Новые данные позволили уточнить родословную животного царства

Новое уточненное эволюционное древо животного царства, основанное на анализе рекордного числа генов и типов животных, позволило разрешить многие спорные вопросы эволюции и систематики. Подтвердилась теория, согласно которой разделение на первичноротых и вторичноротых произошло еще до того, как у животных сформировался целом (вторичная полость тела). Первичноротые подразделяются на две четкие эволюционные линии: Lophotrochozoa (плоские и кольчатые черви, моллюски, брахиоподы, немертины) и Ecdysozoa (круглые и головохоботные черви, членистоногие, онихофоры, тихоходки).
Вплоть до последней четверти XX века биологи реконструировали эволюционную историю животных преимущественно на основе данных сравнительной анатомии, эмбриологии и палеонтологии. Затем к этому списку добавились молекулярные данные, самыми важными из которых являются нуклеотидные последовательности ДНК. Эволюционные реконструкции («деревья»), основанные на молекулярных данных, не всегда совпадали со старыми «классическими» деревьями. Это приводило к бурным спорам среди зоологов.
Поначалу многие придерживались мнения, что старые проверенные методы надежнее новомодных молекулярных. Но постепенно чаша весов склонилась в другую сторону, и сегодня большинство специалистов считает, что молекулярные данные в принципе позволяют гораздо более точно реконструировать пути эволюции животных, чем морфологические и эмбриологические признаки. В России, правда, до сих пор многие не согласны с этим, но на Западе таких «ретроградов», не доверяющих молекулярным реконструкциям, осталось очень мало.
Молекулярные «признаки» (последовательности нуклеотидов) имеют два важных преимущества по сравнению с морфологическими. Во-первых, их просто гораздо больше. Фактически каждый нуклеотид в хромосоме можно рассматривать как отдельный признак — и таким образом получать деревья, основанные на многих сотнях и тысячах признаков, тогда как число морфологических признаков, пригодных для филогенетического (эволюционного) анализа, обычно ограничивается несколькими десятками. Во-вторых, большинство морфологических признаков непосредственно влияют на жизнеспособность организма, тогда как замены многих нуклеотидов являются нейтральными (безразличными). Морфологическое сходство не обязательно свидетельствует о родстве — оно может развиться и у неродственных организмов под воздействием естественного отбора в сходных условиях обитания (это явление называют конвергенцией). Конвергентное возникновение сходных нуклеотидных последовательностей гораздо менее вероятно.
Проблема, однако, состоит в том, что достоверность любых эволюционных реконструкций, в том числе молекулярных, очень сильно зависит от объема и полноты исходных данных.
Главным критерием достоверности молекулярных деревьев считается их устойчивость или повторяемость. Существует несколько разных алгоритмов построения дерева на основе одного и того же массива исходных данных (например, нуклеотидных последовательностей какого-нибудь гена у нескольких разных организмов). Если применение разных алгоритмов дает одинаковый результат, это свидетельствует о его надежности. Разработаны также специальные процедуры для оценки достоверности «узлов» (точек ветвления) получающихся деревьев (см.: bootstrapping).
Первые молекулярные деревья животного царства, основанные на единичных генах и очень небольшом количестве видов, отличались низкой устойчивостью, и потому вызывали мало доверия. Очень скоро стало ясно, что чем больше генов и групп животных вовлечено в анализ, тем устойчивее и надежнее становятся результаты. Ученые, разумеется, стали изо всех сил наращивать объем используемых данных. Постепенно стала вырисовываться картина, довольно сильно отличающаяся от «классической», основанной на морфологии и эмбриологии.

Важнейшее из выявленных отличий касалось родственных отношений между основными типами двусторонне-симметричных животных (билатерий). Согласно классическим представлениям, все билатерии, имеющие целом (вторичную полость тела), происходят от общего предка и противопоставляются «доцеломическим» билатериям, таким как плоские и круглые черви. Целоматы подразделяются на первичноротых (кольчатые черви, моллюски, членистоногие и др.) и вторичноротых (хордовые, полухордовые, иглокожие). Кольчатые черви считались предками членистоногих.
