Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает расти

Долгосрочный эволюционный эксперимент на бактериях, начатый Ричардом Ленски в 1988 году, продолжает приносить интересные результаты. Как выяснилось, по прошествии 50 000 поколений (столько сменилось к февралю 2010 года) подопытные бактерии всё еще продолжают накапливать полезные мутации, неуклонно повышая свою приспособленность к условиям, в которые их поместили экспериментаторы. Рост приспособленности постепенно замедляется, но, судя по всему, не собирается выходить на плато. В шести из 12 подопытных популяций закрепились мутации, резко повысившие темп мутагенеза, что способствовало более быстрому росту приспособленности.
«Элементы» уже рассказывали о долгосрочном эволюционном эксперименте (ДЭЭ) на кишечных палочках Escherichia coli, который проводится в Мичиганском университете вот уже четверть века (см.: E. coli Long-term Experimental Evolution Project и ссылки в конце новости). За это время в колбах с 12 популяциями подопытных бактерий, происходящими от одного и того же предкового штамма, сменилось более 59 000 поколений микробов (каждые 75 дней сменяются примерно 500 поколений). Пока это единственная в мире экспериментальная система, позволяющая в деталях проследить эволюционные изменения в большой популяции на таком длительном интервале времени.
Система предельно упрощена по сравнению с природными сообществами микроорганизмов. Во-первых, бактерии размножаются в монокультуре, что позволяет абстрагироваться от межвидовых взаимодействий. Во-вторых, питательная среда является «бедной», в ней мало пищи (глюкозы), поэтому плотность микробного населения в колбах остается низкой: это позволяет минимизировать влияние бактерий друг на друга посредством выделения тех или иных веществ. В-третьих, популяции являются «бесполыми»: они лишены средств для горизонтальной передачи генов, поэтому гены в подопытных популяциях передаются только вертикально (от родителей к потомкам).
С одной стороны, можно сказать, что всё это делает результаты эксперимента мало приложимыми к реальным эволюционным процессам. С другой стороны, эти упрощения позволяют получать однозначно интерпретируемые результаты, а целью эксперимента является изучение самых фундаментальных эволюционных процессов (мутаций, отбора, дрейфа, адаптации к среде) в максимально чистом, идеализированном виде. «Сложности» можно будет добавлять потом, по мере необходимости, когда станут понятны основы.
Недавно Ленски и его коллеги сообщили об очередном важном результате, полученном в ходе ДЭЭ. На этот раз ученые сосредоточились на динамике роста приспособленности в 12 подопытных популяциях. «Приспособленность» оценивалась как скорость размножения бактерий в стандартных условиях ДЭЭ по сравнению с предковым штаммом (см. новость Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 02.11.2009). Напомним, что предковый штамм сохраняется в замороженном состоянии, причем его в любой момент можно разморозить и использовать в опытах. Кроме того, каждые 500 поколений замораживаются образцы микробов из каждой эволюционирующей популяции. В итоге получается «живая ископаемая летопись», позволяющая проследить все эволюционные события с большой подробностью и даже «проиграть» те или иные события повторно, чтобы отделить случайности от закономерностей.
В трех из 12 подопытных популяций в ходе ДЭЭ произошли настолько радикальные эволюционные изменения, что сравнивать их приспособленность с другими линиями стало затруднительно. В одной из популяций развилась способность использовать в пищу цитрат, что привело к резкому росту плотности популяции (см.: В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества, «Элементы», 25.09.2012), а две другие разучились образовывать колонии на агаре. Из-за этого авторы не смогли применить к ним стандартную методику оценки относительной скорости роста в «соревновательных» тестах. Поэтому поздние этапы эволюции этих трех популяций были исключены из анализа (а ранние этапы учитывались). Между прочим, во всех трех исключенных популяциях закрепились аллели-мутаторы, то есть мутации, повысившие темп мутагенеза (см. ниже).
Ранее было показано, что в отдельных популяциях на протяжении 20 000 поколений приспособленность росла с замедлением. Однако оставалось неясным, собирается ли приспособленность вскоре выйти на плато или ее рост, пусть и замедляющийся, будет продолжаться еще долго.
