Предсказуемый рост приспособленности достигается непредсказуемыми путями

Американские биологи провели эволюционный эксперимент, в ходе которого 640 линий дрожжей, происходящих от 64 генетически различающихся клеток-основателей, приспосабливались к одним и тем же условиям в течение 500 поколений. Рост приспособленности в подопытных линиях шел тем быстрее, чем ниже была исходная приспособленность основателя. В результате к концу эксперимента различия по приспособленности сгладились. Наборы полезных мутаций, закрепившиеся в разных линиях, не зависели от исходного генотипа и оказались разными на нуклеотидном уровне, хотя многие из них затронули одни и те же гены. Исследование не выявило специфических взаимодействий между мутациями, но показало, что полезность одной и той же мутации убывает с ростом общей приспособленности генотипа.
Один из важнейших вопросов эволюционной биологии — вопрос о соотношении случайности и закономерности в адаптивной эволюции. Мутации по большей части случайны, но то, какие из них закрепятся, а какие отсеются, зависит от их влияния на приспособленность (эффективность размножения). Например, если существует лишь одна-единственная мутация, способная повысить приспособленность данного генотипа к данным условиям среды, то процесс адаптации будет вполне предсказуем, невзирая на случайный характер мутагенеза. Все мутации будут отсеиваться до тех пор, пока не произойдет та самая, единственная, которую отбор поддержит.
Если же потенциально полезных мутаций много, то на первый план выходит вопрос об эпистазе (см. Epistasis), то есть о том, как эти мутации друг с другом взаимодействуют. Ход адаптивной эволюции будет определяться тем, как влияют закрепившиеся ранее мутации на полезность тех, что еще не появились.
Если эпистаз отсутствует (то есть эффекты мутаций не зависят друг от друга), то полезные мутации будут закрепляться в произвольном порядке, и ход адаптации будет в значительной мере случаен. Сильный эпистаз ограничивает число разрешенных эволюционных траекторий и делает адаптивную эволюцию более предсказуемой и более зависящей от начальных условий, то есть от того, какие мутации возникнут первыми (подробнее см. в новостях Пути эволюции предопределены на молекулярном уровне, «Элементы», 12.04.2006 и Расширение белковой вселенной продолжается, «Элементы», 24.05.2010).
Элементы уже рассказывали об эволюционных экспериментах, показавших важную роль эпистаза в адаптивной эволюции бактерий (Параллельная эволюция изучена в эксперименте на бактериях, «Элементы», 01.02.2012; В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011). Американские биологи, опубликовавшие результаты своих исследований в свежем выпуске журнала Science, провели аналогичный эксперимент на дрожжах и получили результаты, во многом отличающиеся от тех, что были получены ранее на бактериях.
В эксперименте использовались однополые, гаплоидные дрожжи, не способные к половому процессу (см. Mating of yeast). Правда, некоторые подопытные линии ухитрялись обойти поставленные перед ними препятствия, меняли пол, начинали спариваться и переходили в диплоидное состояние (см. ниже). Приспособленность дрожжей измеряли в конкурентных тестах, заставляя их размножаться на перегонки в смешанной культуре с меченым контрольным штаммом.
Эксперимент состоял из двух этапов (рис. 1). На первом этапе («диверсификация») 432 линии, полученные из единственной предковой гаплоидной клетки, жили в стандартной богатой среде либо при высокой, либо при низкой численности популяции в течение 240 поколений (в сутки сменялось по 10 поколений). Авторы хотели получить набор линий, сильно различающихся по приспособленности. Они исходили из предположения, что в маленьких популяциях из-за генетического дрейфа будут чаще фиксироваться слабовредные мутации, а в больших популяциях, где дрейф слабее, а отбор эффективнее, вредных мутаций накопится меньше, а полезных — больше. Их ожидания в целом подтвердились, и к концу фазы диверсификации они получили набор линий, сильно различающихся по приспособленности. Из этих линий они выбрали 64 клетки-«основателя», причем выборка создавалась таким образом, чтобы покрыть весь спектр вариабельности по приспособленности. Геномы основателей отличались от генома предковой клетки в среднем 4,2 мутациями, которые закрепились за время диверсификации. Наборы мутаций у всех основателей были разные. Это позволило авторам оценить влияние «генетического контекста» на последующий ход эволюции.
Второй этап эксперимента («адаптация») состоял в том, что от каждого основателя произвели по 10 линий, после чего все 640 получившихся линий независимо друг от друга продолжали приспосабливаться к тем же самым условиям в течении 500 поколений. Численность теперь у всех популяций была одинаковая (высокая). Периодически часть каждой популяции замораживали при –80°C, создавая таким образом «живую ископаемую летопись» эксперимента.
Всё это было придумано для того, чтобы оценить, с одной стороны, стохастичность эволюционного процесса, с другой — влияние генетического фона на ход адаптации. Стохастичность оценивали, сравнивая между собой линии, происходящие от одного и того же основателя. Чтобы оценить влияние генетического контекста, сравнивали линии, происходящие от разных основателей.
Приспособленность подопытных дрожжей за 500 поколений выросла в среднем на 6,6%. При этом различия по приспособленности между линиями в значительной мере сгладились (рис. 2).

