Семейства двудольных растений появлялись строго по графику
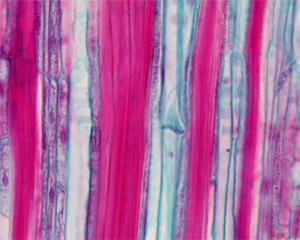
Семейства двудольных растений можно разделить на три группы по строению концевых участков проводящих пучков (мелких жилок в листьях). Сопоставив эту классификацию с данными по времени появления семейств в палеонтологической летописи, Ю. В. Гамалей и его коллеги обнаружили, что все семейства первой группы появились в мелу, второй – в палеогене, третьей – в неогене. Столь строгих, абсолютных закономерностей в ископаемой летописи до сих пор никому не удавалось обнаружить. Строение жилок связано с адаптацией к определенным климатическим условиям, и последовательность появления семейств двудольных хорошо согласуется с данными по глобальным изменениям климата.
1. Три типа маленьких жилок
Свыше 30 лет Юрий Владимирович Гамалей и его коллеги из БИНа занимаются изучением так называемой терминальной флоэмы двудольных растений. Флоэма, или луб – это проводящая ткань растений, которая служит для транспортировки питательных веществ, образующихся в ходе фотосинтеза, из листьев в другие части растения. Терминальная флоэма – это флоэма концевых участков проводящих пучков, самых маленьких жилок в листьях. Именно здесь, в кончиках жилок, происходит «загрузка» флоэмы, то есть транспортировка питательных веществ из фотосинтезирующих тканей листа в ситовидные трубки (так называются проводящие сосуды флоэмы). В начале 1970-х годов ботаники из Королевского Университета в Белфасте и Ю. В. Гамалей с коллегами из БИНа обнаружили, что в терминальной флоэме двудольных встречаются два контрастных типа так называемых клеток-спутников, играющих ключевую роль в загрузке флоэмы. В соответствии с этим было выделено два типа терминальной флоэмы: симпластный (флоэма загружается при помощи системы межклеточных цитоплазматических мостиков – плазмодесм) и апопластный (питательные вещества транспортируются через клеточные мембраны с большими затратами энергии). Симпластный тип характерен в основном для деревьев, апопластный – для трав, но есть и немало исключений. Позже был выделен третий тип – анцестральный, то есть предковый, неспециализированный, самый примитивный. Анцестральный тип может эволюционировать в одном из двух альтернативных направлений – либо в сторону симпластного, либо в сторону апопластного типа.
В течение 30 лет Ю. В. Гамалей с коллегами изучал терминальную флоэму у разных видов двудольных, а результаты заносились в базу данных. К настоящему моменту проанализировано 2300 видов, относящихся к 850 родам, 126 семействам и 8 подклассам класса двудольных – весьма представительная выборка.
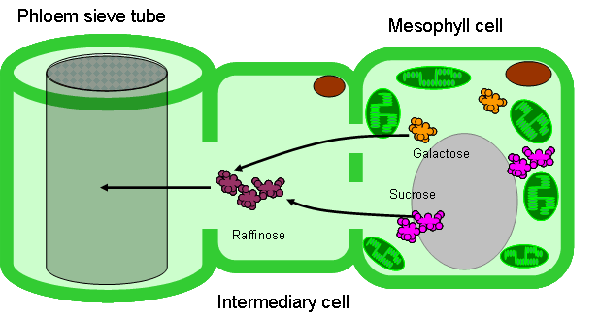
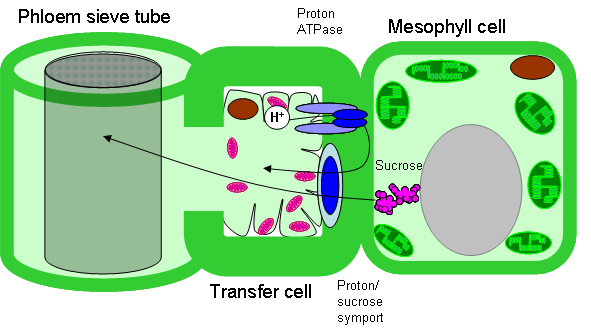
2. Абсолютная закономерность №1: в пределах семейств признак не варьирует
На уровне видов и родов тип терминальной флоэмы не варьирует. Если у одного вида данного рода присутствует симпластный тип, значит, и у всех остальных видов наверняка будет он же. Немного странно, но все-таки в пределах вероятного. А что с семействами? На первых этапах исследований складывалось впечатление, что в большинстве семейств двудольных встречается только какой-нибудь один тип терминальной флоэмы, однако в некоторых семействах присутствует более одного типа. Это было бы вполне естественно, и вот почему. Оба специализированных типа (симпластный и апопластный) встречаются в разных порядках и подклассах двудольных, а это значит, что они возникали много раз совершенно независимо в разных эволюционных линиях (параллелизмы в эволюции случаются часто, и отнюдь не только у растений). Следовательно, вероятность эволюционного перехода от одного типа к другому (по крайней мере от анцестрального типа к любому из двух специализированных) весьма высока. А если так, почему этот признак должен обладать абсолютной устойчивостью в пределах каждого семейства? Семейства у растений бывают весьма разнообразны и по морфологии, и по экологии; признак – экологический значимый и эволюционно пластичный. Он просто обязан варьировать внутри семейств!