Для начала авторы сравнили приспособленность бактерий из поколений №№ 40 000 и 50 000 в девяти популяциях. Оказалось, что у шести популяций приспособленность достоверно выросла на этом интервале, у трех улучшение «не дотянуло» до порога статистической значимости. В среднем подопытные популяции повысили свою приспособленность на целых три процента. Таким образом, даже после 40 000 поколений процесс адаптации бактерий к условиям ДЭЭ не прекратился: микробы все еще продолжают накапливать полезные мутации и повышать свою приспособленность.
Затем авторы проследили траекторию роста приспособленности в подопытных популяциях, заставляя замороженных на разных этапах ДЭЭ микробов соревноваться с предками. Результаты, усредненные по всем 12 популяциям, представлены на рис. 1. Анализ полученных графиков показал, что динамика роста приспособленности лучше описывается степенной моделью w = (bt + 1)a, чем гиперболической w = 1 + at/(t + b), где w — средняя приспособленность, t — время в поколениях, a и b — подбираемые параметры. Степенная модель предполагает неограниченный, хотя и замедляющийся (при a < 1) рост приспособленности, тогда как гиперболическая модель соответствует асимптотическому приближению приспособленности к максимально возможному значению (1 + a). На рис. 1, B видно, что степенная функция, параметры которой были подобраны на основе данных по первым 20 000 поколений, весьма точно предсказывает динамику приспособленности на протяжении следующих 30 000 поколений, тогда как предсказания гиперболической модели сильно «недооценивают» реальный рост приспособленности в будущем.
Итак, рост приспособленности в ДЭЭ пока не собирается выходить на плато. Несмотря на долгую жизнь в одних и тех же условиях, у бактерий всё еще время от времени закрепляются новые полезные (= повышающие приспособленность к этим условиям) мутации. Удивительная неисчерпаемость адаптационных возможностей, впервые столь наглядно продемонстрированная в эксперименте, имеет огромное значение для понимания эволюции. Ведь можно было предположить (многие так и думали), что в постоянных условиях, тем более в монокультуре, возможности для адаптивной эволюции быстро будут исчерпаны, и система придет в состояние полного стазиса. В таком случае объяснить продолжающуюся эволюцию жизни на нашей планете можно было бы только через постоянные изменения среды (абиотической и биотической).
Авторы допускают, что запас потенциально полезных мутаций, возможно, не иссякнет не только за 50 000 поколений бактерий, сменившихся на глазах одного исследователя, но и за 50 000 поколений исследователей. Как ни странно, это не приведет к какому-то невероятному, запредельному росту приспособленности. Ученые рассчитали, что если рост приспособленности будет и впредь идти по той же траектории, описываемой степенной функцией, то пятидесятитысячный по счету преемник Ленски увидит бактерий, удваивающих свою численность каждые 23 минуты. Это высокая скорость размножения для бактерий, но вовсе не какая-то неслыханная (предковый штамм удваивает свою численность в условиях ДЭЭ за 55 минут).
Откуда же берутся новые полезные мутации? Ведь за время эксперимента каждая возможная нуклеотидная замена уже наверняка успела произойти хотя бы у одной бактерии! Основных причин, по-видимому, две. Первая связана с эпистазом — взаимодействием между мутациями. Например, мутация Б становится полезной только после того, как бактерия уже обзавелась мутацией А. Здесь нужно помнить еще и о том, что многие полезные мутации имеют негативные побочные эффекты. Мутация А может улучшить работу одной молекулярной системы, немного повредив другой. Допустим, первый эффект сильнее второго, поэтому в суммарном зачете мутация А оказывается «полезной» и поддерживается отбором. Но теперь, когда в популяции закрепилась мутация А, окажется полезной любая компенсаторная мутация Б, устраняющая или сглаживающая негативный побочный эффект мутации А.