Рис. 2. Приспособленность подопытных популяций в начале фазы адаптации (синие столбики), в ее середине (зеленые столбики) и в конце (оранжевые столбики). По горизонтальной оси — приспособленность, по вертикальной — число популяций с такой приспособленностью. Видно, что к концу эксперимента распределение стало более узким, то есть уменьшился разброс по приспособленности. Рисунок из обсуждаемой статьи в Science
Сглаживание различий объясняется тем, что линии, произошедшие от основателей с высокой начальной приспособленностью, в ходе эксперимента наращивали свою приспособленность медленнее, чем линии с исходно низкой приспособленностью. Лидеры притормозили, отстающие подтянулись, и в итоге разброс уменьшился. Таким образом, несмотря на неодинаковые стартовые позиции, подопытные популяции за 500 поколений пришли к похожим результатам. Можно сказать, что ход адаптивной эволюции оказался отчасти предсказуемым — если брать в расчет только приспособленность и не вдаваться в ее генетические основы.
Статистический анализ позволил оценить влияние различных факторов на скорость роста приспособленности (рис. 3). Оказалось, что выявленные между линиями различия по темпам роста приспособленности на 29% определяются эволюционной стохастичностью (то есть разнообразием путей, по которым пошла адаптация разных потомков одного и того же основателя), на 21% — ошибками измерений, на 50% — свойствами основателя. Причем из этих свойств самым важным является приспособленность: она определяет 46% из вышеупомянутых 50%, и лишь 4% зависят от конкретных особенностей генотипа основателя.
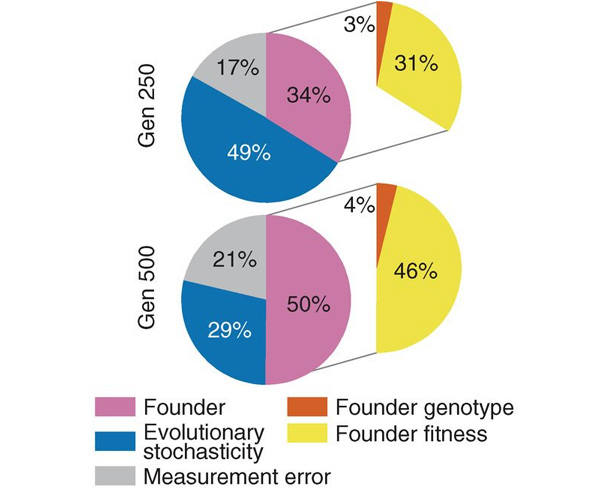
Рис. 3. Вклад различных факторов в различия по величине прироста приспособленности после 250 поколений адаптации (вверху) и после 500 поколений (внизу). Founder — основатель, Evolutionary stochasticity — эволюционная стохастичность, Measurement error — ошибки измерений, Founder genotype — генотип основателя, Founder fitness — приспособленность основателя. Рисунок из обсуждаемой статьи в Science
Иными словами, рост приспособленности в подопытных линиях дрожжей определялся в первую очередь тем, насколько приспособленным был основатель — независимо от того, какой именно набор мутаций обеспечил ему эту приспособленность. Потомки хорошо приспособленных основателей адаптировались медленно, потомки плохо приспособленных — быстро. Конкретный «генетический бэкграунд» оказывал лишь очень небольшое (хотя и статистически достоверное) влияние на скорость адаптации.
Этот результат отчасти согласуется с выводами других эволюционных экспериментов, в которых тоже был показан замедляющийся рост приспособленности. Так, в долгосрочном эволюционном эксперименте Ричарда Ленски было замечено, что каждая закрепившаяся полезная мутация делает последующие генетические усовершенствования в среднем менее полезными. Это явление назвали «эпистазом убывающей доходности» (см. Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает расти, «Элементы», 23.12.2013).
Но придумать название — еще не значит объяснить механизм. Замедление роста приспособленности может быть связано с разными причинами. Например, существует гипотеза «модульного эпистаза». Она предполагает, что многие полезные мутации избыточны в том смысле, что если одна из них уже произошла, то другие ничего не смогут добавить к ее полезному эффекту. Допустим, организму полезно в данных условиях отключить какой-то функциональный модуль. Этого можно добиться, выведя из строя тот или иной ген. Мутаций с таким эффектом может быть очень много. Первая мутация, отключившая ген, будет полезной, и отбор ее поддержит. Но если ген уже отключен, новые мутации, повреждающие его, полезными больше не будут: они станут нейтральными. То же самое будет и в случае, если выгодно не отключить модуль, а усилить его работу. Ведь это тоже можно сделать многими способами (при помощи многих разных мутаций). Замедление роста приспособленности может быть связано с тем, что чем больше модулей уже оптимизировано, тем меньше остается возможностей что-то еще улучшить.
Другая модель («специфический эпистаз») предполагает, что полезность большинства мутаций сильно зависит от генетического контекста, поэтому каждая новая закрепившаяся мутация может резко изменить шансы на фиксацию других мутаций. В этом случае путь к максимальной приспособленности становится похож на лабиринт с множеством развилок и тупиков. География этого лабиринта может быть такова, что чем дальше вы забрались, тем меньше у вас шансов продвинуться вперед еще немного.
Наконец, возможен вариант «глобального эпистаза», когда полезность мутаций мало зависит от конкретного генетического контекста, но сильно зависит от общей приспособленности организма. При этом одна и та же мутация принесет мало пользы хорошо приспособленному организму, а плохо приспособленному даст большой выигрыш.
Если преобладает «модульный» или «специфический» эпистаз, ход эволюции должен сильно зависеть от исходного генотипа. В этом случае подопытные линии дрожжей, произошедшие от одного и того же основателя, будут иметь сходные наборы мутаций. Если же преобладает глобальный эпистаз, наборы мутаций у потомков одного и того же основателя могут различаться так же сильно, как и у потомков разных основателей.
