Новый сценарий происхождения эукариот
Предложен новый сценарий происхождения эукариот путем симбиоза метаногенной археи с древней спорообразующей клостридией. Гипотеза основана на сходстве процесса формирования эндоспор у клостридий с фагоцитозом, а также на том, что у некоторых клостридий перемещение споры внутри материнской клетки осуществляется при помощи белковых структур, похожих на микротрубочки. Возможно, метаногенная архея заместила собой формирующуюся спору и была «проглочена» материнской клеткой спорангия. Архея дала начала ядру, а цитоскелет материнской клетки лег в основу митотического аппарата.
Появление эукариот было важнейшим событием в развитии жизни, которое существенно изменило структуру биосферы и открыло принципиально новые возможности для прогрессивной эволюции (см. обзор «Происхождение эукариот»). Сейчас считается твердо установленным фактом, что митохондрии эукариотической клетки произошли от симбиотических альфапротеобактерий, а пластиды – от цианобактерий. Гораздо более спорным является вопрос о происхождении клеточного ядра – главной отличительной особенности эукариотической клетки, а также цитоскелета.
Цитоплазма и ядро эукариот (вместе их иногда называют нуклеоцитоплазмой) сохранили много специфических черт, характерных для архей. По-видимому, это означает, что археи сыграли важную роль в становлении эукариот. Но многие особенности нуклеоцитоплазмы не встречаются у современных архей (прежде всего, это два вышеупомянутых признака: наличие ядра и цитоскелета, а также способность к фагоцитозу), а многие ключевые гены имеют не архейное, а бактериальное происхождение (см.: А.В.Марков, А.М.Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе). Для объяснения этих фактов предложены десятки различных гипотез. В статье Г.Г.Гончикова из Института общей и экспериментальной биологии СО РАН (г. Улан-Удэ) предложен новый сценарий происхождения ядра и цитоскелета эукариот. По мнению автора, нуклеоцитоплазма эукариот сформировалась в результате эндосимбиоза бактерии, сходной с некоторыми современными клостридиями, и археи, которая была «проглочена» клостридией и дала начала ядру.
Идея основана на следующих фактах и соображениях.
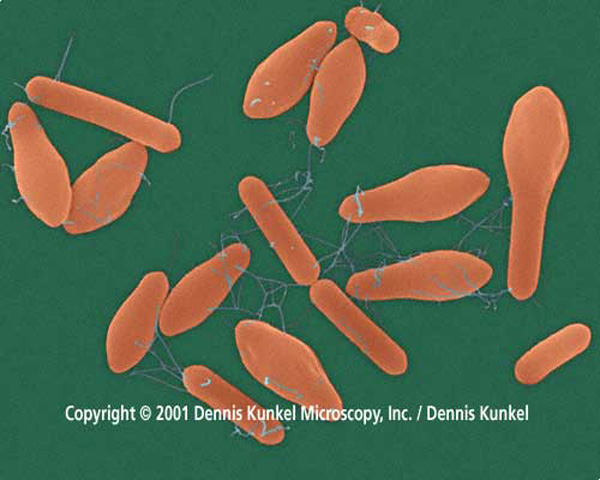
Во-первых, некоторые бактерии, в том числе клостридии, образуют так называемые эндоспоры. Эндоспоры – это покоящиеся бактериальные клетки, которые образуются внутри материнской клетки («спорангия»). В ходе образования эндоспор имеет место так называемый ингалфмент (engulfment) – поглощение, обволакивание материнской клеткой формирующейся споры. Этот процесс сходен с фагоцитозом. При этом возникает уникальная ситуация «клетка в клетке», вообще-то для прокариот не характерная и отдаленно напоминающая то, что мы видим у всех эукариот: снаружи клеточная мембрана, затем цитоплазма, потом еще одна (ядерная) оболочка, окружающая нуклеоплазму с ДНК.