Вторая причина «неисчерпаемости» запаса полезных мутаций состоит в том, что мутации с очень слабым полезным эффектом требуют очень долгого времени для закрепления в популяции. Такая мутация, появившись, с большой вероятностью будет вскоре потеряна из-за генетического дрейфа — случайных колебаний частот аллелей. Чтобы отбор начал хоть немного «помогать» такой мутации распространяться, число ее носителей должно стать довольно большим, а до тех пор мутация будет находиться целиком под властью безжалостного дрейфа. Расчеты показывают, что подавляющее большинство вновь возникающих полезных мутаций теряются. Чтобы данная мутация в конце концов все-таки закрепилась, она должна независимо появиться у разных бактерий множество раз. Авторы рассчитали, что мутация, повышающая приспособленность на одну миллионную, должна возникнуть в среднем 250 000 раз, прежде чем она, наконец, распространится и зафиксируется в популяции. С учетом частоты мутирования микробов, участвующих в ДЭЭ (примерно одна мутация на 1010 нуклеотидов за поколение), и размера подопытных популяций (эффективная численность каждой из них, с учетом ежедневных «бутылочных горлышек», составляет примерно 33 000 000 особей) получается, что такой мутации понадобится в среднем 100 000 000 поколений, чтобы «спастись» от дрейфа, и еще миллионы поколений, чтобы зафиксироваться (достичь 100-процентной частоты). Таким образом, накопление слабополезных мутаций в ДЭЭ вряд ли закончится в обозримом (и даже необозримом) будущем.
Причины замедления роста приспособленности, по мнению авторов, связаны с тем, что каждая закрепившаяся полезная мутация делает последующие генетические усовершенствования в среднем менее полезными. Авторы называют этот эффект «эпистазом убывающей доходности», diminishing returns epistasis, по аналогии с экономическим законом убывающей доходности. Иными словами, сначала происходят наиболее радикальные адаптивные изменения, а затем идет все более тонкая оптимизация фенотипа.
Еще один фактор, способствующий замедлению роста приспособленности и тесно связанный с предыдущим, — клональная интерференция, то есть конкуренция между клонами бактерий с разными полезными мутациями. Напомним, что бактерии, участвующие в эксперименте, — бесполые, они не могут обмениваться генами. Поэтому, если у одной бактерии возникнет одна полезная мутация, а у другой — другая, эти мутации не смогут объединиться в одном геноме (как это произошло бы у нормальных микробов, способных к горизонтальному переносу генов, и уж тем более у организмов, размножающихся половым путем). Вместо этого потомки первой бактерии начнут не на жизнь, а на смерть конкурировать с потомками второй. Победит, естественно, тот клон, чья мутация окажется более полезной. Вторая, менее полезная мутация будет вытеснена и потеряна. Придется теперь ждать, пока она случайно появится снова, а закрепиться она сможет, только если у нее не окажется более удачливых конкурентов. Клональная интерференция во многом объясняет, почему на первых этапах эксперимента фиксировались в основном мутации с сильным полезным эффектом (в среднем первая закрепившаяся в каждой популяции мутация повышала приспособленность на 10% — это очень много), а затем уже постепенно начинали фиксироваться все менее и менее полезные мутации.
Разумеется, в популяции бактерий, способных к горизонтальному переносу генов, адаптация шла бы гораздо быстрее. Но и анализировать результаты было бы неизмеримо труднее, потому что к мутационной изменчивости добавилась бы комбинативная, а на эволюцию стали бы влиять такие факторы, как генетически обусловленные различия по склонности к приему и передаче генов, избирательность при выборе партнеров для конъюгации и т. д.
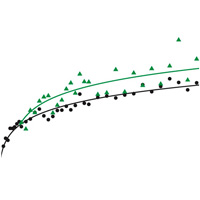
Еще один важный результат авторы получили, сравнив динамику роста приспособленности у двух групп подопытных популяций: у тех, что сохранили исходный (низкий) темп мутагенеза, и тех, где закрепились аллели-мутаторы, то есть мутации, резко (в среднем на два порядка) повысившие темп мутирования (см.: Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 02.11.2009). На сегодняшний день «гипермутаторами» стали шесть из 12 популяций. Из этих шести использовались для сравнения данные только по трем популяциям, в которых аллели-мутаторы закрепились в течение первых 20 000 поколений (включая одну, чью приспособленность удалось оценить только для первых 30 000 поколений). Данные по остальным трем популяциям, поздно вставшим на путь ускоренного мутирования, не учитывались (две из них не учитывались также и в тестах, описанных выше).

