Полиплоидность предков эукариот — ключ к пониманию происхождения митоза и мейоза

Предложена гипотеза, согласно которой эукариоты произошли от полиплоидных архей, обитавших в мелководных микробных сообществах раннего протерозоя, когда уровень свободного кислорода начал расти, но озонового экрана еще не было, что должно было резко повысить темп мутагенеза. Моделирование показывает, что в таких условиях полиплоидность у прокариот, не имеющих митоза, дает кратковременное преимущество, но в долгосрочной перспективе повышает риск вымирания из-за накопления рецессивных вредных мутаций. Полиплоидные микробы могли справляться с этой проблемой несколькими способами, постепенное совершенствование и комбинирование которых логически приводит к появлению сначала митоза, а затем мейоза и полового размножения. Новая гипотеза объясняет несколько ключевых особенностей эукариот, до сих пор остававшихся загадочными, в том числе множественные линейные хромосомы, высокий уровень генетической избыточности и быстрое появление новых генных семейств на заре эволюции эукариот, выявленное сравнительной геномикой. К числу фактов, согласующихся с гипотезой, относится недавно обнаруженная корреляция между полиплоидностью и наличием гистонов у архей.
1. Происхождение эукариот и половое размножение
Появление эукариот — второе по значимости событие в истории земной жизни (первое — само появление жизни). Эукариотическая клетка устроена гораздо сложнее, чем прокариотическая, а промежуточные формы между ними, существовавшие когда-то, давно вымерли. Поэтому вопрос о происхождении эукариот остается одной из самых сложных и спорных тем в эволюционной биологии (см.: А. В. Марков, А. М. Куликов, 2009. Происхождение эукариот как результат интеграционных процессов в микробном сообществе). Правда, недавно ситуация резко изменилась к лучшему в результате открытия локиархей — неизвестной ранее группы архей, обладающей многими признаками, которые до сих пор считались уникальными для эукариот (см.: Новооткрытый микроб заполняет брешь между прокариотами и эукариотами, «Элементы», 12.05.2015).
Но даже с учетом локиархей получается, что эволюционный путь от последнего общего предка с ближайшими архейными родственниками до последнего общего предка всех современных эукариот (LECA, last eukaryotic common ancestor) был долгим и трудным. На этом пути предки эукариот приобрели целый ряд признаков, которых нет ни у кого из прокариот, даже у локиархей. Одним из последних шагов на этом пути, по-видимому, стало приобретение внутриклеточных симбионтов — альфапротеобактерий, давших начало митохондриям (см.: Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016).
К числу эукариотических инноваций, наиболее трудных для объяснения, относится эукариотический секс (называемый также амфимиксисом или, попросту, половым размножением). Это специфический и очень эффективный способ перемешивания генетического материала разных особей, включающий сингамию (слияние гаплоидных клеток или ядер, приводящее к удвоению хромосомного набора) и мейоз (особый вариант клеточного деления, приводящий к сокращению числа хромосом вдвое). Мейоз сопровождается кроссинговером, в ходе которого гомологичные хромосомы обмениваются гомологичными участками.
Прокариотический секс устроен гораздо проще: это однонаправленная передача небольших фрагментов генетического материала от микроба-донора микробу-реципиенту (см. врезку и рис. 2).
Отличия эукариотического секса («полового размножения») от прокариотического (горизонтального переноса генов, ГПГ)
- У эукариот обмен генами взаимный, двусторонний: каждый участник является и донором, и реципиентом.
У прокариот один участник — донор ДНК, другой — реципиент.- Полногеномная рекомбинация: в обмене участвуют два полных генома.
У прокариот в рекомбинации участвует один полный геном и один фрагмент.- Все участвующие гены сохраняют шанс перейти к потомкам.
У прокариот фрагмент ДНК донора либо встраивается в геном реципиента вдобавок к уже имеющимся там генам (но этот вариант не может использоваться регулярно, например, в каждом поколении), либо замещает собой гомологичный фрагмент генома реципиента. При этом замещенные аллели реципиента уничтожаются и теряют шанс перейти к потомкам (рис. 2).
