Геномы новооткрытых цианобактерий свидетельствуют о позднем появлении кислородного фотосинтеза

Считается, что атмосфера Земли стала кислородной 2,4–2,3 миллиарда лет назад благодаря цианобактериям, освоившим окcигенный фотосинтез. При этом датировки таких важнейших эволюционных событий, как появление цианобактерий, оксигенного фотосинтеза и кислородного дыхания остаются спорными. Современные методы метагеномного анализа, открывшие окно в мир некультивируемых микробов, позволили обнаружить два новых класса нефотосинтезирующих цианобактерий, Melainabacteria и Sericytochromatia, вдобавок к давно известным фотосинтезирующим цианобактериям (Oxyphotobacteria). Сравнительный анализ геномов показал, что первые цианобактерии не имели ни фотосинтеза, ни кислородного дыхания. Белки, необходимые для фотосинтеза, появились у предков Oxyphotobacteria после их отделения от Melainabacteria, которое произошло 2,6–2,5 миллиардов лет назад. Затем представители всех трех классов независимо приобрели белковые комплексы, необходимые для кислородного дыхания. Горизонтальный перенос генов играл в этих событиях важнейшую роль.
Появление оксигенного фотосинтеза преобразило земную биосферу и создало предпосылки для эволюции высших форм жизни. Единственными организмами, способными к оксигенному фотосинтезу, являются цианобактерии и их прямые потомки — хлоропласты фотосинтезирующих эукариот. Поэтому для понимания эволюции земной жизни необходимо как можно точнее датировать появление цианобактерий и кислородного фотосинтеза. Несмотря на повышенный интерес к данной проблеме, она до сих пор окончательно не решена (см. ссылки в конце новости). Трудности связаны в частности с тем, что геологическая летопись дает в основном лишь косвенные и трудно интерпретируемые данные об эволюции докембрийских прокариот.
К счастью, геологическая летопись — не единственный источник сведений о давних эволюционных событиях. Не менее важным источником сегодня стала сравнительная геномика, позволяющая реконструировать не только хронологический порядок событий, но и детали молекулярного устройства давно вымерших микробов путем сравнения ДНК их современных родичей.
В последние годы благодаря развитию метагеномики в поле зрения ученых попало множество микробов, не желающих расти на искусственных средах в лаборатории, что не позволяло их изучить (а подчас и обнаружить) классическими методами микробиологии. Среди новооткрытых микробов нашлись переходные формы, пролившие неожиданно яркий свет на величайшую загадку эволюционной биологии — происхождение эукариот (см.: Описан новый надтип архей, к которому относятся предки эукариот, «Элементы», 16.01.2017).
Статья австралийских и американских биологов, опубликованная 31 марта в журнале Science, сообщает об аналогичном прорыве в изучении другого важнейшего эволюционного события, или, точнее, комплекса событий — появления цианобактерий, кислородного фотосинтеза и аэробного дыхания.
Ранее уже было показано, что к типу цианобактерий, помимо давно известных фотосинтезирующих форм (рис. 1), относится ряд микробов, не способных к фотосинтезу. Часть их была выделена в класс Melainabacteria. Эти микроорганизмы встречаются в озерах, подземных и сточных водах, а также в человеческом кишечнике. Мелаинабактерии образуют монофилетическую группу с фотосинтезирующими цианобактериями. Иными словами, мелаинабактерии и фотосинтезирующие цианобактерии являются единственными современными потомками своего последнего общего предка. Фотосинтезирующие цианобактерии вместе с произошедшими от них хлоропластами тоже образуют монофилетическую группу, сестринскую по отношению к мелаинабактериям, которая получила название Oxyphotobacteria (R. M. Soo et al., 2014. An Expanded Genomic Representation of the Phylum Cyanobacteria). Кроме того, анализ генов 16S рРНК из различных сред выявил третью, самую базальную ветвь цианобактерий (сестринскую по отношению к Melainabacteria + Oxyphotobacteria). Полные геномы представителей этой третьей группы до сих пор не были известны, и поэтому она фигурировала в литературе под невразумительным названием «клада ML635J-21».