Молекулярные данные, напротив, показывали, что разделение на две линии, соответствующие первично- и вторичноротым, произошло раньше, еще до того, как у билатерий появился целом. Из этого следовало, что целом, который специалисты по сравнительной анатомии считали надежнейшим таксономическим признаком (основой для естественной классификации), в действительности развился независимо у первичноротых и вторичноротых. Не имеющие целома круглые черви, согласно молекулярным данным, оказались близкими родственниками членистоногих (их объединили в группу «линяющих» — Ecdysozoa), а плоские черви — родней моллюсков, а также кольчатых червей, родство которых с членистоногими молекулярные данные никак не хотели подтверждать. Плоских червей (не имеющих целома), а также имеющих целом моллюсков, кольчатых червей и ряд других типов объединили в группу Lophotrochozoa.
Все эти выводы, однако, до самого последнего времени нельзя было признать окончательными. Молекулярные деревья оставались неустойчивыми. Некоторые из них как будто подтверждали «старую» версию эволюции животных, в которой целом появился лишь однажды (эта версия получила название «целоматной теории»). Чтобы разрешить это противоречие, исследователи постепенно наращивали объем и представительность используемых молекулярных данных в надежде, что получаемые деревья в конце концов обретут устойчивость.
Группа ученых из США, Дании, Германии и Великобритании опубликовала в последнем номере журнала Nature новейший вариант молекулярного эволюционного древа животных, основанный на рекордном количестве генов (150) и групп животных. В анализе использовано 77 видов, относящихся к 21 типу животных, причем для 11 из этих типов геномные данные до сих пор отсутствовали. Многие части (узлы) получившегося дерева действительно оказались значительно более устойчивыми, чем в более ранних исследованиях такого рода.
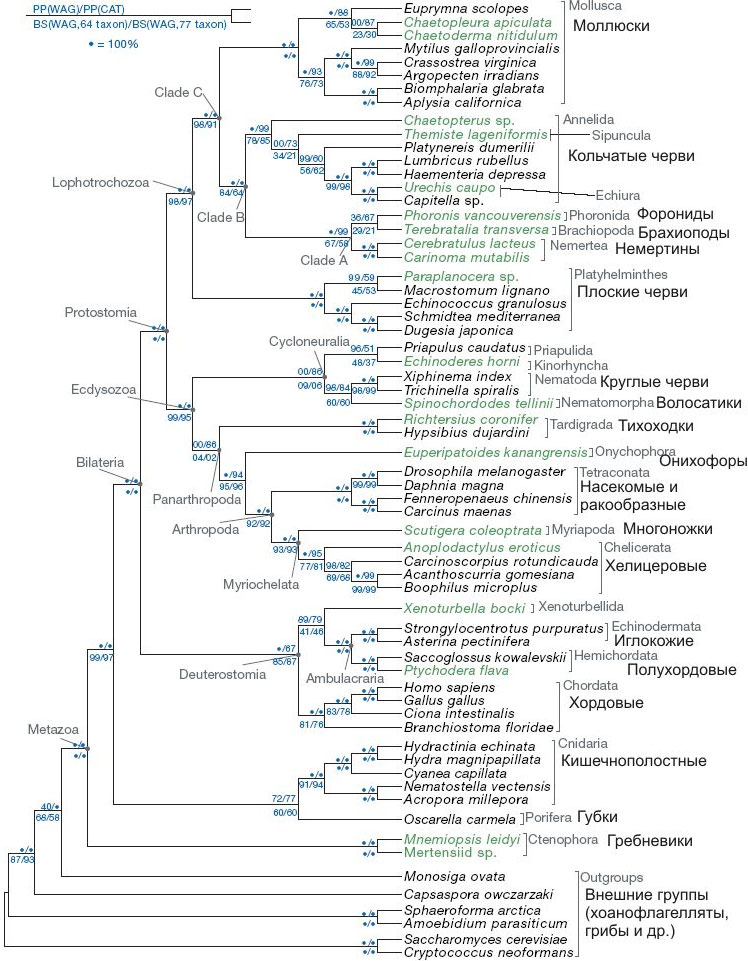
Полученные результаты убедительно свидетельствуют против классической «целоматной теории». Самыми «примитивными» из включенных в анализ групп оказались гребневики. Билатерии сначала подразделяются на линии первично- и вторичноротых, и только потом в каждой из этих линий независимо формируется целом. Первичноротые подразделяются на Lophotrochozoa и Ecdysozoa. Ближайшими родственниками членистоногих оказались онихофоры и тихоходки (что соответствует классическим представлениям), а также круглые черви (что совершенно не соответствует оным). Ближайшими родственниками кольчатых червей оказались не членистоногие, как считалось ранее, а брахиоподы и немертины.
