Древнейшие предки кольчатых червей могли быть похожи на брахиопод

Обзор событий кембрийского взрыва (так обычно называют начавшуюся примерно 540 млн лет назад революцию в животном мире) показывает, что из крупнейших эволюционных ветвей животных последней появляется в палеонтологической летописи группа Spiralia, в которую входят кольчатые черви, моллюски, брахиоподы и еще несколько типов. Выдвинута гипотеза, что уникальной приспособительной стратегией древних спиралий было освоение сидячего образа жизни, сопровождавшееся созданием своеобразной формы тела с изогнутым U-образным кишечником. Эта гипотеза — спорная, но если она окажется верной, то может сильно повлиять на наши представления о ранней эволюции двусторонне-симметричных животных.
От галактики до генома
Какие события можно назвать самыми главными в истории жизни на Земле?
Важнейшим из всех, безусловно, было возникновение жизни как таковой — на нашей планете оно случилось около 4 млрд лет назад. Не менее решающим моментом, судя по всему, оказалось возникновение клеток с ядрами (см., например: М. Никитин, 2014. Выдвинута новая гипотеза происхождения эукариотической клетки). Но уж на третье место большинство биологов наверняка поставит кембрийский взрыв — «почти мгновенное» (по словам некоторых авторов) появление в палеонтологической летописи сразу многих современных типов животных. Событий, настолько сильно поменявших облик Земли, за ее историю было очень мало.
Сухая хронология сообщает нам, что кембрийский взрыв отмечает границу двух периодов — эдиакария (635–541 млн лет назад) и кембрия (541–485 млн лет назад). Все, что было на Земле до кембрийского взрыва, то есть эдиакарий и многочисленные более древние эпохи, традиционно называют докембрием.
Причины кембрийского взрыва не то чтобы не ясны: их слишком много, и ученые не устают спорить о том, какая из них — главная. Больше того, до сих пор спорно, был ли это именно «взрыв», то есть быстрое появление множества новых эволюционных ветвей. Есть гипотеза «долгой скрытой докембрийской эволюции», согласно которой большинство этих ветвей на самом деле возникло гораздо раньше, а в начале кембрия их представители всего лишь обрели скелеты и стали лучше захораниваться (см. Диверсификация животных началась задолго до кембрийского взрыва, «Элементы», 13.12.2011). Гипотезу «взрыва» часто подкрепляют данными палеонтологии и биологии развития, а гипотезу «долгой скрытой докембрийской эволюции» — данными молекулярной систематики. Исследования, проведенные методом молекулярных часов, зачастую «отправляют» предков современных животных даже не в эдиакарий, а в еще более древний период — криогений (720–635 млн лет назад).
Попытки согласовать все эти данные между собой обычно ведут современных авторов к мнению, промежуточному между обеими крайностями: докембрийская эволюция современных типов животных была, но не слишком длинная.
Какими же причинами мог быть вызван кембрийский взрыв (если он реален)? На это есть по меньшей мере три возможных ответа.
Первый ответ дает генетика. Предполагается, что вблизи рубежа эдиакария и кембрия у многоклеточных животных произошла некая генетическая революция, вызвавшая поток «экспериментов» по созданию разных планов строения тела. Однако период этого «экспериментирования» оказался по меркам истории Земли недолгим. Генетическая система, контролирующая развитие, вскоре нарастила дополнительные обратные связи и потеряла гибкость. Сложилось устойчивое консервативное «ядро» (kernel), в чем-то подобное ядру компьютерной операционной системы, которое и сохраняется без особых изменений вот уже 500 млн лет. Именно поэтому новые типы животных с тех пор, насколько мы знаем, больше не возникали. Генетическое объяснение кембрийского взрыва поддерживает, например, крупнейший британский генетик Питер Холланд (см. Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрыва, «Элементы», 13.02.2015).
Второй ответ — экологический. Очевидно, что новые группы организмов массово появляются при наличии вакантных экологических ниш, которые можно занять. Иногда этот процесс называют «заполнением экологической бочки» (filling of the ecological barrel). В начале кембрия он уж точно шел вовсю. С тем дополнением, что эволюционирующие животные не только занимали готовые экологические ниши, но и активно создавали новые, превращая процесс в автокаталитический (см. Цепная реакция видообразования, «Элементы», 11.02.2009). А вот с геномом при этом ничего особенного могло и не происходить, экологических факторов для ускорения эволюции вполне хватило бы. Такой взгляд на вещи много лет развивает американский палеонтолог Дуглас Эрвин, — впрочем, он не склонен к крайностям и возможной роли генетических причин тоже не отрицает (см., например: D. H. Erwin et al., 2011. The Cambrian conundrum: early divergence and later ecological success in the early history of animals).