В своей новой статье Рошель Соо (Rochelle M. Soo) и ее коллеги из Квинслендского университета в Брисбене (Австралия) и Калифорнийского технологического института (Пасадена, США) сообщили о сборке черновых геномов трех представителей клады ML635J-21, которую они переименовали в класс Sericytochromatia (от лат. sero — «поздно» и «цитохром», что указывает на позднее приобретение этой группой цитохромов, необходимых для аэробного дыхания, см. ниже). Представители нового класса Sericytochromatia обнаружены как в освещенных местообитаниях (водорослевые биопленки), так и в темноте (подземные воды, угольные пласты). Кроме того, авторы собрали 28 новых черновых геномов Melainabacteria и переопределили 10 ранее собранных геномов, которые были ошибочно отнесены к другим группам, а на самом деле относятся к мелаинабактериям.
В итоге в распоряжении исследователей оказался большой массив геномных данных, достаточный для содержательного анализа ранних этапов эволюции цианобактерий.

Рис. 2. Эволюционное дерево цианобактерий, основанное на генах 120 консервативных белков. Дерево включает три ветви, соответствующие классам Oxyphotobacteria, Melainabacteria и Sericytochromatia. Разноцветными значками показаны моменты приобретения белков оксигенного фотосинтеза (зеленые звездочка и шестиугольник) и кислородного дыхания (компоненты цепи переноса электронов: Complex III — протонная помпа, перекачивающая протоны через клеточную мембрану и задействованная в обоих процессах — фотосинтезе и дыхании; Complex IV — белковый комплекс, передающий электрон кислороду и задействованный только в дыхании); красный крестик — утрата цепи переноса электронов в одной из групп мелаинабактерий. Разными оттенками показаны белки со сходной функцией, связанные лишь очень отдаленным родством. Рисунок из обсуждаемой статьи в Science
Получившееся эволюционное дерево показано на рис. 2. Новые данные подтвердили деление цианобактерий на три класса и базальное положение Sericytochromatia. Судя по распределению на этом дереве белков, необходимых для фотосинтеза (фотосистемы I и II, ферменты цикла Кальвина), первые цианобактерии не были способны ни к фотосинтезу, ни к фиксации CO2. По-видимому, они были гетеротрофами (потребителями готовой органики). Между прочим, среди мелаинабактерий есть даже хищники, что является большой редкостью в мире бактерий (см.: Vampirovibrio chlorellavorus).
Ничего похожего на белки фотосистем и цикла Кальвина нет ни у одного из представителей Melainabacteria и Sericytochromatia. По-видимому, эти белки (точнее, их гены) были позаимствованы путем горизонтального переноса у каких-то других бактерий и собрались в систему оксигенного фотосинтеза у предков Oxyphotobacteria уже после их отделения от предков Melainabacteria. Судя по молекулярным часам, эти группы разошлись в самом конце архейского эона, 2,6–2,5 миллиардов лет назад (P. M. Shih et al., 2016. Crown group Oxyphotobacteria postdate the rise of oxygen). Получается, что кислородный фотосинтез появился незадолго до периода быстрого роста концентрации кислорода в атмосфере, который имел место 2,4–2,3 млрд лет назад.
Новые данные позволили также уточнить имеющиеся представления об эволюции аэробного дыхания у цианобактерий. Для начала заметим, что оксигенный фотосинтез и аэробное дыхание — процессы, связанные не только по своему результату (первый производит органику и кислород из CO2 и воды, второй превращает органику и кислород обратно в CO2 и воду), но и по происхождению. Ключевую роль в обоих процессах играют электронно-транспортные цепи (ЭТЦ; см. Electron transport chain), состоящие из нескольких вмонтированных в мембрану белковых комплексов. Детальные схемы этих цепей можно посмотреть на сайте KEGG (ЭТЦ оксигенного фотосинтеза, ЭТЦ аэробного дыхания).