Рис. 2. Повышение темпа мутагенеза ускоряет адаптацию. Черные точки — усредненные данные по шести популяциям, в которых темп мутагенеза остался на исходном низком уровне. Зеленые треугольники — усредненные данные по трем популяциям, в которых на ранних этапах ДЭЭ (в первые 20 000 поколений) закрепились аллели-мутаторы. Видно, что приспособленность у вторых росла быстрее, чем у первых. Рисунок из обсуждаемой статьи в Science
Но в трех популяциях, вставших на этот путь рано, адаптация явно шла ускоренными темпами (рис. 2). Повышенный темп мутагенеза пошел бактериям на пользу. С одной стороны (как говорилось в процитированной выше новости), это противоречит распространенной идее о том, что в стабильных условиях организмам было бы выгодно снизить темп мутирования до нуля — и этого не происходит только из-за технической невозможности обеспечить абсолютную точность копирования ДНК. С другой стороны, большинство вновь возникающих мутаций все-таки вредны, поэтому мутация, повышающая темп мутагенеза, в краткосрочной перспективе приносит в среднем больше вреда, чем пользы, и отбор должен действовать против нее. Однако в бесполой популяции мутаторы часто спасаются от отбраковки благодаря эффекту, который называют «генетическим автостопом» (см. genetic hitchhiking). Если какой-то гипермутабельной бактерии повезет и у нее появится полезная мутация, отбор начнет поддерживать последнюю, причем, вместе с полезной мутацией будет автоматически распространяться и сидящий на той же хромосоме аллель-мутатор. Он будет, подобно пассажиру-безбилетнику, ехать верхом на хромосоме, распространение которой в генофонде будет «оплачиваться» полезной мутацией.
В популяции, способной к генетическому обмену, ситуация, скорее всего, будет иной. Связка мутатора с полезной мутацией не будет неразрывной, что позволит отбору работать по отдельности с каждой мутацией (а не с целыми неразделимыми геномами). В итоге отбор сможет закрепить полезную мутацию и отбраковать «породивший» ее аллель-мутатор. Но всё это пока лишь теории: долгосрочный эволюционный эксперимент на организмах, способных к генетическому обмену, еще не поставлен.
Источник: Michael J. Wiser, Noah Ribeck, Richard E. Lenski. Long-Term Dynamics of Adaptation in Asexual Populations // Science. 2013. V. 342. P. 1364–1367.
См. также:
1) Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 02.11.2009.
2) В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011.
3) В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества, «Элементы», 25.09.2012.


> отбор сможет закрепить полезную мутацию и отбраковать
> «породивший» ее аллель-мутатор.
Что-то не верится. Если мутатор сам по себе не снижает жизнеспособность организма, то он сам по себе не будет отсеиваться. Отсеиваться будет он в том случае, если потомство в среднем будет менее плодовитым. А среднее тут посчитать сложно, т.к. в 99.9999% случаев потомок-"мутант" будет менее жизнеспособен, но оставшиеся 0.0001% потомков-мутантов будут невероятно более плодовиты как раз по причине удачных мутаций в их предках, и эти доли процента могут перевешивать. Как итог, я ожидаю точно таких же результатов: мутаторы будут ускорять эволюцию и будут закрепляться. Потом, когда де-факто произойдёт выход на плато, мутаторы будут вредны (будут слишком уж часто ломать и слишком уж редко делать пользу) и отсеются. Полагаю что в реальной природе мутаторы не закрепляются как раз по причине практически бесконечного времени эволюции - фактическое плато давно достигнуто.
-
1)Вы не учитывайете (а А.Марков здесь не повторяет как общеизвестное - пройдитесь по всем сообщениям об ДЭЭ Ленски), что генетический обмен позволяет ФИЗИЧЕСКИ разорвать связь между генами за счет рекомбинации. Рекомбинация может перебросить ген-мутатор в геном одного потомка, а порожденную им полезную мутацию в геном другого потомка. И все, их пути разошлись, генетический автостоп разрушен. А вот насчет того, что долговременный эволюционный эксперимент на животных с рекомбинацией не поставлен, А.Марков ошибается - его поставил японский генетик Мукаи еше в 60-70 гг. на дрозофиле и показал, что на хромосоме в ближайших окрестностях полезных генов накапливаются вредные мутации. Полезный ген они не затрагивают, но сидят так близко, что рекомбинация просто не может разорвать эту связь - более вероятными при кроссинговере оказываются перестройки в полезном гене, с непредсказуемыми, естественно, последствиями.
