На пути к разгадке тайны мейоза
Появление мейоза – особого варианта клеточного деления, в результате которого число хромосом сокращается вдвое – было одним из важнейших эволюционных «достижений» первых эукариот. Механизм мейоза сложился из комбинации готовых «блоков»: механизмов митоза, рекомбинации и репарации ДНК. Ключевым событием стало формирование синаптонемного комплекса – особой белковой структуры, обеспечивающей попарное соединение и точное «выравнивание» хромосом.
Появление эукариотической клетки было важнейшим эволюционным преобразованием (ароморфозом) в истории земной жизни (см. обзор «Происхождение эукариот» ). Одним из главных «достижений» древних эукариотических организмов стало возникновение настоящего полового процесса, то есть слияния двух гаплоидных (содержащих одинарный набор хромосом) клеток – гамет в диплоидную (содержащую двойной набор хромосом) клетку – зиготу.
Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.
Общепризнано, что мейоз произошел из митоза – «обычного» клеточного деления эукариот, в результате которого число хромосом остается прежним. Происхождение митоза само по себе было замечательным ароморфозом. Мейоз, по сути дела, является модифицированной версией митоза.
Мейоз представляет собой два последовательных деления исходной диплоидной клетки и отличается от митоза только первым своим делением (см. схему). Отличие состоит в том, что хромосомы вступают в метафазу, соединенные попарно гомолог с гомологом (гомологичными называют хромосомы, содержащие одни и те же гены/локусы и полученные одна от отца, другая от матери). В метафазе I мейоза центромеры каждой из хромосом «униполярны», т.е. соединены белковыми нитями только с одним из двух полюсов веретена деления .
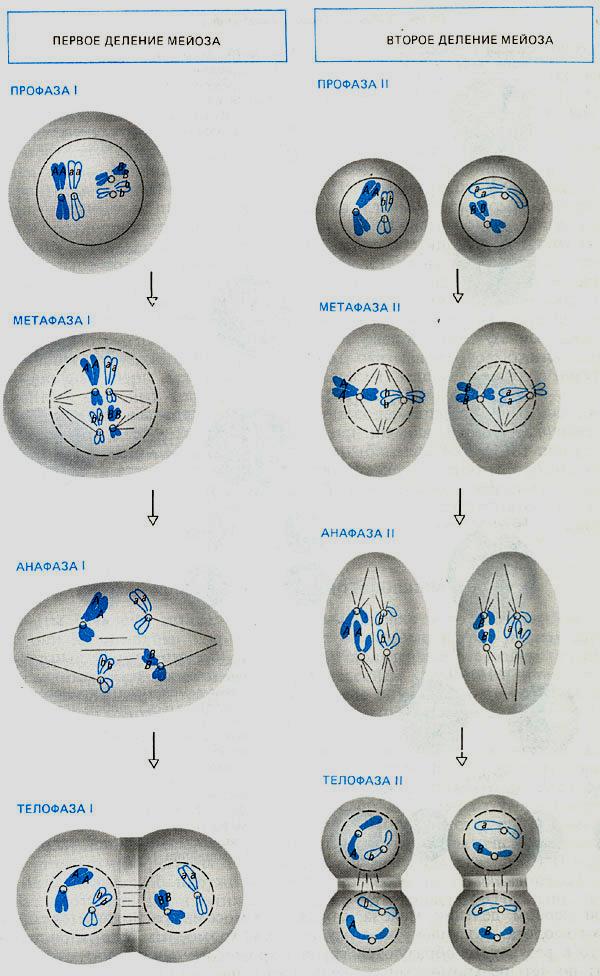
В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Механизм мейоза был «собран» из готовых блоков (как это часто бывает в эволюции) – разумеется, с некоторыми модификациями и новшествами.