Наконец, третий возможный ответ — физический. Что, если ускорение эволюции в начале кембрия объясняется какими-то особыми условиями на Земле, наступившими внезапно и сохранявшимися недолго? Гипотезы такого рода всегда имели у серьезных биологов репутацию дилетантских, но это не причина заранее отвергать их. Например, в последние годы кембрийский взрыв всерьез пытаются связать с климатическими изменениями, происходившими из-за взрывов сверхновых звезд (R. Kataoka et al., 2014. The Nebula Winter: the united view of the snowball Earth, mass extinctions, and explosive evolution in the late Neoproterozoic and Cambrian periods).
В номере журнала Gondwana Research, специально посвященном теме кембрийского взрыва, редакционная вступительная статья получила название, напоминающее о хорошей научной фантастике: «По ту сторону кембрийского взрыва: от галактики до генома» (Y. Isozaki et al., 2014. Beyond the Cambrian explosion: from galaxy to genome). Геном, сообщество, планета, Галактика — на любом из этих уровней можно найти факторы, способные повлиять на крупные эволюционные события. Нет сомнений, что в реальности вызванные ими причины и следствия сложнейшим образом переплетались. А вот какой фактор был решающим — «спусковым крючком», так сказать, — это еще только предстоит окончательно выяснить.
О корнях и ветвях
Разобраться в механизме кембрийского взрыва, конечно, будет тем легче, чем лучше мы будем знать сам ход событий (независимо от причинных объяснений). Итак, что же, собственно, в начале кембрия «взорвалось»?
Очередную попытку навести порядок в этом вопросе предпринял Грэм Бадд (Graham Edward Budd) — известнейший специалист по кембрию, британский палеонтолог, занимающий должность профессора в шведском Упсальском университете (Uppsala University). Бадд пользуется для этого терминологией современной филогенетической систематики, или кладистики, понятийный аппарат которой действительно отличается четкостью.
Нужные здесь понятия довольно просты. Все знают, что эволюцию любой группы организмов (во всяком случае, любой группы животных) можно графически изобразить в виде древа. У древа есть корень (общий предок), ствол и отходящие от ствола ветви. Вертикаль, вдоль которой древо тянется — это, разумеется, время. Некоторые ветви древа доходят до современности, а некоторые обрываются раньше, то есть вымирают.