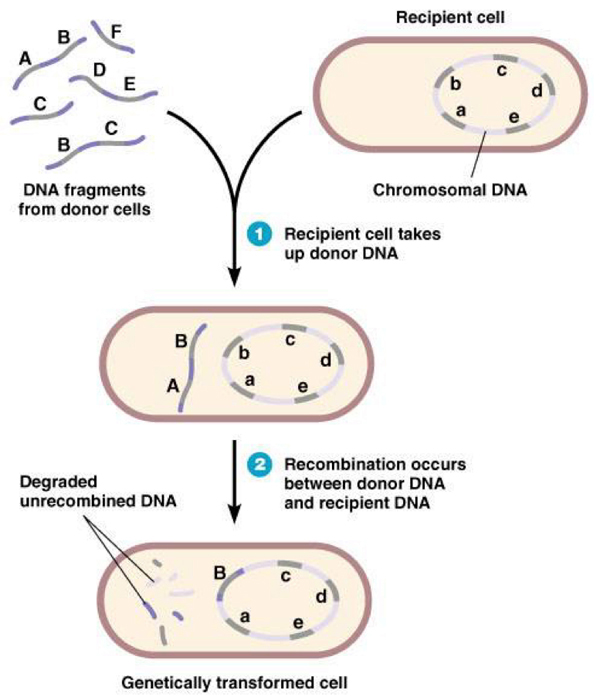
Рис. 2. Один из способов горизонтального переноса генов (ГПГ) у прокариот (естественная трансформация, сопряженная с гомологичной рекомбинацией), наиболее близкий по своим эволюционным последствиям к эукариотическому сексу. Бактерия поглощает фрагмент чужой ДНК из окружающей среды (это могут быть, например, фрагменты геномов погибших бактерий того же вида), а затем встраивает его в свою кольцевую хромосому вместо собственного гомологичного фрагмента. В результате аллели бактерии-реципиента уничтожаются, замещаясь чужими аллелями. В данном случае аллель B встроился в хромосому и будет передан потомкам, а аллель b погиб. Рисунок с сайта bio.miami.edu
Раньше многие специалисты предполагали, что первые эукариоты были бесполыми (размножались клонально — простым делением и не имели полового процесса), а потом у кого-то из них появилось половое размножение. В таком случае проблема появления эукариотического секса не связана напрямую с проблемой происхождения эукариот. Однако по мере накопления геномных данных стало ясно, что эукариотический секс является одной из фундаментальных особенностей эукариот, которая почти наверняка имелась уже у LECA. В частности, появляется всё больше аргументов в пользу того, что все современные бесполые группы эукариот произошли от предков, имевших мейоз, а значит и половой процесс. Стало быть, происхождение эукариот и происхождение полового размножения — проблемы взаимосвязанные.
2. Половой процесс солелюбивой археи Haloferax — нечто промежуточное между типичным прокариотическим и эукариотическим сексом
Если эукариотический секс — исходная черта эукариот, то имеет смысл поискать среди современных архей варианты генетического обмена, переходные между типичным прокариотическим и эукариотическим сексом. Крайне интересно было бы выяснить, как меняются генами локиархеи, но об этом пока ничего не известно. На сегодняшний день подробно изучена система генетического обмена только у одной археи — солелюбивого (галофильного) микроба Haloferax volcanii, обитателя Мертвого моря. Об этом микробе рассказано в новости Альтернативный способ репликации ДНК оказался эффективнее традиционного («Элементы», 12.11.2013). У Haloferax обнаружена система передачи генетического материала, действительно похожая на что-то промежуточное между обычным прокариотическим ГПГ (например, конъюгацией бактерий) и эукариотическим сексом (рис. 3).

Рис. 3. Спаривание архей Haloferax volcanii. Слева — фотографии, сделанные при помощи сканирующего электронного микроскопа. Видны перемычки между клетками — цитоплазматические мостики, по которым передается ДНК. Масштабные отрезки — 1 мкм. Справа — схема временного слияния клеток, которое происходит в лаборатории (а может быть — и в природе) в результате дестабилизации цитоплазматических мостиков. Изображения из статей I. Rosenshine, R. Tchelet, M. Mevarech, 1989. The Mechanism of DNA Transfer in the Mating System of an Archaebacterium и R. T. Ortenberg, R. M. Mevarech, 1999. A model for the genetic Exchange system of the extremely halophilic archaeon Haloferax volcanii
Клетки Haloferax спариваются, соединяясь цитоплазматическими мостиками, по которым геномная ДНК может передаваться в обе стороны, то есть обе клетки могут быть как донорами, так и реципиентами генетического материала. При этом могут передаваться крупные фрагменты хромосомы (длиной до 13% генома как минимум). В лаборатории можно добиться даже слияния клеток: для этого нужно понизить концентрацию Mg2+ в среде, что приводит к растворению клеточных стенок и дестабилизации цитоплазматических мостиков (рис. 3, справа). Предполагается, что такое слияние иногда может происходить и в природе.
