Подведены итоги эволюционного эксперимента длиной в 40 000 поколений

В ходе уникального эксперимента, длившегося более 20 лет, удалось детально проследить эволюционные изменения, происходившие в популяции кишечной палочки Escherichia coli в течение 40 000 поколений. В первой половине эксперимента в популяции фиксировались в основном полезные мутации, повышавшие приспособленность бактерий. Самым неожиданным результатом оказалось постоянство скорости накопления полезных мутаций. До сих пор считалось, что с постоянной скоростью должны накапливаться нейтральные мутации, а не полезные, однако в эксперименте всё оказалось наоборот. В середине эксперимента в популяции зафиксировалась мутация, резко повысившая темп мутагенеза. После этого мутации стали фиксироваться на порядок быстрее, но это были в основном уже не полезные, а нейтральные мутации.
Давняя мечта биологов — напрямую сопоставить темпы эволюции на геномном и организменном уровнях — наконец-то начинает сбываться. До сих пор в этой области приходилось довольствоваться в основном теоретическими соображениями и математическими моделями. Уникальный эксперимент, начатый в 1988 году международной командой исследователей под руководством Ричарда Ленски (Richard E. Lenski), дал возможность с небывалой доселе степенью детальности проследить ход эволюции как на уровне генома (темп накопления мутаций), так и на уровне целого организма (темп развития адаптаций).
Эксперимент проводится параллельно с 12 популяциями E. coli, но в обсуждаемой статье рассмотрена только одна из них. Бактерий выращивают на «минимальной» питательной среде (см.: minimal growth medium), в которой лимитирующим фактором, ограничивающим размножение бактерий, является недостаток пищи (глюкозы). Каждый день из пробирки с микробами берут 0,1 мл содержимого и помещают в новую пробирку с 9,9 мл свежей питательной среды. Периодически часть популяции замораживают при –80°C и сохраняют для последующего изучения. Это мудро, потому что аналитические методики — в частности, методики секвенирования («прочтения») геномов — сейчас стремительно развиваются и столь же стремительно дешевеют. На сегодняшний день авторы полностью отсеквенировали геномы бактерий из поколений №0 (предковый штамм), 2000, 5000, 10 000, 15 000, 20 000 и 40 000.
Регулярно проводится также оценка «приспособленности» популяции. Для этого сравнивают скорость размножения «экспериментальных» микробов с контрольным меченым штаммом (о методике оценки приспособленности см. в заметке: Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009).
Длительность эксперимента и размер популяции были достаточными для того, чтобы каждая из возможных точечных мутаций (нуклеотидных замен) в ходе случайного мутирования произошла более одного раза (размер генома подопытного штамма кишечной палочки — 4,6 × 106 пар нуклеотидов).
Чем отличается «возникновение» мутации от ее «закрепления» (фиксации). Далеко не всякая возникшая мутация закрепляется (фиксируется) в популяции. Каждая мутация изначально возникает только у одного микроба. Чтобы мутация зафиксировалась, потомки этого микроба должны вытеснить всех остальных микробов в своей пробирке. Вредная мутация, скорее всего, будет отсеяна отбором. Полезная мутация под действием отбора может закрепиться, то есть распространиться в популяции и достичь стопроцентной частоты, но может и случайно потеряться, пока ее носители еще не успели как следует размножиться. Наконец, нейтральные мутации, согласно имеющимся теоретическим моделям, должны фиксироваться с постоянной скоростью, равной скорости мутирования. Частота встречаемости нейтральной мутации в популяции колеблется в соответствии с алгоритмом случайного блуждания (см. random walk) до тех пор, пока случайно не достигнет величины 0% (полная элиминация) или 100% (фиксация). Парадоксальным образом скорость фиксации нейтральных мутаций не зависит от размера популяции. Это объясняется тем, что чем больше популяция, тем чаще в ней возникают мутации, но и тем ниже вероятность фиксации каждой из них. В результате размер популяции в уравнении просто-напросто сокращается.