Выделим на древе все ветви, представители которых дожили до современности, и найдем их общего предка. Группа, включающая этого общего предка и всех его потомков — неважно, современных или вымерших, — будет называться кроновой группой (crown group). Остальная часть древа будет называться стволовой группой (stem group). Члены стволовой группы по определению могут быть только вымершими. Если представить эволюционное древо как обычное дерево, то кроновая группа — это крона, а стволовая группа — нижняя часть ствола с прилегающими ветками.
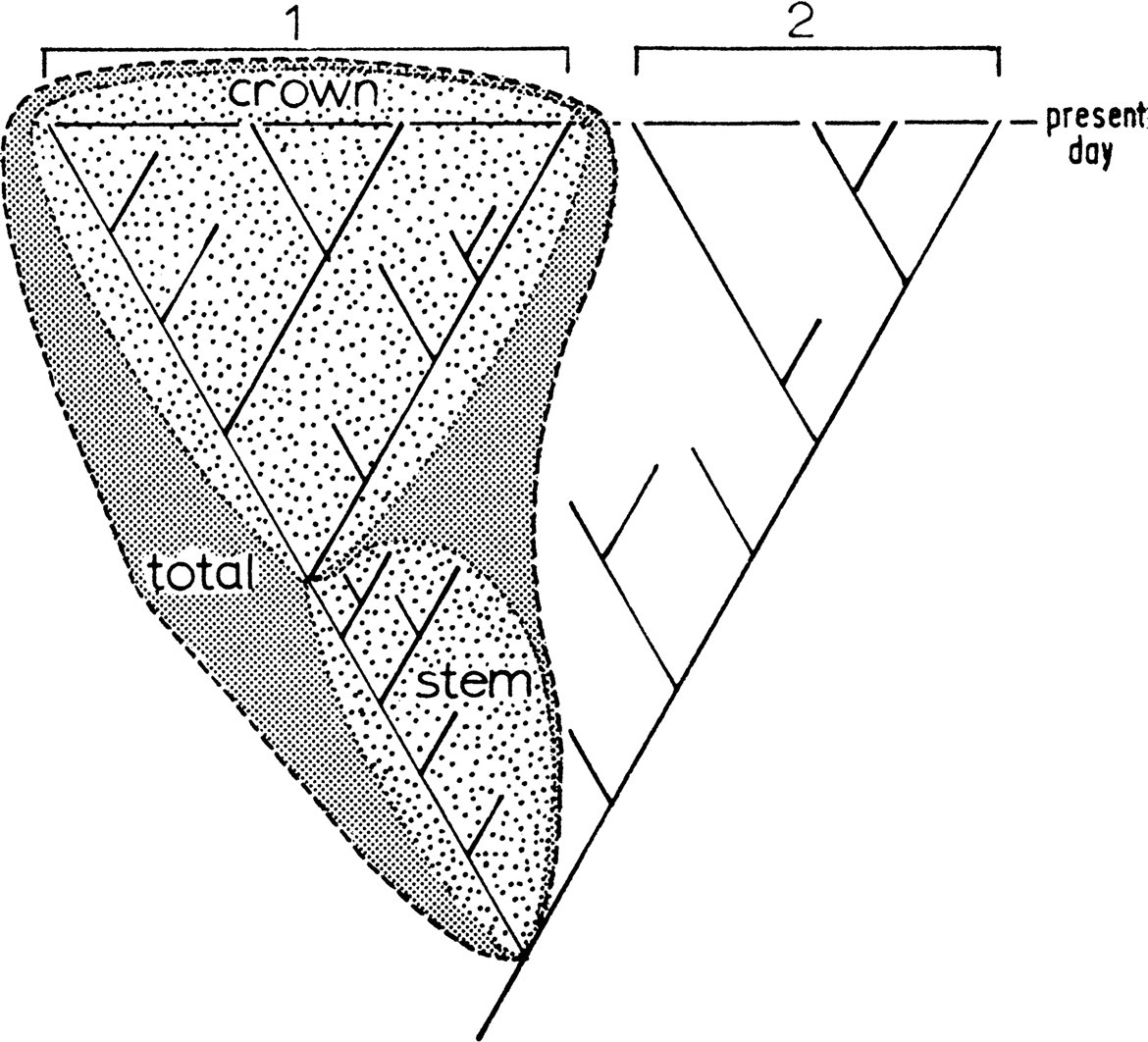
Рис. 2. Кроновая и стволовая группы. Вместе они образуют тотальную группу (total group). Остальные пояснения в тексте. Иллюстрация из статьи: G. E. Budd, 2003. The Cambrian fossil record and the origin of the phyla
Теперь мы можем понять основной тезис Грэма Бадда. Он считает, что кроновые группы всех современных типов животных возникли только после кембрийского взрыва, то есть в кембрийском периоде (а ни в коем случае не в докембрии, как думают сторонники «долгой скрытой докембрийской эволюции»). Более того, для некоторых типов нельзя исключить, что их кроновые группы появились еще позже — в следующем периоде, ордовикском. А вот «корни» современных типов — то есть, говоря по-научному, их стволовые группы — могут уходить и в докембрий. Вряд ли уж очень глубоко, но могут.
Судя по палеонтологической летописи, кроновые группы большинства современных типов животных оформились примерно к середине кембрия (около 515 млн лет назад). Отсюда сразу же вытекают один дополнительный вывод и один вопрос.
Вывод состоит в том, что большинство ископаемых животных из раннего кембрия, даже явно входящих в современные типы, наверняка относится к стволовым, а не кроновым группам этих типов. Это важно, потому что раннекембрийская фауна сейчас изучается довольно активно — понятное дело, она всем интересна. И вот эта фауна — во многих отношениях принципиально иная, чем любые более поздние фауны Земли, от середины кембрия до современности включительно. Например, относить каких-либо раннекембрийских животных к современным классам почти наверняка бессмысленно, этих классов тогда еще просто не было в природе.