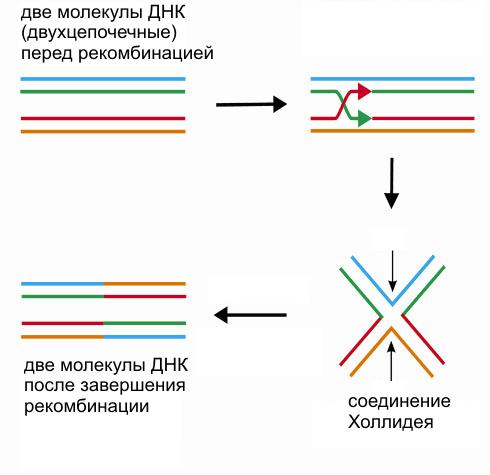
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.: Глазер В.М. Гомологичная генетическая рекомбинация ). Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.
Чтобы «развести» гомологичные хромосомы к разным полюсам клетки, в ходе мейоза используется веретено деления, «унаследованное» от митоза. Но чтобы распределение хромосом по дочерним клеткам прошло без ошибок, гомологичные хромосомы необходимо сначала сгруппировать попарно. Для этого используется механизм гомологичной рекомбинации, точнее, одна из деталей этого механизма – образование так называемого соединения Холлидея.
Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Жизненный цикл древних эукариот (как и многих прокариот), вероятно, состоял из двух фаз и двух соответствующих типов метаболизма: 1) вегетативная фаза (условия благоприятны, клетки размножаются митозом); 2) фаза споруляции (условия стали неблагоприятными, клетки превращаются в споры). У низших эукариот – например, у дрожжей, - перед споруляцией происходит мейоз, и из получившихся гаплоидных клеток образуются споры.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК. Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11. Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Клетка начинает «чинить» разорванные молекулы ДНК при помощи древнего механизма гомологичной рекомбинации. А для этого нужно объединить попарно гомологичные молекулы ДНК (чтобы использовать неповрежденные участки одной молекулы в качестве «матрицы» для исправления повреждений в другой молекуле). На этом и основано попарное объединение хромосом в профазе I мейоза.
Белковые оси хромосом
Эукариотический геном значительно больше прокариотического и содержит больше «избыточной» ДНК (различных некодирующих последовательностей, мобильных генетических элементов и др.) Большие размеры генома стимулировали «распадение» его на отдельные хромосомы (линейные, в отличие от единственной кольцевой хромосомы прокариот). Как следствие, «оказалась полезной новация в структуре хромосом – белковые оси, к которым стали крепиться петли гигантской молекулы ДНК – эукариотической хромосомы».
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Один из когезинов (Rec8) «оказался подходящим субстратом для наслоения на него других мейоз-специфичных белков», из которых в начале мейоза строятся более сложные белковые оси, или «линейные элементы», к которым ДНК крепится петлями, приобретая вид «ершика». Каждая петля содержит несколько тысяч пар нуклеотидов и крепится к линейному элементу только в одном месте, «кнопочно». Такая организация хромосомы позволяет хромосомным локусам легче находить гомологичные им локусы в хромосоме-партнере. Так обстоит дело у некоторых низших эукариот, у которых нет «классического» мейоза, характерного для более высокоорганизованных форм.
Синаптонемные комплексы. Ароморфоз.
У эукариот с классическим мейозом «бывшие линейные элементы хромосом соединяются попарно с помощью белковой «застежки-молнии» в двухосевую структуру, именуемую синаптонемным комплексом (СК)». Это обеспечивает тесное соединение гомологичных хромосом. Ю.Ф.Богданов считает появление СК важным ароморфозом. СК существует ограниченное время в течение профазы I мейоза и затем распадается. СК обнаружен у многих сотен видов – от одноклеточных водорослей, грибов и протистов до многоклеточных. Предполагается несколько возможных функций СК:
1) Организация профазной мейотической хромосомы. Благодаря СК возникает билатеральная организация пары соединившихся гомологичных хромосом. Петли хроматина располагаются по обе стороны от СК. Это позволяет точно сопоставить петли хроматина в трехмерном пространстве клеточного ядра. «По-видимому, это простейший, если не единственный, способ внести порядок и точность в процесс взаимного узнавания локусов гомологичных хромосом и обеспечить достаточную точность (гомологию) рекомбинации в мейозе.»