Некоторые компоненты фотосинтетической и дыхательной ЭТЦ явно имеют общее происхождение. В первую очередь это относится к так называемому комплексу III (Complex III), который использует энергию передаваемого от белка к белку возбужденного электрона для перекачки протонов с одной стороны мембраны на другую. Получившийся электрохимический градиент затем используется для синтеза АТФ при помощи АТФ-синтазы: этот процесс одинаков при фотосинтезе и дыхании.
Родство молекулярных систем оксигенного фотосинтеза и аэробного дыхания особенно наглядно проявляется у фотосинтезирующих цианобактерий, у которых ряд компонентов ЭТЦ, включая комлекс III, задействован одновременно в обоих процессах (рис. 3).
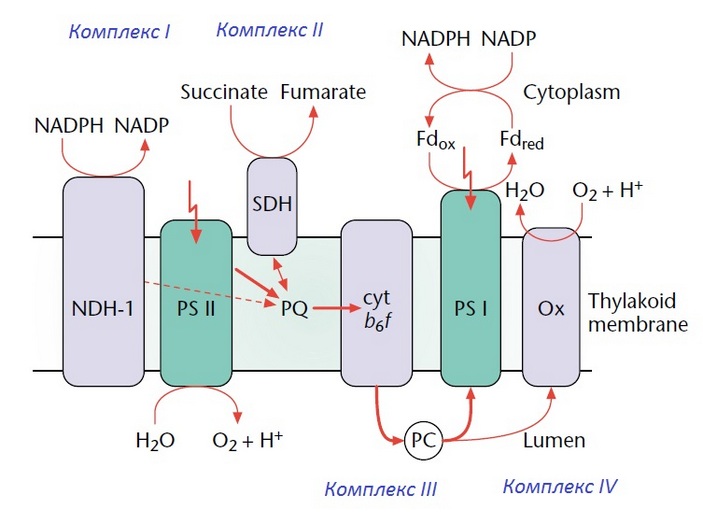
Рис. 3. Схема электронно-транспортных цепей фотосинтеза и дыхания у цианобактерии Synechocystis. Зеленым цветом выделены белковые комплексы, участвующие только в фотосинтезе (PS I, PS II — фотосистемы I и II, содержащие хлорофилл), серым — компоненты дыхательной цепи (комплексы I–IV). Некоторые блоки (PQ — пластохинон, комплекс III, PC — пластоцианин) задействованы одновременно и в фотосинтезе, и в дыхании. Рисунок из статьи W. F. J. Vermaas, 2001. Photosynthesis and Respiration in Cyanobacteria
Если бы первые цианобактерии имели кислородный фотосинтез или дыхание, то у них должны были быть соответствующие белки, включая цитохромы комплекса III, которые затем переходили бы по наследству к их потомкам. В таком случае эволюционные деревья, построенные по отдельности для этих цитохромов и других «дыхательных» белков, совпадали бы с деревом, построенным на основе всего генома и отражающим родственные связи классов цианобактерий.
Ничего подобного, однако, не наблюдается. Дыхательные белки цианобактерий демонстрируют сходство с соответствующими белками самых разных групп бактерий (деревья, построенные для отдельных белков, приведены в дополнительных материалах к обсуждаемой статье). Из этого следует, что, как показано на рис. 2, разные группы цианобактерий приобретали белки дыхательной ЭТЦ независимо друг от друга, в разное время и от разных доноров. Никакой эволюционной преемственности в этом отношении между классами цианобактерий нет. Системы кислородного дыхания явно формировались за счет горизонтального переноса генов от других (причем разных) групп бактерий. При этом одни цианобактерии обзавелись белками, подходящими для дыхания при низких концентрациях кислорода, другие — при высоких.