Теперь подумаем над возникающим вопросом. Хорошо, пусть кроновые группы современных типов животных появились в кембрии, в крайнем случае в ордовике, но никак не позже. А почему, собственно, не позже? Почему бы кроновой группе того или иного типа не появиться, допустим, в юрском периоде (201–145 млн лет назад, эпоха расцвета динозавров) или в меловом, или в кайнозое? Ведь есть же такие примеры. Скажем, гребневики — очень древний тип, уж его-то стволовая группа точно уходит в докембрий. Но при этом общий предок всех современных гребневиков, по всей видимости, жил геологически недавно — в кайнозое, никак не более 66 млн лет назад (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). В это время и динозавры уже вымерли, и млекопитающие процветали. Однако гребневики — редкое исключение, в подавляющем большинстве типов животных мы такую ситуацию не найдем. Так почему же?
Чтобы ответить на этот вопрос, Грэм Бадд воспользовался помощью своего сведущего в математических моделях коллеги, Ильяма Джексона (Illiam Jackson). Вместе они построили модель растущего эволюционного древа, состоящего из множества ветвей и развилок, с заданными скоростями видообразования (S, от англ. speciation) и вымирания видов (E, от англ. extinction). Получился своего рода компьютерный симулятор эволюции. Одни ветви «вымирали», другие дотягивались до уровня, обозначенного как современность. Использованная Джексоном программа позволяла варьировать значения параметров S и E, выясняя, как меняется вид древа при их разных сочетаниях.
Результаты были следующие. При равных скоростях видообразования и вымирания (S = E) момент появления кроновой группы всегда оказывался примерно в середине суммарного времени роста древа. Если бы реальные типы животных вели себя так, их кроновые группы, как правило, появлялись бы в летописи примерно 270 млн лет назад, то есть были бы гораздо моложе, чем в действительности. При скорости вымирания, превосходящей скорость видообразования (S < E), все ветви компьютерного древа быстро «вымирали» и моделировать дальше было нечего. А вот если скорость видообразования была больше скорости вымирания (S > E), то момент появления кроновой группы смещался к началу древа. И чем сильнее величина S превосходила величину E, тем он сильнее туда смещался. При большой разнице S и E кроновая группа возникала очень рано — интервал между «стартом» и появлением кроновой группы мог составлять, например, всего лишь одну десятую от суммарного времени существования древа. Общий вывод: чем быстрее эволюция, тем короче стволовая группа и длиннее кроновая.
Это идеально соответствует современным данным, согласно которым у предков многочисленных и хорошо изученных типов животных — например, членистоногих — в начале кембрия произошло резкое увеличение скорости эволюции (см. Членистоногие подтверждают реальность кембрийского взрыва, «Элементы», 17.11.2013). Обратим внимание: мы сейчас (вместе с Баддом и Джексоном) не обсуждаем причин этого увеличения, а только анализируем скорости процессов, примерно как в химической кинетике. Кроновые группы современных типов животных возникли рано, потому что их эволюция (по крайней мере в тот период) была быстрой.
С другой стороны, из той же самой модели следует, что «опускающиеся» в докембрий стволовые группы современных типов — иначе говоря, стволовые части их эволюционных деревьев — должны быть короткими. Время их существования — считанные миллионы лет, может быть, первые десятки миллионов, но уж никак не сотни миллионов. В этом плане гипотеза «долгой скрытой докембрийской эволюции» не подтверждается.
Сильно огрубляя, можно сказать, что история наугад взятого современного типа животных, скорее всего, будет делиться на два этапа: (1) эволюция стволовой группы, занимающая самый конец докембрия и начало кембрия, (2) эволюция кроновой группы, длящаяся в среднем примерно с середины кембрия — и до сих пор. Причем по длительности второй этап больше первого примерно на порядок. Так выглядит «кинетическое» описание кембрийского взрыва.
Грэм Бадд считает разумным предположение, что стволовая группа двусторонне-симметричных животных (гигантской ветви, к которой относимся и мы) появилась примерно 560 млн лет назад, а кроновая — через 15–20 млн лет после этого. Вполне может быть.
Древо и взрыв
Ну, а как же в картину кембрийского взрыва вписывается реальное эволюционное древо?
Поясним вопрос. Эволюционное древо животных на уровне типов сейчас, в общем, построено. Спорные моменты в нем есть, но они относительно немногочисленны и никаких научных революций уже не обещают (см., например: Сравнительная геномика вынуждает пересмотреть место гребневиков на эволюционном древе животных, «Элементы», 18.12.2015). Однако это древо строится в основном по молекулярным данным, а значит, оно существует как бы вне времени. Например, мы точно знаем, что общий предок человека и морской звезды жил существенно позже, чем общий предок человека и дождевого червя. Но что в данном случае значит «существенно»? И когда, собственно, жили тот и другой общие предки? Обычное эволюционное древо не дает никаких ответов на эти вопросы. Правда, их можно получить с помощью уже упомянутого метода молекулярных часов, но точность будет довольно низкой.