2) "Как итог, я ожидаю точно таких же результатов: мутаторы будут ускорять эволюцию и будут закрепляться" - только в большой популяции в комфортных или субкомфортных медленно изменяющихся условиях среды. Все забывают, что речь идет о БОЛЬШОЙ популяции и именно о такой среде. Даже А.Марков ("Повышенный темп мутагенеза пошел бактериям на пользу" - правильно - Повышенный темп мутагенеза пошел бактериям на пользу в БОЛЬШОЙ популяции в комфортных или субкомфортных медленно изменяющихся условиях среды). В иных условиях (малая популяция и высокоизменчивая среда, малая популяция и очень опасная среда) высокий темп мутирования равносилен самоубийству, потому, что вредных мутаций больше, чем полезных.
-
Re: Мутации к приспособленности не имеют никакого отношения
Любая мутация может влиять на приспособленность. Если в результате мутации особь, к примеру, погибла в раннем возрасте, не оставив потомства, значит вредная это была мутация, и наоборот. Есть и нейтральные мутации. Но то, что влияние мутаций на приспособленность имеет место быть, это так по определению. Вы уж разберитесь с базовой терминологией, для начала.
Re: Одну модель такой обратной связи я предложил.
Творите дальше, по своему адресу www.bio-foton.ru, а здесь лучше не надо. А еще лучше - пишите сразу в Nature.-
А вообще по Вашей терминологии это что мутация? Добавлена к ДНК одна пара нуклеотидов, добавлен триплет, кодирующий аминокислоту или ничего не кодирующий, или изменена энергетическая конфигурация молекулы (Вы понимаете это терминологию или это для Вас абракадабра?)или еще что-то? И как оно по Вашему все добавляется? Прилепляется как жвачка? Или происходят какие-то реакции? Под управлением чего? Может быть Вы знаете как молекулы узнают друг друга? Под воздействием каких сил они сближаются? Я могу с Вами спорить что ничего этого не знаете и если не попадете на плохой для Вас сайт, то и не узнаете. Любой креационист Вас посадит в лужу.
-
Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки -
 23.05.2015Половой отбор защищает от вымиранияАлександр Марков • Новости науки
23.05.2015Половой отбор защищает от вымиранияАлександр Марков • Новости науки
-
 23.03.2015Полиплоидия ускоряет эволюциюАлександр Марков • Новости науки
23.03.2015Полиплоидия ускоряет эволюциюАлександр Марков • Новости науки
-
 03.03.2015Ранние этапы адаптации предсказуемы, поздние — случайныАлександр Марков • Новости науки
03.03.2015Ранние этапы адаптации предсказуемы, поздние — случайныАлександр Марков • Новости науки
-
 30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
30.06.2014Предсказуемый рост приспособленности достигается непредсказуемыми путямиАлександр Марков • Новости науки
-
 23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
23.12.2013Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает растиАлександр Марков • Новости науки
-
 23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
23.10.2012Процесс появления новых ферментов прослежен в эволюционном экспериментеАлександр Марков • Новости науки
-
 25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
25.09.2012В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшестваАлександр Марков • Новости науки
-
 01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
01.02.2012Параллельная эволюция изучена в эксперименте на бактерияхАлександр Марков • Новости науки
-
 25.03.2011В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность»Александр Марков • Новости науки
25.03.2011В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность»Александр Марков • Новости науки
Последние новости









Рис. 1. Рост приспособленности бактерий в течение 50 000 поколений. Показаны усредненные значения по всем 12 подопытным популяциям. По вертикальной оси: относительная приспособленность (скорость роста по сравнению с предковым штаммом). По горизонтальной оси: время в поколениях. Красные линии — гиперболическая модель (приспособленность асимптотически стремится к предельному уровню), синие линии — степенная модель (неограниченный, хотя и замедляющийся, рост приспособленности). A — параметры кривых подобраны на основе данных по всем 50 000 поколений; превосходство степенной модели над гиперболической не очевидно (хотя и выявляется статистическими методами), B — кривые построены для первых 20 000 поколений, пунктиром показаны «предсказания»; видно, что гиперболическая модель хуже предсказывает динамику приспособленности в течение последующих 30 000 поколений, чем степенная. Рисунок из обсуждаемой статьи в Science