Однако, как выяснилось в дальнейшем, он все-таки не варьирует. Те семейства, в которых было более одного типа терминальной флоэмы, были впоследствии подразделены (на основе молекулярно-генетических данных) таким образом, что теперь в каждом семействе остался только один тип. Невероятно, но факт.
3. Абсолютная закономерность №2: семейства появлялись в строгом хронологическом порядке
Авторы сопоставили распределение типов терминальной флоэмы в семействах двудольных со временем первого появления этих семейств в ископаемой летописи. Выявилась удивительно строгая закономерность. Практически все «анцестральные» семейства появились в мелу, начиная со 140 млн лет назад, и полностью прекратили появляться около 60 млн лет назад (начало палеогена). Все «симпластные» семейства появились в промежутке между 68.1 и 19.5 млн лет назад. Ни до, ни после этого интервала ни одно симпластное семейство не появилось. Почти все моменты появления симпластных семейств приходятся на палеоген. Наконец, период появления апопластных семейств начался 31.2 млн лет назад и продолжается по сей день. Подавляющее большинство апопластных семейств появилось в неогене.
Нужно заметить, что соответствующие данные приведены в статье в виде трех таблиц, и между этими таблицами есть кое-какие расхождения. Вышеприведенные цифры взяты из табл. 2. Однако табл. 3 показывает еще более «абсолютизированную» картину: здесь уже мы видим, что все без исключения анцестральные семейства появились в мелу, все до единого симпластные – в палеогене, апопластные же появлялись исключительно в неогене. Расхождения между таблицами состоят в том, что время появления некоторых семейств указано в них по-разному. Например, возраст апопластных семейств Asteraceae, Boraginacea, Solanaceae, согласно табл. 2, составляет 31.2 млн лет (ранний олигоцен), однако в табл. 3 они помещены в графу, соответствующую промежутку времени от 10 до 20 млн лет (миоцен). Возможно, это связано с тем, что авторы использовали палеонтологические датировки из нескольких разных источников; досадно, что в статье не объясняются причины расхождений между таблицами.
Независимо от того, какую «версию» данной закономерности мы выберем в качестве основной – менее абсолютизованный вариант из табл. 2 или более абсолютизированный из табл. 3 – следует признать, что такой строгой закономерности в хронологии параллельного появления семейств с определенными признаками никому ранее не удавалось обнаружить.

4. Все объясняется глобальными изменениями климата?
По мнению авторов, обнаруженная закономерность может быть связана с климатическими изменениями. В палеогене было тепло и влажно – это оптимальные условия для «симпластных» растений. Именно такие растения и появлялись в палеогене. Ну а потом, когда климат стал холоднее и суше, они уже не могли перейти от симпластного типа обратно к анцестральному или, тем более, к апопластному. Но они не вымерли, а продолжают процветать там, где сохранились подходящие для них условия – в первую очередь в дождевых тропических лесах. И их разнообразие осталось очень высоким – около 100 000 видов! Вот только новые семейства симпластных растений уже не появляются. В статье говорится даже, что «в неогене не появилось ни одного нового семейства или хотя бы вида симпластных двудольных». Это, я думаю, опечатка. Чтобы 100 000 видов за 20 млн лет не породили ни одного нового? Да еще при том, что все это происходит в тропических дождевых лесах? Так не бывает. Или же все виды, порожденные ими, имели апопластную или анцестральную терминальную флоэму? Тоже не годится, ведь мы знаем, что в пределах семейств типы флоэмы не варьируют, следовательно, каждый такой новый вид автоматически давал бы начало новому семейству двудольных... К тому же и палеонтологическая летопись не дает оснований для таких утверждений, т.к. на видовом уровне она весьма малопредставительна, и на ее основе нельзя судить о том, появлялись или нет в неогене новые симпластные виды.