Надежно привязать узлы древа к историческому времени можно только одним способом: соотнеся их с палеонтологическими данными. Кембрийский период — важнейший в этом отношении, ведь чуть ли не все известные типы животных возникли в кембрии или немного раньше. Это значит, что почти все интересующие нас развилки древа относятся или прямо к моменту кембрийского взрыва, или к его не слишком далекой временной окрестности.

Рис. 3. Эволюционное древо животного мира (до типов). Lophotrochozoa = Spiralia (в том смысле, в каком последнее название употребляется в тексте). Там, где родство не установлено однозначно, альтернативные версии даны пунктиром. Иллюстрация из статьи: M. J. Telford et al., 2015. Phylogenomic insights into animal evolution
Как в целом устроено древо животного мира? От его основания отходят три ветви «низших» животных: гребневики, губки, пластинчатые (последние неизвестны в ископаемом состоянии, поэтому дальше обсуждать мы их тут не будем). На условной вершине древа находится ветвь «высших» животных, которая делится на стрекающих и билатерий, или двусторонне-симметричных. Группа билатерий исключительно велика и разнообразна, поэтому древо билатерий обычно рассматривают отдельно. Оно включает три мощные ветви и одну тонкую. Мощные ветви, в каждую из которых входит несколько типов — вторичноротые, линяющие и спиральнодробящиеся. Тонкая ветвь состоит из одного-единственного типа щетинкочелюстных, или морских стрелок. Линяющих, спиральнодробящихся и морских стрелок вместе называют первичноротыми.
Вот теперь мы можем вместе с Грэмом Баддом взглянуть на палеонтологическую летопись кембрия. В самых низах кембрийской системы можно найти остатки губок, гребневиков («довольно современно выглядящих», по замечанию авторов) и некоторых червей, входящих в большую группу линяющих (см. Китайские палеонтологи нашли древнейшего головохоботного червя, «Элементы», 4.06.2014). Но в целом фауна самого начала кембрия удивительно бедна. Это означает, что «кембрийский взрыв» произошел хоть и быстро, но даже в геологическом масштабе времени не мгновенно.
Следующий важный рубеж проходит около 536 млн лет назад (спустя шесть миллионов лет после границы кембрия). С этого момента скелетные остатки становятся гораздо более разнообразными. В летописи начинают обнаруживаться типичные спиральнодробящиеся, в том числе брахиоподы и ныне вымершие халькиерииды — очень своеобразные существа, близкие одновременно к кольчатым червям и к моллюскам (см. Обнаружено ископаемое животное, близкое к общим предкам моллюсков и кольчатых червей, «Элементы», 6.03.2007). Наконец, примерно 521 млн лет назад появляются «звезды» палеонтологии кембрия — трилобиты, относящиеся, как и все их членистоногие родственники, к линяющим. Они очень быстро достигают расцвета и заполняют все кембрийские моря.
Как обстоит дело с другими группами билатерий — морскими стрелками и вторичноротыми? Предшественники морских стрелок появляются в летописи очень рано — в самом начале кембрия. А вот вторичноротые, наоборот, делаются хорошо заметными довольно поздно. Есть три типа, бесспорно относящиеся к вторичноротым: иглокожие, полухордовые и хордовые. Вероятные следы активности некоторых из них (например, остатки норок, подобных тем, что роют современные полухордовые) встречаются уже в начале кембрия. Но вот самые древние остатки тел вторичноротых — в данном случае иглокожих — обнаруживаются, по современным данным, только 520 млн лет назад. Вторичноротые «собственной персоной» появляются в палеонтологической летописи позже, чем трилобиты. Впрочем, новые открытия вполне могут изменить эту картину.
Загадка спиралий
Разобравшись в первом приближении с родословным древом, можно бросить взгляд на земную фауну. Именно на фауну в целом, как на сообщество, в котором происходит (или не происходит) что-то качественно новое. Можно сказать, что это другой уровень увеличения воображаемого прибора, сквозь который мы смотрим на эволюцию.