2) СК не только соединяет гомологичные хромосомы, но и не дает им «склеиться», удерживая их на расстоянии 70-120 нм друг от друга. В конце профазы I (стадия диплотены) СК распадается во всех локусах, кроме локусов хиазм (перекрещивания хроматид). Гомологичные хромосомы, взаимно оттолкнувшиеся во всех локусах, кроме локусов хиазм, выстраиваются на экваторе веретена деления в метафазе I. Они готовы разойтись к полюсам, как только освободятся от хиазм.
3) СК необходим для формирования хиазм. СК также не позволяет хиазмам располагаться слишком близко друг от друга. Иными словами, благодаря СК хроматиды могут «рваться» и обмениваться участками лишь в ограниченном количестве мест. Ю.Ф.Богданов оценивает это как положительное явление, поскольку «чрезмерная частота кроссинговера – не столько «благо» комбинаторики, сколько угроза стабильности адаптационно выгодных фенотипов».
Происхождение белков СК
У разных групп эукариот СК строится по единому «плану», но при этом используются совершенно разные (структурно негомологичные) белки. Это похоже на постройку домов по сходному плану (стены, крыша, окна), но из совершенно разных материалов. Материал не важен – «важно, чтобы СК выравнивал параллельно лежащие гомологичные хромосомы, сохранял между ними пространство, в котором происходит рекомбинация ДНК, и сохранял бы эти условия столь долго, сколько необходимо для завершения рекомбинации и формирования хиазм».

Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Ю.Ф.Богданов приводит убедительные доводы и экспериментальные факты, свидетельствующие о том, что структуры СК формируются путем самосборки из белковых молекул, подобно многим другим сложным молекулярным комплексам. Автор отмечает, что «самоорганизующиеся структуры могут состоять из разных белковых субъединиц и нуклеиновых кислот, как это наблюдается в отношении вирусов и рибосом». Важно, что белки, сходные по размерам и трехмерной организации, могут служить компонентами одинаковых структур у далеких друг от друга организмов даже в тех случаях, когда первичная структура (аминокислотная последовательность) этих белков сильно различается.
Почему в первом делении мейоза сестринские хроматиды не расходятся?
В отличие от митоза и второго деления мейоза, в первом делении мейоза сестринские хроматиды остаются «склееными», потому что ген CDC31, продукт которого нужен для гидролиза когезинов, в мейозе I оказывается временно отключен продуктом другого гена – SGO1 (у дрожжей S. cerevisae). Во время второго деления мейоза ген SGO1 не функционирует, «запрет» на работу CDC31, снимается, и сестринские хроматиды разделяются. У дрозофилы вместо SGO1
Итак, причина нерасхождения хроматид в мейозе I – сохранение соединяющей их когезиновой оси. В результате после первого деления мейоза «программа» клеточного деления остается как бы «не выполненной». Эта программа – в основе которой лежит древняя программа митоза – после завершения мейоза I, не останавливаясь, продолжает работать (хроматиды не разошлись, когезиновые оси не гидролизовались – значит, нужно делиться дальше). И поэтому клетки сразу же, без интерфазы и без синтеза ДНК, вступают в новое деление (мейоз II), идущее уже в точности по схеме классического митоза.
В заключительнй части статьи Ю.Ф.Богданов рассматривает ряд интересных случаев отхода некоторых организмов от схемы «классического» мейоза. Например, у самцов дрозофил (в отличие от самок) СК не образуется, и мейоз идет по примитивной схеме, характерной для некоторых архаичных эукариот (дрожжи S. pombe). Таким образом «генетическая программа» примитивного мейоза у высших организмов не исчезает (вероятно, она просто входит в состав более сложной программы «классического» мейоза) и может вновь оказаться востребованной.
См. также:
Ю. Ф. Богданов. Белковые механизмы мейоза (популярная статья в журнале «Природа»).
С фактами и теориями, касающимися происхождения мейоза, можно ознакомиться в статьях Ю.Ф.Богданова и его коллег:
Yu.F.Bogdanov. 2003. Variation and Evolution of Meiosis .
Ю.Ф.Богданов, С.Я.Дадашев, Т.М.Гришаева. Сравнительная геномика и протеомика дрозофилы, нематоды Бреннера и арабидопсиса. Идентификация функционально сходных генов и белков синапсиса мейотических хромосом .
Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Популярные синопсисы