Чтобы проверить эти предположения, авторы отсеквенировали и сравнили полные геномы 104 подопытных клонов, произошедших от 13 основателей. На самом деле было отсеквенировано 150 потомков 15 основателей, но у многих клонов, как выяснилось, произошли «несанкционированные» эволюционные изменения, такие как фиксация аллеля-мутатора (см. Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает расти, «Элементы», 23.12.2013) или восстановление способности к половому процессу и переход в диплоидное состояние. Такие клоны были исключены из рассмотрения.
У остальных 104 клонов обнаружилось в общей сложности 1149 мутаций, закрепившихся на стадии адаптации. У 13 основателей на стадии диверсификации зафиксировалось 55 мутаций. В каждой линии, таким образом, фиксировалось примерно по одной мутации каждые 50 поколений — как на этапе диверсификации, так и на этапе адаптации.
Авторы исключили из рассмотрения синонимичные замены и мутации в межгенных промежутках, потому что они, скорее всего, не имеют функционального значения. Осталось 818 мутаций, предположительно важных для организма. Прежние эксперименты по параллельной эволюции бактерий и вирусов показали, что адаптация к одинаковым условиям часто приводит к независимой фиксации одинаковых мутаций в разных подопытных линиях. Эксперимент на дрожжах дал другие результаты: все выявленные мутации, кроме четырех, оказались уникальными , то есть характерными только для одного клона из 104.
Хотя сами мутации на нуклеотидном уровне были разными, выявилось значительное сходство в наборах генов, в которых закрепились мутации у разных клонов (как и в экспериментах по параллельной эволюции бактерий). Например, обнаружилось 24 гена, мутации в которых независимо закрепились в трех или более клонах. При случайном распределении это число было бы на порядок меньше. Очевидно, мутации в этих 24 генах часто оказывались полезными в условиях эксперимента.
Можно было бы предположить, что у потомков менее приспособленных основателей (чья приспособленность выросла сильнее) в процессе адаптации закрепилось больше функционально важных (и предположительно полезных) мутаций, чем у потомков основателей с высокой приспособленностью. Это, однако, не подтвердилось: у тех и других число закрепившихся предположительно полезных мутаций оказалось в среднем одинаковым. Это согласуется с результатами эксперимента Ленски, где число мутаций росло с постоянной скоростью, хотя приспособленность росла с замедлением (см. Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 02.11.2009).
Самый главный результат авторы получили, сравнив наборы мутировавших генов у клонов, произошедших от одних и тех же и от разных основателей. Оказалось, что происхождение от одного основателя (то есть одинаковый генетический фон) нисколько не повышает вероятность возникновения мутаций в одних и тех же генах. Уровень сходства по набору мутировавших генов оказался одинаковым у клонов с одинаковым и разным генетическим фоном. Этот результат — сильный аргумент в пользу гипотезы «глобального эпистаза» и против моделей «модульного эпистаза» и «специфического эпистаза».
Дополнительные генно-инженерные эксперименты тоже подтвердили глобальный эпистаз. Авторы выбрали три гена, в которых у нескольких клонов независимо закрепились мутации, выводящие эти гены из строя (рис. 4). Это значит, что их отключение, скорее всего, повышает приспособленность в данных условиях. Ученые искусственно отключили эти гены у 13 основателей, у общего предка и у нескольких дополнительных клонов с разной приспособленностью. Оказалось, что эта процедура действительно повышает приспособленность, причем сила положительного эффекта зависит от исходной приспособленности клона. Дрожжи с исходно высокой приспособленностью получают лишь небольшой выигрыш от отключения любого из трех генов, тогда как дрожжам с низкой приспособленностью оно приносит заметно больше пользы.

Рис. 4. Полезный эффект одной и той же мутации убывает с ростом общей приспособленности генотипа, в котором эта мутация произошла. Показано влияние удаления одного из трех генов (gat2, whi2, sfl1) на приспособленность разных клонов дрожжей в зависимости от исходной приспособленности этих клонов. По вертикальной оси — изменение приспособленности (в %) после удаления гена, по горизонтальной оси — исходная приспособленность клона. Удаление гена ho (серые значки), не влияющее на приспособленность, использовалось в качестве контроля. Рисунок из обсуждаемой статьи в Science
Таким образом, исследование показало, что глобальный эпистаз играет в адаптивной эволюции дрожжей важную роль. Чем выше приспособленность, тем меньше пользы приносят новые полезные мутации, причем этот эффект практически не зависит от конкретного генетического контекста. Полезность мутаций зависит только от общей приспособленности, но не от того, какими генетическими особенностями эта приспособленность обусловлена. Причины и механизмы, обеспечивающие глобальный эпистаз у дрожжей, пока неизвестны: для их выявления нужны дополнительные исследования.