Так вот, на уровне земной фауны одним из главных событий первой половины кембрия было появление 536 млн лет назад спиральнодробящихся животных. Две другие крупные группы билатерий — линяющие и вторичноротые — так или иначе мелькают в летописи с самого начала кембрия, с оговоркой, что присутствие вторичноротых там лишь предположительно (по ископаемым следам). Но тут Грэм Бадд замечает, что с точки зрения характеристики фаун ископаемые следы даже надежнее, чем ископаемые тела, потому что сохранение тел гораздо сильнее зависит от анатомических особенностей животных (например, от того, имели они твердый скелет или были мягкотелыми). А никаких следов или остатков, явно принадлежащих спиральнодробящимся (по-латыни они называются Spiralia), в самых низах кембрия нет. Получается, что из крупных групп билатерий спиралии выходят на сцену все-таки последними. Интересно, в чем же был «секрет» спиралий, позволивший им найти в уже начинавшем складываться сложном сообществе свое особое место?
Спиралии уступают линяющим (своей сестринской группе) по числу видов, но, пожалуй, превосходят их по многообразию форм. К спиралиям относятся кольчатые черви, моллюски, брахиоподы, мшанки, форониды, плоские черви, коловратки, немертины и еще несколько типов, мало известных широкой публике, но от того не менее удивительных (например, недавно открытые циклиофоры). Одно это перечисление уже впечатляет. Трудно усмотреть сходство, например, между знакомой каждому садовой улиткой и микроскопической мшанкой, живущей в трубочке в составе прикрепленной подводной колонии. Да и есть ли такое сходство вообще? Ведь единство группы Spiralia в ее нынешнем объеме установлено в основном по молекулярно-биологическим данным.

Рис. 4. Spiralia. Слева вверху — пресноводная улитка Melanoides tuberculata (фото Christoph Kuhne с сайта uni-mainz.de). Справа вверху — брахиопода Lingula (фото Eric Roettinger с сайта discoverwildlife.com). Слева внизу — морской кольчатый червь Nereis (фото Александра Семенова с сайта flickriver.com). Справа внизу — сипункулида Themiste alutacea (фото Alvaro Migotto c сайта cebimar.usp.br)
Конечно, было бы очень интересно придать системе спиралий, как говорится, зоологическую интерпретацию, наглядно объяснив, как был устроен их общий предок, какой он вел образ жизни и как менялись его наследники в разных эволюционных линиях. Но для такой разнообразной группы, родственные связи внутри которой к тому же до сих пор запутаны, сделать это очень непросто. Кроме того, всякая попытка такого рода будет уязвима для критики профессиональных зоологов, которые сразу отыщут контрпримеры к любому предложенному эволюционному сценарию.
Тем не менее Грэм Бадд рискнул выделить один чисто анатомический признак, характерный для многих спиралий и почти исключительно для них. Это изогнутый кишечник, часто связанный с сидячим образом жизни.
Речь идет вот о чем. Типичным билатериям (насколько тут вообще можно говорить о чем-то типичном) свойствен прямой кишечник, соединяющий находящиеся на противоположных концах тела рот и анус. Но у некоторых животных кишечник становится «U-образным», то есть образует петлю, выпирающую в спинную или брюшную сторону. Рот и анус в этом случае могут сильно сближаться. А форма тела у таких животных получается не особо обтекаемой, так что они обычно сидячие или медленно ползающие.
Типичный U-образный кишечник есть у таких спиралий, как брахиоподы. Палеонтологическая летопись брахиопод исключительно подробна — они хорошо сохраняются из-за своих раковин, да и численность их в древних морях была высокой. Ископаемые находки позволяют считать, что U-образный кишечник брахиопод — древний, примитивный признак, имевшийся у этого типа с самого начала, а вовсе не приобретенный недавно.
Сложнее обстоит дело с моллюсками. Самый многочисленный класс в типе моллюсков — брюхоногие, то есть улитки. А в раковине улитки находится огромный мягкий «горб» — внутренностный мешок, заполненный петлеобразно изгибающейся кишкой и прилегающими к ней органами (печенью, например). Чем же это не U-образный кишечник? Правда, у некоторых брюхоногих внутренностный мешок вместе с раковиной исчезает, и кишечник становится прямым: таковы наземные слизни и морские голожаберные моллюски. Но это вторичное состояние, служащее как раз тем исключением, которое подтверждает правило; сравнительная анатомия вполне строго доказывает, что у предков слизняков и голожаберных раковина была.