В 2010 году была предложена красивая гипотеза, связывающая воедино происхождение эукариот и полового размножения и опирающаяся в том числе на данные по половому процессу Haloferax (см.: J. Gross, D. Bhattacharya, 2010. Uniting sex and eukaryote origins in an emerging oxygenic world, статья в открытом доступе). В статье обосновываются три положения:
1) Первые эукариоты появились в раннепротерозойских мелководных местообитаниях, когда концентрация кислорода начала расти, но озонового экрана еще не было. Под действием ультрафиолета на мелководьях должна была повыситься концентрация активных форм кислорода — сильных мутагенов. Появление эукариот стало закономерным итогом попыток архей защититься от этой напасти.
2) Архейные предки эукариот обменивались генами примерно так же, как современный Haloferax. Авторы выстроили гипотетический сценарий постепенного эволюционного перехода от полового процесса Haloferax к полноценному эукариотическому сексу.
3) Совершенствование системы генетического обмена было необходимо предкам эукариот, потому что они использовали чужую ДНК в качестве матрицы для починки разрывов двойной спирали ДНК при помощи гомологичной рекомбинации (примерно так, как это делает радиоустойчивый микроб Deinococcus raduodurans, см.: Разгадана тайна микроба, не боящегося радиации, «Элементы», 03.10.2006).
При всех достоинствах этой гипотезы, она не учитывает одну важную деталь, которая не противоречит первым двум пунктам, но сильно бьет по третьему. Дело в том, что Haloferax, как и многие другие археи, является полиплоидом. В каждой клетке Haloferax содержится в среднем по 17 копий генома (кольцевых хромосом).
3. Полиплоидность архей и ее эволюционные следствия
Все привыкли думать, что у прокариот в клетке только одна кольцевая хромосома. У многих бактерий и архей это действительно так. «По умолчанию» считалось, что Haloferax тоже является моноплоидом: это отражено и на рис. 3, который взят из статьи 1999 года, и на рисунках в статье Гросса и Бхаттачарьи 2010 года. Однако исследования последних лет показали, что полиплоидность (наличие многочисленных копий генома в одной клетке) широко распространена у бактерий и архей. В частности, полиплоидами являются, по-видимому, все галоархеи (к которым относится Haloferax), метаногены и термоплазмы. Полиплоидной клетке не нужна чужая ДНК, чтобы чинить разрывы при помощи гомологичной рекомбинации: у нее для этого достаточно собственных хромосом.
Полиплоидность может быть полезна прокариотам по целому ряду причин, включая упомянутую репарацию разрывов ДНК, отсроченное фенотипическое проявление вредных рецессивных мутаций и повышенную надежность синтеза белка в неблагоприятных условиях. Помимо прочего, «лишняя» геномная ДНК служит запасом пищи на черный день (полиплоидные археи съедают часть своих хромосом, когда им не хватает фосфора).
Полиплоидность Haloferax заставляет по-новому взглянуть на природу факторов отбора, которые привели к появлению эукариотического секса. При этом всё становится даже интереснее, чем в исходной модели Гросса и Бхаттачарьи.
В журнале Biology Direct (где ранее уже было опубликовано несколько важных статей по происхождению эукариот, в том числе статья Гросса и Бхаттачарьи) вышла новая статья, написанная мной в соавторстве с программистом Ильей Казначеевым, в которой мы при помощи компьютерного моделирования попытались выяснить, что будет происходить с популяцией полиплоидных архей, если она попадет в сильно мутагенную среду. Анализ эволюционно-генетических последствий полиплоидности у прокариот, обитающих в мутагенной среде, шаг за шагом логически привел нас к появлению полового размножения.
Для начала нужно пояснить, что у полиплоидных архей нет митоза. При делении родительской клетки хромосомы распределяются между потомками примерно поровну, поэтому уровень плоидности остается более или менее постоянным, но то, какие именно хромосомы достанутся каждому потомку, по-видимому, никак не регулируется и определяется случайностью (рис. 1). Из этого вытекают интересные эволюционно-генетические следствия. В частности, копии гена, расположенные на разных хромосомах, не могут поделить между собой функции, как это часто происходит у имеющих митоз эукариот после полиплоидизации (см.: Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008). Ведь если нет митоза, то нет и никакой гарантии, что каждый потомок получит оба специализированных варианта гена.