Таким образом, у первых цианобактерий не было не только оксигенного фотосинтеза, но и кислородного дыхания. В небольших количествах свободный кислород мог производиться и за счет абиотических процессов, таких как фотолиз водяного пара, но этого, очевидно, было недостаточно, чтобы стимулировать эволюцию кислородного дыхания до появления оксигенного фотосинтеза. После этого события многим микробам пришлось срочно приспосабливаться к присутствию кислорода, токсичного для неадаптированных к нему организмов. В ходе этой адаптации горизонтальный перенос генов играл ключевую роль, а дыхательные ЭТЦ развивались как модификации сложившихся ранее фотосинтетических ЭТЦ.
Один из главных выводов работы состоит в том, что цианобактерии начали производить кислород лишь 2,6–2,5 млрд лет назад или даже позже. Этот вывод противоречит распространенной точке зрения о раннем (около 3 млрд лет назад) начале кислородного фотосинтеза (см.: «Великое кислородное событие» на рубеже архея и протерозоя не было ни великим, ни событием, «Элементы», 02.03.2014). Малый временной интервал между изобретением оксигенного фотосинтеза и ростом уровня кислорода в атмосфере подчеркивает ключевую роль данного эволюционного события в оксигенизации атмосферы (в противовес идеям о ведущей роли абиотических процессов, таких как подъем материков и изменение характера вулканизма (см.: Новая модель связала образование земной коры, вулканы и кислород, «Элементы», 17.10.2011).
Вывод о позднем появлении кислородного фотосинтеза согласуется с геохимическими данными, свидетельствующими о существовании 2,415 млрд лет назад фотосинтезирующих организмов, окислявших марганец (J. E. Johnson et al., 2013. Manganese-oxidizing photosynthesis before the rise of cyanobacteria). Марганцевый фотосинтез считается непосредственным эволюционным предшественником оксигенного. Напоминанием об этом этапе эволюции является водоокисляющий марганцевый кластер фотосистемы II (см. главу 16 книги Михаила Никитина «Происхождение жизни. От туманности до клетки»). Возможно, марганцевым фотосинтезом занимались базальные Oxyphotobacteria.
Источник: Rochelle M. Soo, James Hemp, Donovan H. Parks, Woodward W. Fischer, Philip Hugenholtz. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria // Science. 2017. V. 355. P. 1436–1440.
См. также о появлении кислородного фотосинтеза и оксигенизации атмосферы:
1) Новая модель связала образование земной коры, вулканы и кислород, «Элементы», 17.10.2011.
2) «Великое кислородное событие» на рубеже архея и протерозоя не было ни великим, ни событием, «Элементы», 02.03.2014.
3) Михаил Никитин. Происхождение жизни. От туманности до клетки (Глава 16).
-
В товарищах явно согласья нет. :)
Вот пара недавних статей, в которых тоже довольно убедительно утверждается, что оксигеному фотосинтезу минимум 3.2, а скорее, и все 3.5 миллиардов лет:
http://biorxiv.org/content/biorxiv/early/2017/02/17/109447.full.pdf
http://onlinelibrary.wiley.com/doi/10.1111/pala.12178/pdf
Возможно, что весь комплекс генов для оксигенного фотосинтеза с обоими фотосистемами, каротинами, цитохромом B и т.д., был занесён в геном последнего общего предка цианобактерий вирусами от какого-то другого ныне вымершего фотосинтетика. Иначе очень странно, что фактически не сохранилось промежуточных форм, даже у самой примитивной цианобактерии все многие десятки генов для кислородного фотосинтеза уже наличествуют в практически готовом виде.-
Это демонстрирует только точность часов на таких отдаленных гомологиях и таких больших временах.
Схема Кардоны предполагает долгую эволюцию ФС2, а если учесть, что и она сама возникла из ФС1, стало быть, тоже, гомодимерной, и СР47 и СР43 по Кардоне тоже независимо от ФС2 образовались из ФС1, вся эта дивергенция до почти полной неузнаваемости требует очень большого времени. А кислородный фотосинтез, стало быть, возник с самого начала, т.е. потенциально был уже в структуре (гомодимерного же) ФС1. А еще где-то рядом были и чем-то (интересно, чем) занимались два предка аноксигенных фототрофов с хиноновыми и железо-серными РЦ - иначе как бы они сохранились сейчас.