У крупных и сложно устроенных головоногих моллюсков, которые совсем не похожи на улиток, кишечник тоже изогнут. И у очень древних, чудом доживших до современности глубоководных моноплакофор — тоже. И у лопатоногих моллюсков, малозаметных жителей песчаного морского дна, образующих самостоятельный древний класс, — тоже. Так, может быть, U-образный кишечник — древний общий признак всех моллюсков?
Правда, есть класс моллюсков, которому присущ как раз совершенно прямой кишечник. Это хитоны. Но при таком эволюционном «соседстве» их состояние может быть вторичным — как у слизней, только в масштабе более крупной ветви.
С другой стороны, в типе моллюсков есть вымерший класс Helcionelloida. Это ползающие существа с очень высокой, слегка закрученной раковиной, внешне похожие то ли на улиток, то ли на моноплакофор (когда-то их даже приняли за последних). Внутренняя анатомия гельционеллоид известна плохо, но при такой форме тела U-образный кишечник наверняка был и у них. Проще всего предположить, что большинство моллюсков унаследовало этот признак от единственного общего предка.
А как быть с кольчатыми червями, которые являются довольно близкими родичами моллюсков? У подавляющего большинства из них кишечник прямой — каждый, кто видел дождевого червя или трубочника, согласится, что иначе и быть-то не может. В тонком червеобразном теле U-образной кишке разместиться негде. Но тут Грэм Бадд обращает внимание на сипункулид. Это маленькая (по числу видов), но исключительно интересная группа морских червей, близких к кольчатым. Сипункулиды отличаются от кольчатых червей прежде всего полнейшим отсутствием сегментации. Тело у них относительно компактное, а кишечник петлеобразно закручен, так что анус открывается не слишком далеко от рта. Опять U-образная конструкция! Сипункулиды малоподвижны, переднее положение ануса позволяет им легко выводить продукты обмена наружу, когда большая часть туловища погружена в песок. Но главное — они настолько близки к кольчатым червям, что в последнее время во многих статьях их просто включают в этот тип. Тогда имеет право на существование гипотеза, по которой мешковидное тело с U-образным кишечником есть первичное для кольчатых червей состояние, а классическая червеобразная форма с прямым кишечником — как раз вторичное.
В итоге Бадд предполагает, что приобретение U-образной кишки в сочетании с освоением малоподвижного (сидячего или роющего) образа жизни было уникальным общим признаком (синапоморфией) если не всех спиралий, то, по крайней мере, эволюционной ветви под названием Lophotrochozoa. Брахиоподы, моллюски и кольчатые черви относятся именно к ней.
Надо честно признать: слабых мест у этой гипотезы, что называется, вагон и маленькая тележка. Прежде всего, к ветви Lophotrochozoa относятся еще и немертины — существа, являющиеся прямо-таки воплощением червеобразности. Их прямой сквозной кишечник, по Бадду, тоже должен быть эволюционно вторичным состоянием. Но независимых подтверждений этому маловато.

Рис. 5. Вероятное происхождение плана строения брахиопод (слева) и форонид (справа). Пояснения в тексте. Иллюстрация из статьи: В. В. Малахов, 2009. Революция в зоологии: новая система билатерий
Еще серьезнее то, что Бадд не обсуждает в контексте своей гипотезы такую группу, как форониды. Как и брахиоподы, форониды — донные обитатели, живущие фильтрацией воды; но анатомически они отличаются от брахиопод достаточно сильно. По словам известного зоолога Владимира Васильевича Малахова, «форониды — это антибрахиоподы». У них тоже U-образный кишечник, но если у брахиопод петля направлена на спинную сторону, то у форонид — на брюшную. Взрослая брахиопода как бы сложена пополам брюхом внутрь, а у взрослой форониды кишечник втянут в длинный — длиннее первоначального туловища! — мускулистый брюшной вырост, которым она зарывается в грунт (В. В. Малахов, 2009. Революция в зоологии: новая система билатерий). Это совершенно разные планы строения. Как форониды и брахиоподы могли произойти от общего предка с U-образной кишкой — непонятно.
Всё это никоим образом не значит, что гипотезу Бадда надо отбросить. Ее надо просто проверять. Более того, само ее появление настраивает скорее на оптимизм. Это прекрасный пример взаимодействия новейшей молекулярной систематики со сравнительной анатомией — заслуженной наукой, которая сейчас, по справедливому выражению того же Малахова, вступила в свой новый золотой век.