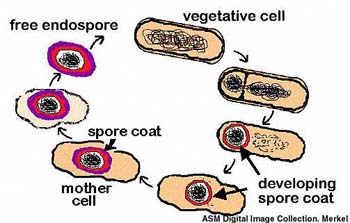
Во-вторых, у некоторых видов клостридий во время формирования эндоспор в спорангии образуются белковые структуры, очень похожие на цитоскелет, в том числе микротрубочки, прикрепляющиеся к эндоспоре снаружи. Когда зрелая спора выходит из спорангия, эти придатки остаются прикрепленными к ней в виде внешних структур. По-видимому, рост придатков проспоры (формирующейся споры) в спорангии происходит «путем агрегации белковых глобул в лентообразно и цилиндрически расположенные тяжи». Рост этих придатков ведет к перемещению (миграции) проспоры в спорангии. В результате проспора прижимается к одному из полюсов материнской клетки. Таким образом, функции «цитоскелета» клостридий – «силовые», двигательные – напоминают таковые эукариотического цитоскелета в процессе клеточного деления (митоза).
Если предположить, что на месте проспоры в материнской клетке каким-то образом оказалась эндосимбиотическая архея, которая была «проглочена» материнской клеткой путем ингалфмента вместо проспоры, и к которой затем пристроились микротрубочки, то получится химерный организм, который мог бы претендовать на роль прообраза эукариотической клетки.
Клостридии относятся к группе фирмикут (Firmicutes), которая, по-видимому, является одной из древнейших групп живых организмов на Земле.
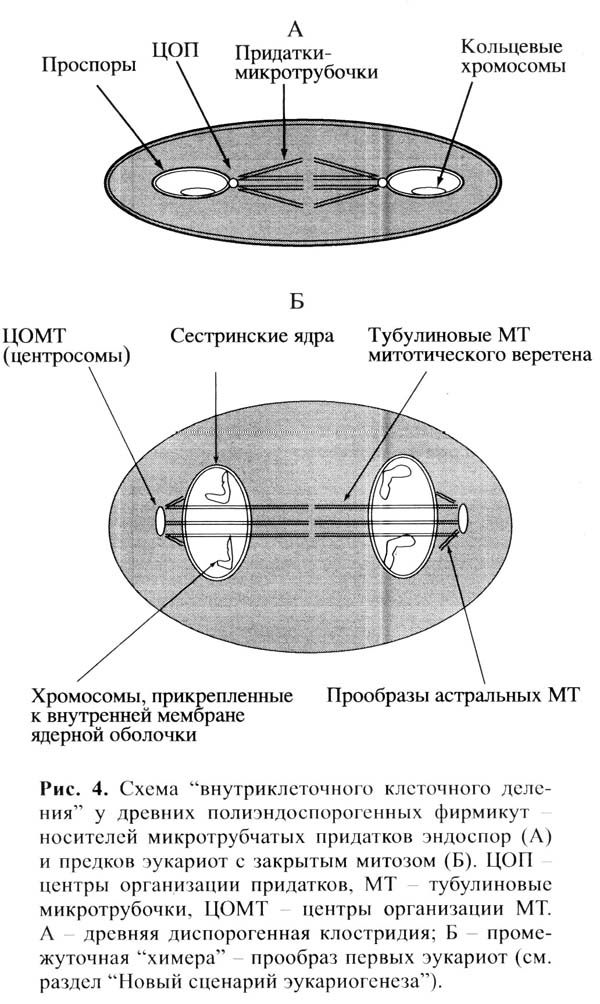
Автор отмечает, что у некоторых фирмикут спорангий может образовывать не одну, а несколько (до 5-7) зрелых эндоспор. Возможно, образование множественных эндоспор – «полиэндоспорогенез» – был предшественником механизма деления ядра в ходе митоза у эукариот. То и другое – своего рода «деление клетки в клетке», причем «сегрегация хромосом, цитодифференциация, процессы внутриклеточного транспорта и их регуляция у спорулирующей бактериальной и митозирующей эукариотической клеток схожие».