Весь этот нарратив выглядит немного сказочным.
Ваше "занесен в геном вирусами" не спасает. Вирусами можно занести один фотосинтетический комплекс, как у Gemmatimonadetes, или несколько недостающих ферментов цикла Креббса (как у Oscillochloris), или антенну, как у Chloroflexi, но сразу весь фотосинтетический аппарат, это, пожалуй, чересчур.-
Ну, кроме молекулярных часов есть и данные о наличии в среде "кислородных оазисов" уже в породах возрастом 3 млрд. лет и даже ранее.
Я так понимаю, что фотосистема гелиобактерий самая архаичная, она причудливо сочетаят в себе черты реакционных центров более продвинутых фотосинтетиков. Например, электроны с РЦ могут скидываться как на железо-серные кластеры, так, вероятно, и на хиноны, цитохром b состоит из двух протеинов как у цианобактерий, но электроны при этом уходят не на цитохром f, как у оксигенов, а на цитохром с, как у аноксигеных фотосинтетиков и т.д., список можно продолжить. Впоследствии от данного РЦ произошли предки современных РЦ обоих типов и их дальнейшая дивергенция и специализация. Кислородный фотосинтез возник когда уже произошло разделение на ФС1 и ФС2 (причём у цианобактерий были и тот, и другой), но оба протеина ещё были гомодимерами.
Горизонтальный перенос, это основной путь распространения генов фотосинтеза, это видно хотя бы по тому, что на филогенетическом дереве бактерий он проявляется у совершенно не связанных между собой групп бактерий. Как я уже писал выше, все гены из фотосинтетического кластера гелиобактерий - несомненный горизонтальный перенос, тому есть масса независимых подтверждений (если есть желание, можно обсудить это подробнее). То, что весь кластер фотосинтетических генов Gemmatimonadetes (а там не только гены, кодирующие РЦ, но и те, что необходимы для его сборки, гены для синтеза бактериохлорофиллов, и даже гены для синтеза каротинов) перенесён от Rhodobacter считается уже практически доказанным. Кстати, по филогении гены для синтеза бактериохлорофиллов хлороби ближе всего к хлорофлекси, а ацидобактерий - к протеобактериям, но вот незадача, у них РЦ разных типов!
-


Или, наоборот, ставит ее под сомнение? Теория «После более полумиллиарда лет упорного и самоотверженного производства О2 фотосинтетикам, наконец, удалось изменить состав атмосферы планеты» выглядела гораздо правдоподобнее, чем «фотосинтетики перевернули мир в результате удачного 100миллионлетнего блицкрига». Короткий интервал между этими событиями скорее показывает первобытных фотосинтетиков не героями биохимического труда, а оппортунистами, которым повезло втрендиться в меняющиеся условия среды. Например, имея систему для производства кислорода, благоразумно иметь также систему защиты от него. А последняя оказывается очень полезной, если мир вокруг начинает обогащаться кислородом по независящим от фотосинтетика причинам.
Трудно сразу сообразить как же цианобактерии могли несколько раз(!) и независимо сформировать нужные "оксигенные" комплексы, заимствуя их элементы у разных бактерий, у которых комплекса, как раз, и не было. И чем были так особенны Oxyphotobacteria, что удавался этот фокус и не раз только им (из тех, кто дожил до сего дня)?
Может молекулярные часы "сбились"? Как же окисление железа в океанах, джеспелиты Исуа (3,7-3,8 млрд.лет)? А соотношение изотопов С12 и С13 изменяется только при оксигенном фотосинтезе? Или при других вариантах тоже?
Но неродственность "дыхательных белков" все равно впечатляет
-
-
Соотношение С12/С13 изменяется в цикле Кальвина, который входит и в кислородном фотосинтез, и в бескислородный, и в ряд вариантов хемосинтеза.