Мы разработали компьютерную модель, имитирующую эволюцию популяции одноклеточных организмов с различной плоидностью. У каждой клетки есть определенное число исходно одинаковых кольцевых хромосом, каждая хромосома содержит определенное число локусов (генов). Каждый ген характеризуется «качеством работы», которое может ухудшаться из-за вредных мутаций и улучшаться из-за полезных. От качества генов зависит конкурентоспособность клетки и ее шансы на выживание в каждом поколении. В модели можно регулировать скорость мутагенеза, соотношение вредных и полезных мутаций, численность популяции, параметры отбора и дрейфа, способ распределения хромосом по дочерним клеткам при делении и многое другое. Кроме того, модель позволяет имитировать разные варианты генетического обмена и гомологичной рекомбинации как внутри клетки, так и между клетками. Для простоты мы приняли, что все вредные мутации рецессивны, и поэтому для вычисления приспособленности полиплоидной клетки использовалась «лучшая» из копий каждого гена. Это справедливо для многих мутаций, снижающих функциональность кодируемого белка. Таким образом, проведенный анализ относится только к тем генам, одной исправной копии которых достаточно для выживания клетки.
Моделирование показало, что при высокой скорости мутагенеза моноплоиды в долгосрочной перспективе более жизнеспособны, чем полиплоиды. Однако поначалу (первые несколько сотен поколений) полиплоиды опережают моноплоидов по приспособленности (рис. 4).
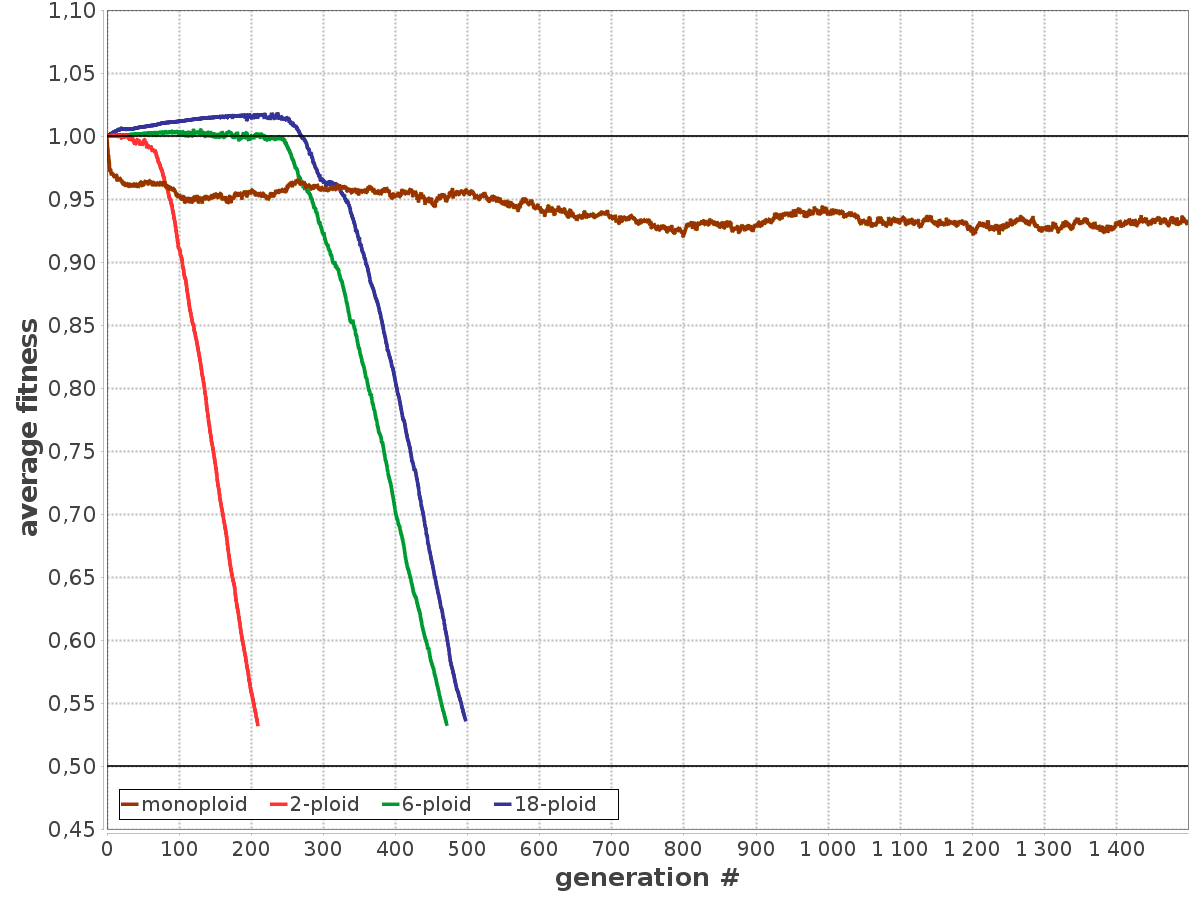
Рис. 4. Пример работы модели, показывающий эволюционную судьбу популяций с различной плоидностью при высокой скорости мутагенеза. Митоза нет, генетического обмена и рекомбинации тоже нет. По горизонтальной оси — поколения, по вертикальной — средняя приспособленность клеток в популяции. Моноплоиды выходят на равновесный уровень приспособленности (см. Mutation-selection balance) и существуют неопределенно долго. Полиплоиды сначала имеют сильное преимущество, но затем быстро деградируют и вымирают. Рисунок из обсуждаемой статьи в Biology Direct
Причин кратковременного преимущества полиплоидов две. Первая — отложенное проявление в фенотипе вредных рецессивных мутаций. Такая мутация у полиплоидов не дает о себе знать, пока не начнут появляться клетки, несущие ее в каждой копии генома (гомозиготные). Вторая причина — ускоренное накопление редких доминантных полезных мутаций. При фиксированной частоте их возникновения в расчете на локус, вероятность приобретения клеткой полезной мутации прямо пропорциональна плоидности.