Нет пока и ответа на вопрос, почему специфический эпистаз — взаимное влияние мутаций, ярко проявившееся в экспериментах на бактериях, — у дрожжей выявить не удалось. Возможно, это как-то связано с фундаментальными различиями геномной архитектуры прокариот и эукариот (об этих различиях подробно рассказано в книге Евгения Кунина «Логика случая»). Для прокариот характерны огромные популяции, что повышает эффективность очищающего отбора. Возможно, поэтому геномы прокариот оптимизированы и компактны, в них мало «генетического мусора», но мало и степеней свободы для эволюционных преобразований; отдельные функциональные блоки наследственной информации слишком плотно подогнаны друг к другу — в том числе из-за сцепленного наследования, порождаемого отсутствием настоящего полового процесса и относительной редкостью горизонтального обмена генами между организмами. Эукариоты, даже одноклеточные, в среднем гораздо крупнее прокариот, поэтому популяции у них меньше, а очищающий отбор не столь эффективен. Поэтому структура геномов у них более «рыхлая», в них много некодирующих участков и различных эгоистических элементов. К тому же аллели постоянно перетасовываются в ходе полового размножения и потому не так сильно «притерты» друг к другу. Одним из проявлений этих общих свойств эукариотических геномов может быть ослабление зависимости полезности мутаций от конкретного генетического контекста. Впрочем, у нас пока еще слишком мало данных по генетическим основам адаптивной эволюции эукариот, чтобы делать глобальные выводы.
Источник: Sergey Kryazhimskiy, Daniel P. Rice, Elizabeth R. Jerison, Michael M. Desai. Global epistasis makes adaptation predictable despite sequence-level stochasticity // Science. 2014. V. 344. P. 1519–1522.
См. также об «эпистазе убывающей доходности» и замедляющемся росте приспособленности:
1) Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 02.11.2009.
2) Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает расти, «Элементы», 23.12.2013.
О роли «специфического эпистаза» (влияния конкретного геномного контекста на полезность мутаций):
1) Параллельная эволюция изучена в эксперименте на бактериях, «Элементы», 01.02.2012.
2) Пути эволюции предопределены на молекулярном уровне, «Элементы», 12.04.2006.
3) Расширение белковой вселенной продолжается, «Элементы», 24.05.2010.
4) В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011.
-
Интересный эксперимент, но вот его результаты не согласуются с другими. Кроме экспериментов с бактериями и вирусами хотелось бы добавить наблюдения за эволюцией раковых клеток in vivo. Так вот, для развития рака (а это уже не спишешь на какую-то «рыхлость» генома) генетический бэкграунд предка имеет практически определяющее значение.
-
Именно в таких нюансах, на мой взгляд, и кроется различие между "продуктами" абиогенеза и канцерогенеза. Проводить аналогии между клетками опухоли и колонией дрожжей (или бактерий, например) можно лишь в ограниченном контексте, эволюционные предпосылки изначального возникновения этих "продуктов" отличаются. Предки раковых клеток являются одновременно и их субстратом.
-
Любопытно, а можно ли как-то увидеть ссылку(-и) на подобные исследования, в которых говорилось об определяющем (!) значении генетического бэкграунда в новообразованиях. Причем, генетической особенности первичной опухоли, характеризующей не ее (опухоль) наличие и тип (например PML/RARA при М3-ОМЛ, BCR/ABL1 при ОЛЛ и ХМЛ), а той, которая делает опухоль более устойчивой к проводимой терапии (в том числе таргетной). Был бы очень признателен.
Дело в том, что мне встречались подтверждения скорее «глобального эпистаза» - так например, малочисленные клоны опухолевых клеток, несущих мутации абсолютной резистентности к некоторым видам препаратов, выявляющиеся на начальном этапе терапии, не отбираются (как следовало бы ожидать) в процессе терапии этими препаратами, а элиминируются наряду с чувствительными, в подавляющем большинстве случаев (но не во всех!) и не влияют на эффективность проводимой терапии. Резистентность же развивается, скорее, вследствие какой-то общей приспособленности, вызванной разными причинами (высокая скорость элиминации препарата из клетки, множественный сигнальный шунт, гиперэкспрессия онкогенна (-ов), защита мишени действия и т.д.). Спасибо.
-


Понятно, что раз мы тут сидим вполне живые и здоровые, то ничего такого в истории не случалось, жизнь всегда находила правильные решения. Но чисто теоретически возможно ли такое, что жизнь (в широком смысле) не сможет подобрать ключи к своему дальнейшему существованию и исчезнет? Если невозможно (а 3.5 миллиарда лет непрерывного существования жизни пока это подтверждают), то получается, что жизнь - это такое устойчивое состояние материи, которое однажды появившись уже не исчезает.
Ну например, как после Большого Взрыва образовались вещество и силовые поля, так до сих пор и существуют, и с жизнью так же :о)) Но это сравнение, наверное, все же некорректно, однако дает маленькую надежду на то, что жизнь проросла не только на нашей планетке, ее появление тут и там просто-напросто неизбежно :о))
-
Я согласен с тем, что в мире сейчас наблюдается баланс.
При этом не надо строить иллюзий, что живой мир вот такой добрый и пушистый, и в нем, например, есть «мирный» цветок ромашка. На самом деле идет очень жесткая война всех против всех. В этой войне дай волю ромашке, то она заполонит всю землю и не оставит шанса, например, одуванчику.
Постоянная война идет между высшими живыми существами и бактериями и вирусами. Почему одна из сторон не побеждает – видимо потому, что есть сдерживающие факторы. За счет долгой эволюции, например, животные выработали иммунитет к разного рода вирусам и бактериям. При этом «атомные бомбы» среди вирусов были обезврежены уже давно.
Исследование, которое описано в статье, показывает, что процесс адаптации больше напоминает аналоговый процесс – возникла проблема, живые существа приспосабливаются к этой проблеме множеством путей, причем непрерывно, а не скачкообразно. То, что мы еще живы, объясняется тем, что у животных есть эффективная система адаптации защитных механизмов к этим угрозам, которая совершенствовалась сотни миллионов лет. Похоже, что сейчас война между вирусами и животными носит позиционный характер.-
Спасибо всем за ответы, но я немного о другом. Может не так выразился.