В конце палеогена началось оледенение Антарктиды, и климат на всей планете стал холоднее и суше. Пышные леса стали сменяться травяными биомами – степями и саваннами. Это был совершенно новый, доселе невиданный тип растительных сообществ. По мнению многих палеонтологов, в его формировании, помимо климатических факторов, немалую роль сыграли крупные растительноядные млекопитающие, в том числе жвачные. В условиях дефицита тепла или влаги апопластные растения получают преимущество по сравнению с симпластными. Поэтому именно в неогене стали появляться апопластные семейства, которые и по сей день доминируют в травяных биомах.
Что касается анцестральных семейств, то они, с одной стороны, появлялись в благоприятных климатических условиях (в мелу было тепло и влажно). С другой стороны, в наши дни они, согласно приведенному в статье графику, приурочены в основном к экстремальным местообитаниям (тундрам и высокогорьям). Авторы предполагают, что эти древние семейства были оттеснены в неблагоприятные природные зоны более эволюционно продвинутыми молодыми конкурентами – сначала симпластными, а затем и апопластными двудольными.
График распределения трех типов семейств по климатическим зонам, приведенный в статье, выглядит очень впечатляюще, и лишь немного «недотягивает» до того, чтобы можно было его назвать «абсолютной закономерностью №3». К сожалению, некоторые детали этого графика вызывают вопросы, на которые мне не удалось найти ответ в тексте статьи. Например, график показывает, что практически все представители анцестральной группы приурочены к экстремальным местообитаниям: тундрам, высокогорьям, пустынным степям и полярным пустыням. С другой стороны, в списке семейств этой группы (табл. 2) видим довольно много семейств, названия которых для непосвященного читателя звучат совсем не экстремально: магнолиевые, аралиевые, платановые, кленовые, крапивные, молочайные, лавровые, буковые, тутовые, вязовые, кизиловые, миртовые, розовые, чайные, ореховые, липовые. Сплошь обитатели тундр, засушливых степей и высокогорий? Я не ботаник, и я чего-то тут явно не понимаю. Но Журнал Общей Биологии расчитан на широкую аудиторию, поэтому нелишне было бы разъяснить подобные сложные места для не ботаников.
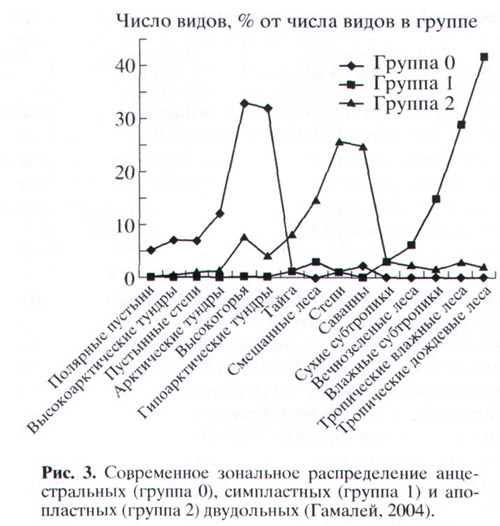
P.S. Должен признаться, что статья Ю. В. Гамалея, М. В. Пахомовой и С. Н. Шереметьева очень меня удивила. Я довольно внимательно слежу за поисками всевозможных закономерностей в палеонтологической летописи, и сам принимаю в этих поисках посильное участие. И если бы меня до прочтения этой статьи спросили «бывает ли такое?», я бы уверенно ответил: «нет!». Мы живем в статистическом мире. Если коэффициент корреляции равен 0.48, если правило соблюдается в 62% случаев, если средние различаются на 8.2%, и вероятность случайности такого различия меньше 0.05 – вот и отлично, на большее никто и не расчитывает. Более того, если вдруг я вижу, что коэффициент корреляции получился равным 1, или правило соблюдается в 100% случаев, то не кричу «эврика», а думаю: «черт, опять я что-то перепутал, придется пересчитывать». Я хотел бы привлечь максимальное внимание к результатам, полученным питерскими ботаниками, потому что они могут иметь важное мировоззренческое значение. Если в эволюции растений могут наблюдаться столь строгие закономерности, то они, вероятно, должны быть и в развитии других групп организмов. Вот только почему никто раньше не находил ничего подобного?
Надеюсь, что авторы не обидятся на меня за доброжелательную критику и естественный в такой ситуации скептицизм. В действительности я хочу лишь обратить внимание авторов и научного сообщества на то обстоятельство, что результаты потрясающе интересны и в значительной мере уникальны, и поэтому их необходимо срочно опубликовать по-английски в международном журнале. Ведь должны же мы, в конце концов, разобраться, что с ними делать: кричать «эврика» (причем если уж кричать, то очень громко), или все-таки что-то пересчитывать?
Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Популярные синопсисы