-
Но получить горизонтальным переносом почти одновременно 2 фотосинтетических комплекса от разных источников, плюс пластоцианин, отрастить себе bf-комплекс, фикобилиновую антенну и водоразлагающую систему? Слишком много совпадающих событий, да и противоречит это большинству гипотез о происхождении фотосинтеза. Пожалуй, экономнее горизонтально приобрести вместо этого единственный рибосомный оперон.
-
С вашего разрешения вставлю пару слов. Вот в этой статье: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4773611/ автор на основе анализа всех описанных на сегодняшний день типов реакционных центров показывает, что оксигенный фотосинтез, по видимому, появился ещё до формирования RC2 хлорофлекси. Кроме того, окислять железо при фотосинтезе, насколько я знаю, на сегодняшний день умеют лишь появившиеся уже после цианобактерий протеобактерии.
Не очень понятно, что объясняет предположение о горизонтальном переносе всего рибосомного оперона. Ведь мелаинобактерии получаются сестринской группой цианобактерий не только на деревьях по 16s RNA и рибосомным белкам, но и по многим другим генам, никакого отношения к рибосоме не имеющим. В общем, похоже, вечер перестаёт быть томным. :)-
Железным фотосинтезом балуются и хлоробии, есть такой Chlorobium ferroxidans, близкий к Chl. clathratiforme и симбионтам консорций (о коих мало что известно). Я бы не рискнул судить, до или после - вот пока считалось, что цианобактерии очень древняя группа, можно было смело говорить, что протеобактерии появились позже. А теперь нет. Часы тут совсем не точны.
Я пока не стал воровать новую статью, но в предыдущих деревья строили только по 16S и рибосомным белкам. Отсюда рибосомный оперон.
Да, ситуация с эволюцией фотосинтеза очень сильно запутывается.-
По поводу Chlorobium ferroxidans - спасибо, не знал, новая для меня информация. Общие предки и альфа и бета протеобактериий, судя по всему, уже были аэробами. Из этого факта, а так же с учётом того, что по 16s RNA и многим другим деревьям общий предок мелаинабактений и цианобактерий ответвился от клостридий, на мой взгляд, можно достаточно уверенно утверждать, что альфа, бета и гамма протеобактерии появились уже после GOE.
-
-
-
Что же до Кардоны, то я могу согласиться, что фс2 впервые появилась у процианобактерий, но, вероятно, когда они ещё не были кислородными. Железо они там окисляли или водород - вопрос десятый. Но позднее появление "оксифотобактерий" этот сценарий практически убивает. Они должны были нахватать уже готовое. Не верю.
А почему не могло быть двух параллельных редукций фс1 - первая могла дать фс2 цианобактерий плюс их ближнюю антенну, а вторая, независимая - хиноновые фотосистемы хлорофлексий и протеобактерий?-
А откуда, собственно, следует "позднее" появление окситрофов? По моим прикидкам, основанным на молекулярных данных, получается, что общий предок троицы "Oxyphotobacteria, Melainabacteria, Sericytochromatia" отделился от клостридий где-то примерно в районе 3.7 млрд. лет назад. Последующая их дивесификация произошла довольно быстро. Кстати, верные признаки оксигенного фотосинтеза - пузырьки с кислородом, образующиеся внутри цианобактериальных матов, наблюдаются уже начиная с рубежа в 2.7 млрд. лет, и никто из сторонников "позднего пришествия" оксигенов, насколько я понимаю, данный аргумент пока не опроверг.
Если верить Кардоне, то по его реконструкциям выходит, что предковый для всех RC второго типа комплекс (он был образован гомодимером) уже имел некотрые предадаптации для разложения воды, в частности, лигады для Mn4CaO5 кластера.
Собственно, в том, что первая примитивная фотосистема больше напоминала нынешнюю ФС1, по моему, сейчас мало кто сомневается. Причём, ФС гелиобактерий это, похоже, что-то типа окаменелости первой примитивной фотосистемы цианобактерий, сохранившейся в геноме гелиобактерий до наших дней. Очень интересно, что один из участков протеина, представляющего её реакционно-антенный комплекс, имеет довольно хорошую гомологию с фрагментом антенного комплекса RC2 цианобактерий CP47!-
Combinator, myugor1960,
> Кроме того, окислять железо при фотосинтезе, насколько я знаю, на сегодняшний день умеют лишь появившиеся уже после цианобактерий протеобактерии.