Имел в виду такую ситуацию: вообще, имеется ли ненулевая вероятность исчезновения абсолютно всей жизни на планете Земля, начиная от бактерий (да даже вирусов) и заканчивая высшими многоклеточными. Ну допустим в результате какой-либо мутации (или комплекса мутаций) в самой основе белковой жизни, которая еще ни разу не происходила за всю историю и не существует никакого механизма, противостоящего ей.
Другими словами, возможно ли срубить древо жизни под самый корень, да так, что оно больше никогда не сможет вырасти.
Почему такой вопрос возник? По прочтении некоторых книг по эволюции, вышедших в последнее время (в т.ч. и трилогии Александра Маркова, сейчас за Евгения Кунина принимаюсь), сформировался довольно противоречивый образ о развитии белковой жизни на Земле. С одной стороны эволюция вроде бы отбирает самые приспособленные варианты организмов к окружающей среде, а с другой делает это из материалов, которые первые под руку попадутся. И получается что-то эдакое кособокое с кучей подпорок и большим количеством ненужных элементов.
Так вот, не осталось ли в самом основании этого здания еще не проверенной отбором за 3.5 миллиарда лет подпорки, которая безвозвратно сломавшись, потянет за собой все здание? Или все нижележащие подпорки превратились в непробиваемый монолит и их уже не сдвинуть с места?
Вопрос, конечно, слишком общий и может быть наивный :о)) Но каждый раз, когда берусь за очередную книгу по эволюции, он не дает мне покоя.-
Концепции, позволяющие ответить на этот вопрос (точнее, конкретнее его сформулировать), нужно искать уже не в биологии, а в теоретической физике (термодинамика, теория хаоса и диссипативных систем) и философии (что такое жизнь? принципиальным ли является наличие материального носителя генетической информации? неизбежно ли возникновение жизни и сознания?)
Топология фундаментальных (в том числе ещё не открытых) физических законов определяет массы электронов, время жизни протонов; способа объединения их в атомы, в молекулы, в полимеры; определяет первичную, вторичную и n-ую структуру этих полимеров; объединяет звёзды в галактики, галактики в скопления и сверхскопления галактик. У каждой из перечисленных структур (видов организации материи) есть свои граничные пространственно-временнЫе масштабы, за которыми рассматриваемая структура либо мгновенна и точечна, либо вечна и бесконечна. Потому определяющей становится точка зрения наблюдателя (как физической концепции), который своим масштабом (размером и продолжительностью жизни) и определяет вечность или мгновенность того или иного физического явления.
Если попытаться ответить более конкретно, то я бы сформулировал это так: скорее всего, неизвестность продолжительности существования жизни - принципиальное и неотъемлемое (физическое) свойство этой самой жизни. Что-то типа квантовой неопределённости. Вытекает из того, что наблюдатель, пытающийся ответить на этот вопрос, сам является этой самой жизнью. (Кстати, уже упомянутая в обсуждении этой статьи книга "Гёдель, Эшер, Бах" построена как раз вокруг парадоксов автореферентности.)
(Надеюсь, этот псевдонаучный бред хотя бы немного соответствовал заданному вами вопросу.) -
Я думаю, что жизнь сама по себе не исчезнет по следующим причинам.
1. Любая жизнь на Земле организована на единой основе – на основе размножения с использованием известного кода ДНК. Альтернативного варианта нет. Вернее, может быть альтернативная кодировка http://elementy.ru/problems?discuss=839 с использованием других оснований в цепочке ДНК, но эта альтернативная кодировка неэффективна. Базовая организация жизни это, возможно, единственный вариант, который допустила природа для организации живой материи. Вирусы и бактерии это также жизнь, построенная с использование базовых принципов. Поэтому другая жизнь, жизнь, как таковую, уничтожить не может.
С этой точки зрения, чтобы уничтожить жизнь, необходимо изменить законы природы.
2. Я думаю, что микроорганизм, который жил на Земле несколько сотен миллионов (или миллиардов) лет назад не сможет повредить современной жизни, поскольку составляющие этого организма (например, белки) работают менее эффективно.
3. Любая мутация, в результате которой возникло существо с другими характеристиками и которое может уничтожить старую жизнь, является также жизнью, возможно более совершенной. Поэтому жизнь, как таковую, мутация уничтожить не может.
4. Для того,чтобы микроорганизм (вирус, бактерия) мог выиграть войну, необходимо чтобы его конструкция учитывала детальную информацию о противнике. Например, вирус ВИЧ должен «знать», как устроена иммунная система человека. А это может быть только в случае, если вирус или другой организм в своей эволюции очень долго находился с человеком в соприкосновении, и имел опыт «боевых» действий. Организация эффективной «боевой машины» в виде вируса или бактерии при конструировании этой машины с нуля, путем случайной мутации практически невероятна.
5. В принципе, нашу жизнь может уничтожить (или вытеснить) чужая жизнь, организованная на тех-же принципах, что и наша, но которая прошла более долгую и более жесткую эволюцию. Например, инопланетная бактерия. Еще более эффективным терминатором будет сложно организованное существо, например инопланетный клещ или разумное существо.
6. Безусловно, нашу жизнь может уничтожить техногенная жизнь, построенная на основе других (не биологических) принципов.