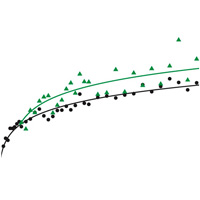
За первые 20 000 поколений в экспериментальной популяции зафиксировалось 45 мутаций, в том числе 29 однонуклеотидных замен и 16 иных мутаций (вставок, выпадений, инверсий, встраиваний мобильных элементов). Самое интересное, что скорость накопления мутаций на этом этапе была строго постоянной (все отклонения от линейной модели статистически недостоверны), тогда как приспособленность сначала росла очень быстро, а затем ее рост замедлился (см. рисунок).
Постоянная скорость фиксации, согласно теории, характерна для нейтральных мутаций. Однако все 45 мутаций не могли быть нейтральными. Ясно, что по крайней мере некоторые из них были полезными — об этом свидетельствует рост приспособленности. Полученные результаты не очень легко увязать и с гипотезой о том, что все 45 мутаций были полезными. Авторы рассматривают несколько простых моделей с разными наборами исходных допущений, из которых следует, что, если бы все мутации были полезными, то обе величины — приспособленность и число накопленных мутаций — должны были бы меняться сходным образом, то есть или расти с постоянной скоростью, или параллельно замедляться.
Простейшее из возможных объяснений состоит в том, что среди 45 зафиксировавшихся мутаций большинство были нейтральными, но некоторые были полезными, причем основная масса полезных мутаций зафиксировалась вскоре после начала эксперимента. Возможности для «полезного мутирования» довольно быстро исчерпались, и в дальнейшем фиксировались преимущественно нейтральные мутации.

Авторы, однако, приводят четыре серьезных аргумента против такого объяснения.
1) В случае преобладания нейтральных мутаций должно быть резко повышено число синонимичных нуклеотидных замен (то есть таких изменений ДНК, которые не ведут к изменениям аминокислотной последовательности белков). Вопреки этим ожиданиям, все без исключения зафиксировавшиеся мутации в кодирующих областях генов являются значимыми (несинонимичными).
2) В случае преобладания нейтральных мутаций следует ожидать, что в 12 экспериментальных популяциях (в 11 из которых полные геномы пока не секвенировались) за 20 000 поколений мутации зафиксировались в разных генах. Напротив, мутации в одних и тех же генах, закрепившиеся независимо в разных популяциях, были бы доводом в пользу того, что мутации фиксировались в результате отбора, а не генетического дрейфа (то есть мутации были полезными). Чтобы проверить это, авторы отсеквенировали у бактерий поколения №20 000 из остальных одиннадцати экспериментальных популяций 14 генов, в которых у «главной» экспериментальной популяции закрепились мутации. Оказалось, что в подавляющем большинстве случаев в других популяциях эти гены тоже изменились.
3) Если бы большинство мутаций были нейтральными, наблюдалась бы значительная внутрипопуляционная изменчивость по этим локусам (потому что полезные мутации под действием отбора фиксируются быстро, а нейтральные сначала должны долго «случайно блуждать» между нулевой и стопроцентной частотой). Это предположение не подтвердилось.
4) При помощи генной инженерии авторы смогли непосредственно определить степень полезности 9 мутаций из рассматриваемых 45. Эти мутации искусственно внедряли в геном предкового штамма. В восьми случаях из девяти приспособленность бактерий резко повысилась. Что касается девятой мутации, то авторы думают, что она тоже полезна, но не сама по себе, а в сочетании с другими мутациями, потому что точно такая же мутация закрепилась у других подопытных популяций. Для сравнения, в другом эксперименте в геном кишечной палочки вносили случайные мутации по одной, и при этом ни одна из 26 мутаций не дала ни малейшего выигрыша в приспособленности.
Таким образом, в течение первых 20 000 поколений в популяции фиксировались преимущественно полезные мутации, причем их фиксация шла с постоянной скоростью. Замедление роста приспособленности, по-видимому, было связано с тем, что средняя степень полезности мутаций постепенно снижалась. Наиболее радикальные адаптивные изменения произошли в течение первых 2000 поколений, а затем, вероятно, происходила более тонкая оптимизация фенотипа.