> Железо они там окисляли или водород - вопрос десятый.
Под окислением железа в океанах понимается совсем другой процесс: абиогенное окисление свободного (растворенного) двухвалентного железа до нерастворимого трехвалентного свободным кислородом. Преобладающее мнение сейчас, что пока почти всё двухвалентное железо в океане не окислилось, накопления кислорода в результате оксигенного фотосинтеза в атмосфере в занчительных масштабах не происходило, ибо кислород поглощался океаном. Когда процесс окисления железа в океане в основном закончился, тогда и произошла кислородная катастрофа в атмосфере.-
Насколько я знаю, в настоящее время есть как сторонники биогенного окисления железа в океанах арехея аноксигенными фототрофами (тот же М.Никитин, например), так и те, кто придерживается теории его окисления кислородом, выделяемым уже тогда существовавшими цианобактериями - оксигенами. Не уверен, что можно говорить о каком-то "пробладающем" мнении, хотя мне лично тоже ближе вторая точка зрения.
-
Да бог с ним, с преобладающим мнением, посмотрите просто на залежи железных руд, взвесьте их. Если весь этот Fe3+ биологического происхождения, то есть кто-то в архее просто _питался_ этим железом-2, пропуская его через свои животики и перерабатывая в железо-3, то тогда окей, соглашаемся с первой точкой зрения.
-
-
-
Ну что Вы, под окислением железа (в океанах и где угодно) понимается любое окисление, независимо от механизма, и биогенное, и абиогенное.
-
Вы так и не поняли, даже неловко объяснять. В двух постах выше вы противоречите сами себе. Сначала пишите, что окисление биогенное, если кислород биогенного происхождения, а теперь пишите, что любое окисление это просто окисление (правильно конечно второе, но важна терминология и что именно за ней стоит).
Разрежьте яблоко на свежем воздухе, оно покроется ржавчиной - это Fe2O3. Как по-вашему - это какое окисление? Правильно - это обычная химическая реакция из области неорганической химии, окисление двухвалентного железа до трехвалентного в присутствии кислорода. Никакие при этом биологические каскады, структуры, белковые комплексы, метаболизм в яблоке в этой реакции не работают в том смысле, что ничего они такого не делают для протекания этой реакции. Это мертвая реакция, пассивный процесс (со стороны яблока). Можно сказать это абиогенное окисление, ничем не отличающееся от реакции в какой-нибудь пробирке. А можно типа сказать, что окисление биогенное, т.к. исходное сырье (ионы двухвалентного железа, атмосферный кислород) биологического происхождения, в том смысле, что их бы там не было в таком виде, если бы не жизнь на Земле, но это утверждение хоть и верное, но дополнительных знаний нам не приносит, потому бессмысленное.
А теперь возвращаемся к вашему первому утверждению: "Железо они там окисляли или водород - вопрос десятый". Всякому непредвзятому будет очевидно, что вы здесь перепутали пассивную, мертвую, химическую реакцию окисления железа в океане - и именно это везде понимается под окислением железа в океане, а не что-то иное, т.е. соединение железа, находящегося в воде океана, и кислорода, находящегося в атмосфере, а вовсе не соединение железа и кислорода, находящихся внутри живых клеток - с активным процессом сродни питанию, азотфиксации и проч. из того же ряда, которые протекают внутри живых клеток _как часть их метаболизма_. Так вот в этом вашем смысле никакого биогенного, метаболического окисления железа в океане, по крайней мере сравнимого в объёме с первым, не существует, разве что в воображении Никитина. Никто железо в океане не окислял - само оно окислилось, понимаете? Ваша фраза "Железо они там окисляли или водород - вопрос десятый" - абсурдна и свидетельствует о каше в вашей голове.
Так понятно разжевал?-
>но это утверждение хоть и верное, но дополнительных знаний нам не приносит, потому бессмысленное.