7. На Земле известны массовые вымирания живых существ. Их, в основном, объясняют ухудшением условий окружающей среды. Я думаю, что есть еще один фактор – это устаревание «конструкции» живых организмов. Например, динозавры вымерли потому, что их конструкция зашла в тупик и они (динозавры) не смогли отвечать требования естественного отбора. Необходима была смена модели.
В общем, я думаю, что жизнь в своем существовании опирается на базовые законы физики. При этом, если существуют подходящие внешние условия (диапазон температур, химический состав окружающей среды и т.д.), то жизнь уничтожить невозможно. Губительным фактором для жизни может быть только изменение физических законов и фундаментальных постоянных.
Я не являюсь специалистом по биологии, поэтому прошу воспринимать написанное как мнение дилетанта.-
А вот скажите: если представить себе универсальный вирус, который смог внедриться во все живые клетки и уничтожить их, можно ли считать, что одна жизнь вытеснила другую? Ведь после гибели клеток вирус неизбежно разрушится и перестанет быть живым по любым критериям (даже если изначально его причисляли к жизни).
Также непонятно про "техногенную жизнь" даже если таковая будет создана: есть сомнение, что ее можно будет причислить к жизни: ведь производство новых существ будет происходить не на принципах самоорганизации материи (где все стадии развития жизни кроме первичного возникновения имеют существенную вероятность),а за счет целенаправленного конструирования, то есть цепочки крайне маловероятных событий. Остается открытым, возможна ли эволюция такой "жизни", ведь для эволюции материальные носители идеи (проекта) существа (допустим, электроны) должны случайным образом образовывать не меньшее разнообразие комбинаций, чем генетический код.
Разного рода идеализм, а именно эволюцию идей без какого-либо материального носителя обсуждать принципиально не буду.-
Для возникновения такого вируса принципиально не может сложиться необходимых факторов отбора, на мой взгляд. Не могу придумать условия, в которых бы было эволюционно выгодно появление и закрепление таких свойств (уничтожение собственного субстрата) в вирусном геноме (принцип "жизнь/обед" по Докинзу, из которого формулируется асимметричность положения убийцы и жертвы и давления отбора на них).
Концепцию небиологической эволюции (возникновение "пост-человека") считаю возможной и даже неизбежной, хотя формальных доказательств такого развития событий на текущий момент не существует: возможность создания так называемого сильного ИИ является всё ещё дискуссионным вопросом.-
Так первый комментарий был не про постепенную эволюцию, а про возможность (крайне маловероятную) единовременного самопроизвольного возникновения такого вируса (существа). Ведь возникла же жизнь самопроизвольно? Почему не может самопроизвольно возникнуть нечто, что связывается с любыми живыми системами (ну или хотя бы с эукариотами/прокариотами допустим), эксплуатируя какие-то самые общие принципы организации жизни, которые не были отвергнуты эволюцией по той причине, что это редкое событие еще ни разу не произошло?
Возможным возражением на это может быть, что возникновение и дальнейшее развитие жизни не есть какое-то одно крайне редкое событие, а есть очень длинная цепочка вполне вероятных событий при определенных условиях. Соответственно, жизнь не имеет общих уязвимых мест.
Согласно гипотезе эволюции молекул-репликаторов, сначала накапливались определенные продукты вполне вероятных реакций и процессов. Вероятность образования каждой молекулы корреллировала с вероятностью ее распада/превращения, то есть никакое вещество/молекула-репликатор не накапливалось неограниченно. В какой-то момент вероятности образования определенных молекул с участием (катализом) самих этих молекул стали расти, но не скачкообразно, а постепенно. Когда вероятность образования определенных сложных молекул существенно превысила вероятность их распада (при наличии "строительных частей"), можно говорить о первом шаге в возникновении жизни. Но каждая мутация (модификация) молекулы увеличивала вероятность успешной репликации по чуть-чуть, поэтому невозможно поделить процесс на "не жизнь", "не вполне жизнь", "почти жизнь"...-
«Так первый комментарий был не про постепенную эволюцию, а про возможность (крайне маловероятную) единовременного самопроизвольного возникновения такого вируса (существа)…»
Для того, чтобы ответить на Ваш вопрос, я напишу как я понимаю возникновение и развитие жизни на Земле.
1. Я считаю, как Вы правильно заметили, что возникновение и дальнейшее развитие жизни не есть какое-то одно крайне редкое событие, а есть очень длинная цепочка вполне вероятных событий при определенных условиях.
Эта цепочка в конечном итоге приводит к единственному варианту организации жизни - на основе известного кода ДНК.
2. Я думаю, что в начале развития, для организации жизни использовались строительные элементы (белки или другие органические молекулы), которые отличались относительно сильными внутримолекулярными и межмолекулярными связями. При этом живые организмы в то время были термофилами и могли жить в широком диапазоне температур.
В связи с тем, что количество возможных комбинаций построения живых существ на основе таких элементов ограничено, за время очень долгой эволюции природа смогла перебрать все варианты без исключения и выбрать наиболее оптимальные.
3. Когда варианты организации живой материи с использованием сильных связей были исчерпаны, в дальнейшем эволюция пошла по пути применения все более слабых связей, которые существенно расширили количество вариантов построения живых организмов. Увеличение количества вариантов позволило эволюции проводить более тонкую оптимизацию, учитывающую различные характеристики внешней среды, а также взаимодействия живых организмов между собой.
