обсуждаемой статье логично и последовательно доказывается реальность второй модели на примере эволюционно молодого рода бабочек голубянок (Agrodiaetus).

Самки голубянок разных видов имеют примерно одинаковый темно-коричневый цвет крыльев, окраска же самцов может быть синей, коричневой, зеленоватой, серебристой и даже оранжевой. Самки узнают «своих» самцов по цвету крыльев.
В начале исследования была проведена филогенетическая реконструкция рода бабочек голубянок по фрагментам митохондриальных генов. Было изучено 89 видов, соединенных 88 узлами ветвления: 66 из них представляют сестринские аллопатрические пары, а 28 – симпатрические. Принципиальные изменения окраски крыльев отмечены 19 раз: 15 раз у симпатрических видов, и только 4 случая у видов с разграниченным местообитанием. Из этого следует, что чаще цвет крыльев различается у видов, проживающих на одной территории.
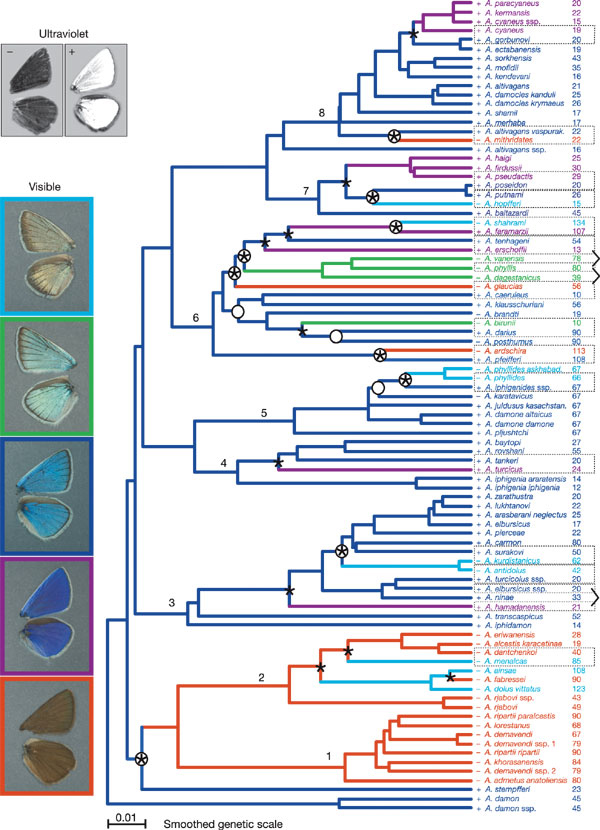
Далее было высказано предположение, что филогенетически старые таксоны, длительно пребывавшие в изоляции, накопили за это время такое количество расхождений по разным признакам, которого уже достаточно для создания презиготического барьера; этим таксонам нет нужды резко менять цвет при встрече, дабы не перепутаться. Конечно, и у них периодически появляются различия по окраске, но случайно и сравнительно медленно – не быстрее, чем при раздельном проживании. Молодым сселившимся таксонам, напротив, необходимо в ускоренном режиме приобретать презиготическую изоляцию, чтобы избежать репродуктивных потерь при невыгодном межпопуляционном скрещивании. Оказалось, что доля симпатрических пар с контрастной окраской действительно максимальна среди молодых видов и уменьшается с возрастом. Итак, наблюдения за динамикой накопления различий в окраске крыльев между видами во времени подтверждают преобладание видов, развивающихся по сценарию Добржанского.
Анализ экониш голубянок выявил отсутствие корреляции между пигментами кормовых растений гусениц и последующим цветом крыльев выросших бабочек. Окраска крыльев также не была следствием температурного режима: крылья не осветлялись при высоких температурах и не темнели в холодных биотопах. Следовательно, расцветка бабочек не связана с экологическими условиями, а служит исключительно для маркировки видов, для различения своих и чужих.
Дополнительное исследование показало видоспецифичность кариотипов у Agrodiaetus: количество пар хромосом варьировало у разных видов от 20 до 268. Хромосомный анализ представителей симпатрических таксонов не выявил ни одного межвидового гибрида.
Таким образом, можно видеть, что предложенная Добржанским схема видообразования, которое стартует в аллопатрии, а при вторичном контакте популяций усиливается, доводя репродуктивную изоляцию до 100%, является наиболее правдоподобным объяснением картины эволюционных преобразований, обнаруженных в роде Agrodiaetus.
Последние выпуски
- Том 86, № 4. Июль-август 2025
- Том 86, № 3. Май-июнь 2025
- Том 86, № 2. Май-июнь 2025
- Том 86, № 1. Январь-февраль 2025
Популярные синопсисы