Я так не считаю, наоборот, imho это вопрос очень существенный, поскольку без фотосинтеза не было бы и архейского рудообразования (уф-фотолизный кислород я в расчет не принимаю). Т.е. чтобы образовывались руды, должна была фиксироваться (восстанавливаться)углекислота. Без этого никак, и неважно, был ли при этом интермедиат в виде кислорода или не было. В любом случае, железо окислял именно "кто-то", а были это оксигенные или аноксигенные фототрофы - неважно.
> вы здесь перепутали пассивную, мертвую, химическую реакцию окисления железа в океане
Нет, не перепутал, я говорил о фотосинтезе, а не о "мертвой реакции".
Железо в водоемах окисляется не только абиогенно, но и железобактериями и железоокисляющими АФБ. В аэробных условиях, разумеется, абиогенный процесс преобладает, но в микроаэробных, уже при концентрации кислорода навскидку менее 5% поверхностной, биота успешно конкурирует с абиотическим окислением. Ну, а в анаэробных условиях, да еще в отсутствие сульфатов и нитратов/нитритов "железный" фотосинтез - вообще единственный способ окисления железа. Поверьте, современные океанические условия - не лучшая модель процессов, которые шли тогда. Если вы, конечно, не считаете, что в архейском океане были современные 8-10 мг/л кислорода. -
>Разрежьте яблоко на свежем воздухе, оно покроется ржавчиной - это Fe2O3
Это ж сколько в яблоке железа, что оно так эффектно окисляется! И какое интересное, кстати - побланшировали яблоко в кипятке 4-5 минут, и "двухвалентное железо" в нем больше вовсе не окисляется, сколько его на воздухе не держи.
А вообще, погуглите про полифенолы, про то, как полифенолоксидаза окисляет их до хинонов, и как хиноны реагируют с аминокислотами, образуя окрашенные в коричневый цвет продукты реакции.
-
-
-
-
Если же такого четкого подтверждения нет, то вместо термина "молекулярные часы" следует употреблять термин "дурацкая экстрополяция"
Ведь имеется очень надежная информация о том, что условия на Земле например в архее очень сильно отличались от современных. Два примера :
а) Земля вращалась намного быстрее, а Луна была в 10 раз ближе так, что если земной океан по площади был сопоставим с современным то приливные волны имели высоту в сотни метров и несколько раз в сутки (современные) обрушивались на сушу (такая волна способна почти полностью затопить бассейн современной Амазонки);
б) в условиях отсутствия кислорода в водоеме с остатками какой-нибудь органики может начаться процесс брожения (завелись протодрожжи) и мы получим озеро бражки и мутагенез в заспиртованном состоянии.
Р.S. Особенно прикольно выглядят рассуждения о молекулярных часах на основании метагеномного анализа, самих бактерий никто ещё в глаза не видел, об их метаболизме имеются самые общие представления, а вот ход часов известен.
-
Я не Марков, но отвечу. Скорость накопления замен определяется в основном отбором, а не мутациями. Мутаций всегда более чем достаточно, но стабилизирующий отбор пропускает хорошо если 1% самых безвредных (а то и 0,0001%). Молекулярные часы работают, если ген у всех исследуемых организмов имеет одну и ту же функцию и не подвергается движущему отбору. Эти условия плюс-минус верны для рибосомной РНК. А для дыхательных и фотосинтетических белков бактерий скорее неверны, поэтому для молекулярных часов их брать нельзя.
Последние новости







Рис. 1. Разнообразие фотосинтезирующих цианобактерий (класс Oxyphotobacteria). Показаны представители отрядов Chroococcales (a–e), Pleurocapsales (f), Oscillatoriales (g–p), Nostocales (q–t), Stigonematales (u–v). Длина масштабных отрезков: 10 мкм (a–u), 20 мкм (v). Изображение из статьи P. Dvorak et al., 2015. Species concepts and speciation factors in cyanobacteria, with connection to the problems of diversity and classification