В настоящее время, диапазон температур, в котором могут существовать наиболее развитые живые организмы, сузился до 0,5..2 градуса - если температура тела у нас повысилась на 1 градус, то мы уже заболели. В тоже время, за счет сужения температурного диапазона количество вариантов в построении белков и разнообразие их связей увеличилось до очень большой величины. При этом, эволюция белков продолжается и в настоящее время (См. статью «Расширение белковой вселенной продолжается»)
В рамках процесса развития жизни, который я описал, нет места случайности. Поэтому фундамент жизни является монолитным. В связи с этим, я считаю, что вероятность самопроизвольного возникновения вируса, который разрушит основы жизни, практически равна нулю.
Гипотезу, которую я изложил выше, можно проверить. Для этого необходимо провести описанный в статье эксперимент в разных климатических условиях. Например, при повышенной температуре, на грани выживания дрожжей. Если гипотеза верна, результатом эксперимента должно быть резкое снижение приспособляемости дрожжей к другим внешним факторам. Это будет объясняться тем, что мутации будут направлены в сторону использования более сильных межмолекулярных связей, которые более устойчивы к повышению температуры, с одновременным сужением возможности к адаптации к другим внешним факторам. (Т.к. с использованием более сильных межмолекулярных связей сужается количество вариантов, а варианты со слабыми связями, разрушаются температурой.) А при снижении температуры эволюция будет идти. Однако ее темп резко замедлится.
У меня по этому поводу вопрос к Александру Маркову, или к другим специалистам: проводились или нет подобные эксперименты? Возможно, я изобретаю "велосипед".-
" Когда варианты организации живой материи с использованием сильных связей были исчерпаны, в дальнейшем эволюция пошла по пути применения все более слабых связей, которые существенно расширили количество вариантов построения живых организмов. Увеличение количества вариантов позволило эволюции проводить более тонкую оптимизацию, учитывающую различные характеристики внешней среды, а также взаимодействия живых организмов между собой." - А в честь чего она должна идти, если не секрет? Оптимальные варианты отобраны, еда у термофилов всегда под боком, ни один термофил вне термальной обстановки долго не живет. Зачем эволюция? По тому же принципу Докинза "жизнь/обед" нужно сплотиться у кормушки, встроить плотностно-зависимое ограничение численности и жить долго и счастливо.
-
-
-
-
-
«А вот скажите: если представить себе универсальный вирус, который смог внедриться во все живые клетки и уничтожить их..»
Универсально-кровожадных вирусов, по-моему, в природе нет. Есть весьма серьезные вирусы - оспа, чума и т.д. Однако они не привели к глобальному вымиранию всего живого. Например, деревья и ползающие по ним муравьи во время эпидемий вполне себя неплохо чувствовали.
«Также непонятно про "техногенную жизнь"»
Здесь имеется в виду жизнь, созданная человеком или другим разумным биологическим существом. Принцип размножения такой жизни не обязательно может совпадать с принципом размножения биологической жизни. Техногенный организм, также как и биологический может быть построен из универсальных кирпичиков – клеток. Возможно, эти клетки будут существенно большими по размерам и более сложно организованными, чем живые клетки. Эти клетки могли бы синтезироваться в устройствах (органах), чем-то напоминающих продвинутые 3D принтеры, которые могут печатать 3D конструкции с субмикронными топологическими размерами. Подобная цивилизация Репликаторов показана в фильме «Звездные врата- Атлантида».
Хотя, в принципе, от саморазмножения техногенных тел в первое время можно отказаться. Оставить эволюционировать только мозг существа, где эволюция будет сводиться, к совершенствованию архитектуры мозга и его интеллектуальных способностей. А тело будет подстраиваться под эволюцию потом, по итогам эволюции мозга, и изготавливаться во множестве вариантов на заводах. Конкуренция этих вариантов и будет эволюцией тела. Там же может изготавливаться аппаратная часть мозга, в которую потом будут переписываться личности техногенных существ. На мой взгляд, эволюция техногенных существ в таком виде будет более гибкой и эффективной, чем биологическая эволюция. А зоной обитания и ареной эволюции этих существ будет весь космос.-
В том, что техногенная жизнь возможна в части самокопирования, особых сомнений нет. Есть сомнения в том, что она сможет эволюционировать. Ведь эволюция построена на случайных изменениях материальных носителей информации (в данном случае молекул). В техногенной жизни другие носители (электроны, фотоны, магнитные домены etc). Необходимо обеспечить возможности разнообразных преобразований этих носителей и возможности проверки жизнеспособности получившейся комбинации не хуже, чем в случае реальной жизни. Сейчас достигнут очень примитивный уровень как возможностей модификации, так и проверки.
Представим, к примеру, эволюцию генетических алгоритмов, мутацией которых является случайная модификация одной буквы программного кода. Ничего не выйдет, нужны более тонкие механизмы или колоссальные мощности, сравнимые с количеством органических молекул в биосфере Земли.
Или обучение нейросети, жизнеспособность которой проверяется максимизацией соответствий целевого вектора обучающему множеству. В случае реальной жизни длина целевого вектора и размер обучающего множества также может быть сравнима с числом молекул.-
Сама по себе техногенная жизнь на начальной стадии развития жизни, скорее всего, эволюционировать не может. Однако может эволюционировать связка разума (интеллекта, личности, души) и техногенного тела включая аппаратную часть мозга. Для техногенного существа важно со временем сохранить свою личность. Поэтому эволюция души, скорее всего, вредна. А вот техногенное тело можно менять, в соответствии с модой, с профессиональными потребностями, с освоением планет, с обороной своей территории и ее расширением. Эта часть техногенного существа будет эволюционировать. Из множества вариантов будет идти отбор наиболее приспособленных. При этом эволюция будет идти не случайно, и не будут перебираться все случайные варианты, потому, что разум в состоянии смоделировать результаты и отфильтровать все явно неработоспособные варианты. Эволюция будет идти целенаправленно. Это будет «разумная» эволюция.