Источник: Graham E. Budd, Illiam S. C. Jackson. Ecological innovations in the Cambrian and the origins of the crown group phyla // Philosophical Transactions of the Royal Society B. 2016. V. 371. № 1685. P. 20150287.
Сергей Ястребов
-
Разрешите поинтересоваться, кто таков Малахов, о котором восхищенно вспоминают здесь в каждой заметке по биологии?
-
Заведующий кафедрой зоологии беспозвоночных биофака МГУ. Легко гуглится. Там и ссылка есть на Википедию, если я не ошибаюсь.
-
-
да, гуглится, когда знаешь что он зав.кафедрой зоологии. А без этого всё замусорено безумными целителями и ведущими..
Может вы и не с кафедры МГУ, но в вашей и всех других статьях здесь ссылка на Малахова как на критерий истины присутствует всегда. А что там думает Малахов по этому поводу? А не высказался ли еще Малахов по данному вопросу аж в 18-м году? Надо посмотреть у Малахова. И Малахов так считает. А вот Малахов думает иначе. Спрошу у Малахова при встрече... и проч. и проч. Зачем?-
-
Дело не в том. Либо он специалист по всем без исключения вопросам в биологии (что наверняка не так), либо статьи на Элементах исключительно односторонние и на одну и ту же тему (что не очень хорошо для ресурса), либо авторам особо нечего сказать, как в очередной раз зачем-то упомянуть ув.Малахова. Эти отсылки к Малахову по делу и без дела уже режут глаз и воспринимаются как плохой анекдот. Откройте любую статью здесь - найдете Малахова либо в тексте, либо в комментариях.
-
Просто у него несколько довольно интересных работ именно по той тематике, которая сейчас особенно актуальна (или популярна). Причём, что важно, написаны эти работы были существенно раньше, чем тематика стала актуальна.
Ну а если Вам кажется, что ресурс однобок - всё в Ваших руках. Садитесь и пишите. Или наоборот - "не любо, не слушай".
-
-
-
-
Насчет "критерия истины" - это ваша произвольная интерполяция. Я далеко не во всех вопросах согласен с Малаховым, но это на данный момент, пожалуй, крупнейший наш зоолог, с огромным кругозором и с большим количеством удобных для чтения популярных работ, так что его мнение служит удобным ориентиром. И да, он в той или иной мере специалист почти по всем вопросам в _зоологии беспозвоночных_. Естественно, что он упоминается в статьях, посвященных этой теме.
-
Пользуйтесь поиском по научным сайтам на «Элементах»: Владимир Васильевич Малахов
-


-
"Правильно я понимаю, что англичане в моделировании исходили из допущения, что S и E - константы на весь период, охватываемый моделью?"
Конечно, нет! Известно же, что скорости эволюции меняются, и здесь параметры были переменны, модель это позволяла. Просто в популярное изложение не влезли все выкладки.-
Ффух. А то я уж засомневался в природе вещей - не могут же серьёзную работу делать без учёта такой базовой вещи!?
Но тогда тем более непонятно. По-моему, начать стоило, как с моделированием эволюции Солнечной системы, - погонять модель в разных сочетаниях [динамики] параметров, дабы попробовать найти варианты, приводящие к наблюдаемой картине.
Ну да ладно. За такими подробностями, действительно, надо ковырять оригинал, а сил нет.
В любом случае - спасибо, Серёж, за хорошие рассказы об интересных работах. Мониторить саму литературу получается последнее время плохо :(.
-
Ну так, моя идея.
Последние новости











Рис. 1. Некоторые кембрийские ископаемые. a–c — остатки животных, живших ранее, чем 536 млн лет назад: a — протоконодонт Prothertzina unguliformis (элемент ротового аппарата), b — Anabarites tristichus (трубочка), c — Anabarites hexasulcatus (трубочка). Протоконодонты — древние родственники морских стрелок. Anabarites — мелкие раннекембрийские сидячие животные, систематическая принадлежность которых совершенно неизвестна.
d–g — раковины представителей Lophotrochozoa, живших после отметки 536 млн лет назад: d — Barskovia hemisimmetrica (моллюск), e — Anabarella plana (моллюск), f — Purella antiqua (моллюск), g — Tumuldaria incomperta (брахиопода). Иллюстрация из обсуждаемой статьи в Philosophical Transactions of the Royal Society B