До сих пор речь шла только о первой половине эксперимента. Во второй его половине эволюционная динамика подопытной популяции радикально изменилась. Дело в том, что после 26 000 поколений зафиксировалась мутация в гене mutT. Этот ген кодирует белок, участвующий в репарации («починке») ДНК. В результате частота мутирования резко выросла. Как следствие, более чем на порядок выросла и частота фиксации мутаций. В течение второй половины эксперимента зафиксировалось 609 мутаций — в 13,5 раз больше, чем за первые 20 000 поколений.
Аналогичные по своей функции мутации, увеличившие темп мутагенеза, закрепились и в нескольких других экспериментальных популяциях. Из этого следует, что рост темпов мутагенеза дал бактериям адаптивное преимущество. Это, между прочим, противоречит распространенной идее о том, что в стабильных условиях организмам было бы выгодно снизить темп мутирования до нуля — и этого не происходит только из-за технической невозможности обеспечить абсолютную точность копирования ДНК (см.: В. П. Щербаков, 2005).
В «основной» популяции эта мутация появилась не позднее, чем в поколении №26 500 (из трех проверенных микробов этого поколения мутация есть у одного и отсутствует у двух). Начиная с поколения №29 000 мутация стала преобладать в популяции и, по-видимому, вскоре после этого зафиксировалась, то есть достигла частоты 100%.
«Полезность» мутации, повысившей темп мутагенеза, могла состоять только в том, что она повысила вероятность возникновения новых полезных мутаций после того, как большинство возможностей для «полезного мутирования» уже было исчерпано. Но в качестве побочного эффекта интенсификация мутагенеза неизбежно должна была привести к росту числа вредных и нейтральных мутаций.
Исходя из сказанного выше, следовало ожидать, что теперь большинство фиксирующихся мутаций будут нейтральными, а не полезными. Как мы помним, скорость фиксации нейтральных мутаций в популяции равна скорости мутагенеза независимо от размера популяции. Скорость мутирования у предкового штамма E. coli была низкой, и поэтому в первые 20 000 поколений нейтральных мутаций фиксировалось очень мало. Мутация гена mutT увеличила скорость мутирования приблизительно в 70 раз (от 1,6 × 10–10 до 1,1 × 10–8 на пару нуклеотидов на поколение). Разнообразные статистические тесты, примененные авторами, подтвердили предположение о том, что большинство из 609 «поздних» мутаций были нейтральными.
Почему-то в статье не сказано, как изменилась приспособленность бактерий во второй половине эксперимента.
Биологи активно используют результаты сравнений геномов для реконструкции путей и темпов эволюции организмов. До сих пор при этом приходилось довольствоваться в основном математическими моделями, основанными на разных более или менее правдоподобных допущениях. Теперь наконец появляется возможность экспериментальной проверки этих моделей. Первые результаты оказались во многом неожиданными. Например, мало кто ожидал, что постоянный темп накопления полезных мутаций может сопровождаться замедляющимся ростом приспособленности; или что возможны столь резкие скачки в соотношении темпов фиксации нейтральных и полезных мутаций. Основной вывод состоит в том, что количественные соотношения между разными аспектами эволюционного процесса (нейтральностью и адаптивностью, влиянием дрейфа и отбора, темпами изменений на уровне генотипа и фенотипа) могут быть существенно более сложными, неоднозначными и переменчивыми, чем предполагалось ранее.
Источник: Jeffrey E. Barrick, Dong Su Yu, Sung Ho Yoon, Haeyoung Jeong, Tae Kwang Oh, Dominique Schneider, Richard E. Lenski, Jihyun F. Kim. Genome evolution and adaptation in a long-term experiment with Escherichia coli // Nature. 2009. V. 461. P. 1243–1247.
Об экспериментальном изучении эволюции см. также:
1) Способность к сложному коллективному поведению может возникнуть благодаря единственной мутации, «Элементы», 25.05.2006.
2) Микробиологи утверждают: многоклеточность — сплошное жульничество, «Элементы», 06.04.2007.