Многое прояснилось, однако родственные связи некоторых групп так и остались неопределенными (их положение на новом древе оказалось неустойчивым). Эти группы не показаны на приведенном рисунке (за исключением губок). Причины неустойчивости авторы видят в том, что для одних групп они не сумели собрать достаточное количество молекулярных данных (губки, бескишечные турбеллярии, мизостомиды), а другие были представлены недостаточным количеством видов (мшанки, коловратки). Кроме того, авторы не смогли включить в анализ трихоплакса, который, судя по результатам анализа митохондриальной ДНК, является самым примитивным из современных животных.
Источник: Casey W. Dunn et al. Broad phylogenomic sampling improves resolution of the animal tree of life // Nature. 2008. V. 452. P. 745–750.
См. также:
1) Расшифрован геном хоанофлагеллят — ближайших одноклеточных родичей всех многоклеточных животных, «Элементы», 18.02.2008.
2) Cамым примитивным животным на земле оказался трихоплакс, «Элементы», 01.06.2006.
3) Геном актинии оказался почти таким же сложным, как у человека, «Элементы», 11.07.2007.
4) Обнаружено ископаемое животное, близкое к общим предкам моллюсков и кольчатых червей, «Элементы», 06.03.2007.
-
Классификация организмов возникла до эволюционной тeории. Цель тогда была одна - ввести некий порядок при описании существующих видов и предусмотреть место для новонайденных, так чтобы легко найти нужную информацию в огромной базе данных. Делалось это на уровне функциональных и морфологических признаков. Возникновение эволюционной тeории вдруг привнесло огромную идеологическую нагрузку на эту совершенно рутинную процедуру - теперь стали строить 'дерево эволюции'. Инерция такого типа мышления продолжается даже после того, как узнали о горизонтальном переносе ген и полной неадекватности идеи о 'дереве эволюции'. Вопрос 'кто от кого произошел' совершенно лишен смысла, так как ныне живущие организмы могут нести в себе часть или даже полный геном совершенно чужих организмов - вирусов или даже бактерий. Таким образом, наличие 'молекулярных признаков' ровным счетом ничего не доказывает.
-
Открытие горизонтального переноса не может отменить факт происхождения одних организмов от других. Эволюционное древо - это не просто красивая метафора, но и реальность, в которой никто из серьезных исследователей не сомневается. Рекомендую пройти курс сравнительной анатомии позвоночных и тогда многое станет Вам ясно... Что касается обсуждаемой статьи, то она подводит некоторый промежуточный итог того, что специалистам уже было известно. Надо было и раньше больше верить данным эмбриологии, больше внимания уделять личинкам. Тогда бы не сближали так членистоногих и кольчатых червей, не уповали бы на то, что трохофора просто куда-то потерялась за ненадобностью. Ну и тип погонофор надо прикрыть. Увы!
-
Почему погонофор прикрывают? На западе их относят к кольчатым, это подтвердилось по ДНК? Интересно мнение Малахова...
Что касаемо статьи:
-самые интересные группы - Placozoa, Turbellaria, те же Pogonorhora - отсутствуют;
-губки могут пониматься как альтернативная собственно многоклеточности форма существования, которая кстати достаточно успешна - совершенные фильтраторы. Соответственно, логично было противопоставлять их, как на картинке из Догеля;
-Не вчера было высказано мнение, что деление на первично- и вторичноротых следует заменить на преимущественный тип дробления яйца - спиральное и радиальное, которое даёт несколько иное "дерево".
-
-


Последние новости