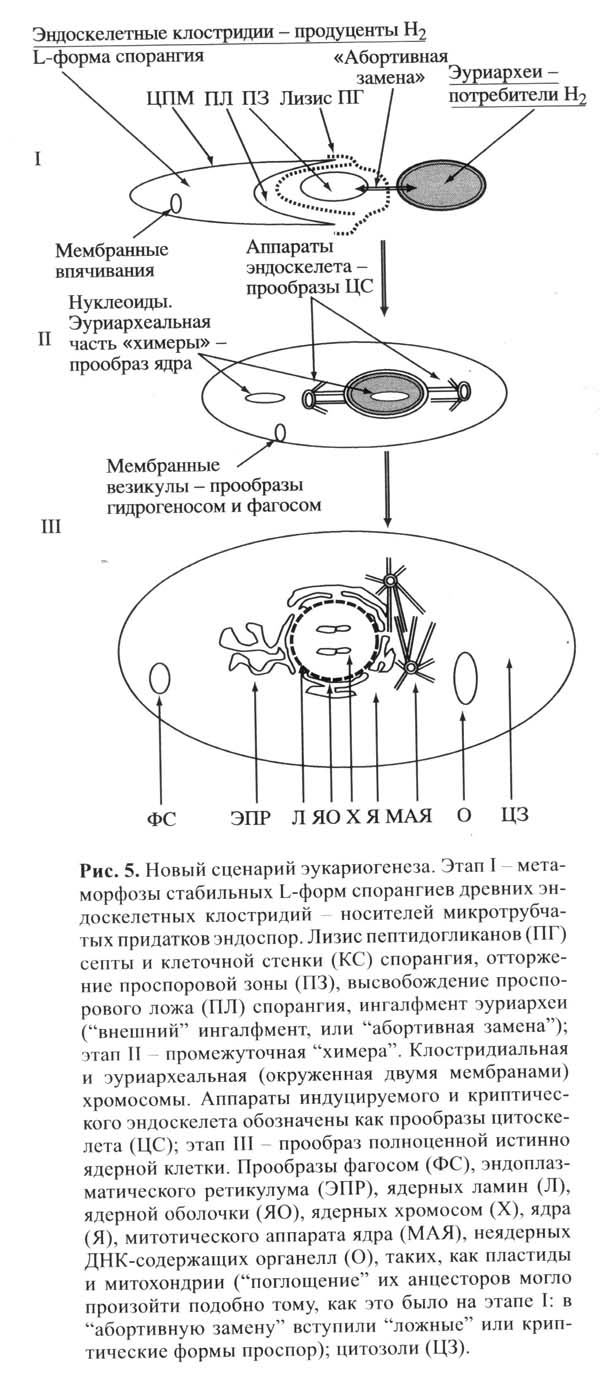
Автор предлагает следующий сценарий происхождения эукариот. Перед этим событием на планете истощились запасы абиогенного водорода, который необходим археям-метаногенам. Тогда они вступили в симбиоз с клостридиями-бродильщиками, которые выделяют водород в качестве отхода жизнедеятельности. Сначала эти микробы просто жили по соседству в одном микробном сообществе (бактериальном мате).
Первым шагом к объединению стало появление стабильных L-форм спорангиев у клостридий (то есть «оголенных» спорангиев, лишенных жесткой клеточной стенки). Такое иногда бывает у прокариот. Затем у какого-то «оголенного» спорангия проспора заместилась симбиотической археей-метаногеном, которая подверглась ингалфменту вместо проспоры и оказалась внутри материнской клетки спорангия.
Получилась химера, содержащая две кольцевые хромосомы в разных компартментах клетки: архейная хромосома внутри будущего ядра (клетки археи), и хромосома клостридии в цитоплазме материнской клетки. В дальнейшем геном клостридии «перебрался» в ядро. Митотический аппарат сформировался на основе микротрубчатого цитоскелета спорангия, причем это могло произойти практически сразу, одномоментно, на основе имевшихся у эндоскелетеных клостридий механизмов деления спор внутри спорангия. Изначально протоядро химеры делилось по прокариотному типу: ядерная оболочка не разрушалась, хромосомы оставались прикрепленными к оболочке (нечто подобное наблюдается у современных жгутиковых водорослей-пирофитов).
Автор подчеркивает, что для проверки предложенной гипотезы важное значение будет иметь анализ полных геномов Clostridium penicillum и других эндоскелетных бактерий – носителей цилиндрических (микротрубчатых) придатков эндоспор.
См. также:
А.В.Марков, А.М.Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе.
Для эукариот более важны гены, полученные от архебактерий. «Элементы», 15.10.10
Последние выпуски
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
- Том 86, № 4. Июль-август 2025
- Том 86, № 3. Май-июнь 2025
Популярные синопсисы