В данном случае имеется ввиду, что существует множество техногенных существ и имеются множество заводов по производству техногенных тел, принадлежащих конкурирующим сообществам техногенных существ.
Дополнение от 6.07.2014г.
Эволюцию техногенной жизни нельзя рассматривать отдельно от эволюции биологической жизни. Наблюдения показывают, что эволюция очень редко отвергает старое. Например, в своем развитии зародыш человека проходит все стадии от рыбы до человека. Современный персональный компьютер сохранил совместимость по программному коду и некоторым базовым принципам с одним из самых первых персональных компьютеров.
Эволюция человека к техногенному разумному существу, по-видимому, должна проходить подобным образом.
Основу человека составляет не только биологическая оболочка, но и информационное наполнение человека, включая человеческую культуру. В ходе эволюции людей в направлении техногенных разумных существ тело человека может быть заменено на техногенное, однако информационная составляющая человека, по-видимому, должна быть перенесена в техногенное существо и в культуру сообщества техногенных существ без изменений. Иначе возникнут войны. Даже между людьми возникают войны, когда одно сообщество пытается навязать другому сообществу свою культуру. При этом люди другого сообщества воспринимают попытку навязать чужую культуру как попытку убить в них часть своей личности. В связи с этим сообщество техногенных существ, имеющих другую культуру, будет восприниматься людьми как враги.
Из этой проблемы есть выход.
Выход заключается в том, что личность техногенного существа должна формироваться на биологической стадии роста человека. А потом, когда человеческий организм износится, информационная составляющая личности человека должна переписываться в мозг техногенного существа. Это будет продолжение жизни после смерти. Противоречие между культурами здесь нивелируется тем, что техногенные существа будут воспринимать людей как своих детей. Каждый из техногенных существ (или каждый человек с техногенным телом) был в свое время ребенком. А Землю будут воспринимать как свою колыбель. При этом сохраняется преемственность культур.
-
-
-
-
-
1) Хищник устает, а вирус - нет, поэтому балансовая парадигма применима только к вирусу (это и есть пресловутая кривая "хищник-жертва", которая на деле относится не к хищнику, а к паразиту). Все, что нужно от жертвы - это устать позже хищника. Что и наблюдаем в природе на каждой охоте. Баланс численностей не нужен.
2) Никакая эволюция вируса, адаптированного к одному хозяину, не позволит ему перепрыгнуть на другого хозяина, если хозяева не контактируют. Иными словами, кривая "хищник-жертва" априорно действует в пределах конечного круга хозяев. Исчерпав этот круг, вирус "злой волк" погибнет. Что и наблюдаем в природе для каждой эпидемии.
Из сказанного следует, что раз у нас без перерыва существует жизнь, в ней никогда не было единого здания, "кособокого с кучей подпорок и большим количеством ненужных элементов" на едином фундаменте. Общий предок - филогенетическая фикция, любая программа поиска сходства нарисует вам его, не зависимо, был ли он на самом деле. Жизнь сразу возникла разнообразной, общего предка не было, был общий материал, что и доказывает данная статья. Из одного набора можно построить множество зданий. Сломаешь одно, оно упадет на другое, повалит его, но всегда окажется одно, стоящее в стороне, до которого эта волна не дойдет. Такую жизнь вирусам в принципе выкосить невозможно, а если они в ходе эволюции станут хищниками, т.е. станут не пассивно переносится от жертвы к жертве, а активно охотиться на них, они получат усталость как внутренний корректив. Жизни может положить конец только машина, но именно потому, что эволюция работает как "cлепой часовщик", она не создает неустающих машин.
Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки -
 07.09.2015Неадаптивная пластичность ускоряет адаптивную эволюциюАлександр Марков • Новости науки
07.09.2015Неадаптивная пластичность ускоряет адаптивную эволюциюАлександр Марков • Новости науки
-
 23.05.2015Половой отбор защищает от вымиранияАлександр Марков • Новости науки
23.05.2015Половой отбор защищает от вымиранияАлександр Марков • Новости науки
-
 23.03.2015Полиплоидия ускоряет эволюциюАлександр Марков • Новости науки
23.03.2015Полиплоидия ускоряет эволюциюАлександр Марков • Новости науки
-
 03.03.2015Ранние этапы адаптации предсказуемы, поздние — случайныАлександр Марков • Новости науки
03.03.2015Ранние этапы адаптации предсказуемы, поздние — случайныАлександр Марков • Новости науки
-
 30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
-
 23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
-
 23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
-
 25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
-
 01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
Последние новости









Рис. 1. Схема эксперимента. Из единственной предковой гаплоидной клетки (DivAnc) получили 432 линии, которые развивались в одинаковых условиях, но при разной численности популяции в течение 240 поколений (фаза диверсификации, Diversification). Из этих линий были отобраны 64 «основателя» (Founder). От каждого основателя получили по 10 подопытных линий, каждая из которых еще 500 поколений приспосабливалась к тем же условиям при одинаковой (высокой) численности популяции (фаза адаптации, Adaptation). Оттенками зеленого цвета показана приспособленность (чем темнее, тем выше). Рисунок из дополнительных материалов к обсуждаемой статье в Science