3) Начальные этапы видообразования воспроизведены в эксперименте на дрожжах, «Элементы», 06.06.2007.
4) Для видообразования достаточно одного гена, «Элементы», 28.11.2007.
5) Амёбы-мутанты не позволяют себя обманывать, «Элементы», 06.10.2009.
6) Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009.
-
Да, глобальный подход. Тут ничего не скажешь. На мой взгляд, одним из самых интересных результатов встало проявление по крайней мере одного из механизмов убыстрения мутаций в явном виде. В данном случае, на снятие ранее сформированного механизма ограничения мутаций - как вредных и нейтральных, так даже и полезных ввиду их неразличимости на этапе формирования. Яркое экспериментальное подтверждение в пользу того, что есть периоды "медленной" эволюции с малым изменением генома, и есть скачки, когда мутирование происходит с на порядки большей частотой (в данном случае - 70 раз)
Интересен и факт фиксации исчерпывания резервов возможностей приспособления для изолированной биологической системы. Для дальнейшего приспособления необходим качественный скачек, который, возможно, связан с межвидовым генетическим обменом и, возможно, конечным формированием нового вида. -
Замечательная работа, я только вчера читал ее в оригинале. В принципе это должно войти в учебники по эволюционной биологии (или дарвинизму, как этот недавно еще назывался). Но до закостенелых наших профессоров еще долго это будет доходить. Потому и призываю студентов к САМООБРАЗОВАНИЮ.... А этот сайт будет в подмогу!
-
Насколько бредовой является идея, что популяция представляет собой информационную систему, которая анализирует ситуацию и принимает эволюционные решения, наподобие того, как какой-нибудь нервный ганглий принимает решения по работе управляемого им органа?
Могут ли, например, существовать положительные или отрицательные обратные связи, проявляющиеся через много поколений?
Может ли быть так, допустим, что существует связь, например, что если мутация гена А оказалась успешной, то через N поколений мутабельность гена Б совершенно в другой части популяции уменьшится/увеличится? -
Крайне интересная статья.
На мой взгляд, отсутствие закрепленных нейтральных мутаций в первые 20000 поколений говорит о том, что в условиях эксперимента давление отбора оказывается намного больше фактора генетического дрейфа.
У меня нет сейчас возможности прочитать оригинальную статью, но было бы интересно узнать, использовались ли в эксперименте популяции разного размера. Это позволило бы пронаблюдать зависимость соотношения сил отбора и дрейфа от размера популяции. -
каждое новое поколение увеличивало свою численность в 100 раз, следовательно, если учесть лимитирование по субстрату, то отбор шел при каждом пересеве, что снимает вопрос почему у отобранных клонов выявлены позитивные мутации.
не закрыт вопрос физиологической адаптации за счет перестройки метаболизма без участия мутаций.
далее, частота мутаций в популяции зависит от количества организмов - об этом в статье тоже молчок.
ну и наконец о встраивании мобильных элементов - откудова они взялись в закрытой то системе?


Сотни накопленных мутаций ни на йоту не сдвинули таксономическое положение бактерии...
Вывод - обычная идеологическая профанация! :о)
Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки -
 30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
-
 23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
-
 23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
-
 25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
-
 01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
-
 25.03.2011В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность»Александр Марков • Новости науки
25.03.2011В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность»Александр Марков • Новости науки
-
 01.11.2009Подведены итоги эволюционного эксперимента длиной в 40 000 поколенийАлександр Марков • Новости науки
01.11.2009Подведены итоги эволюционного эксперимента длиной в 40 000 поколенийАлександр Марков • Новости науки
-
 23.10.2009Опыты на червях доказали, что самцы — вещь полезнаяАлександр Марков • Новости науки
23.10.2009Опыты на червях доказали, что самцы — вещь полезнаяАлександр Марков • Новости науки
-
 02.01.2008Эволюционная роль мобильных генетических элементов остается загадочнойАлександр Марков • Журнал общей биологии • №5, 2007
02.01.2008Эволюционная роль мобильных генетических элементов остается загадочнойАлександр Марков • Журнал общей биологии • №5, 2007
Последние новости