Причины последующего вырождения — слабый очищающий отбор, быстрое накопление рецессивных вредных мутаций и рост сегрегационного груза. Это значит, что в какой-то момент жизнеспособные полиплоидные клетки начинают производить нежизнеспособных потомков. Это неизбежное следствие накопления рецессивных вредных мутаций при отсутствии митоза. Термин «сегрегационный груз» обычно применяют к эукариотам в ситуации, когда гетерозиготы имеют более высокую приспособленность, чем гомозиготы (сегрегация — разделение, в данном случае имеется в виду распределение родительских хромосом по геномам потомков). При скрещивании двух гетерозигот часть потомков оказываются гомозиготами и имеют пониженную приспособленность — это и называют сегрегационным грузом. В приложении к полиплоидным археям имеется в виду следующее. Представим себе триплоидную клетку, у которой из трех жизненно необходимых генов A, B и C первый исправен только на первой хромосоме, второй ген исправен на первой и третьей хромосомах, третий — только на второй. Такая клетка, сама по себе вполне жизнеспособная, может поделиться на двух нежизнеспособных потомков, если ей не повезет:
ABc репликация ABc ABc деление ABc abC
abC → abC abC → ABc + abC
aBc aBc aBc aBc aBc
Как видим, у первого потомка нет ни одной работающей копии гена C, а второму не досталось рабочих копий гена A. Заметим, что при наличии митоза этой проблемы не было бы: оба потомка имели бы точно такой же генотип, как у родительской клетки, и жили бы припеваючи.
Поскольку полиплоидность приводит к вырождению не сразу, а сначала дает сильное преимущество, она может стать своеобразной «эволюционной ловушкой» для прокариот в мутагенной среде. Если разрешить модельным микробам иногда при делении распределять хромосомы не поровну, то есть менять свою плоидность, то полиплоиды сначала быстро вытесняют моноплоидов, а затем сами деградируют и вымирают. Это происходит даже в том случае, если подавляющее большинство клеток в исходной популяции — облигатные моноплоиды, и лишь немногие клетки способны иногда производить потомков, имеющих на одну хромосому больше. Полиплоидность распространяется, как инфекция, и приводит популяцию к гибели. При тех же параметрах популяция, состоящая только из облигатных моноплоидов, может жить неопределенно долго.
4. Четыре способа защиты полиплоидов от вырождения
Итак, быть не имеющим митоза полиплоидом — дело рискованное, особенно в мутагенной среде. Чтобы выжить, такие полиплоиды должны выработать специальные адаптации, замедляющие накопление вредных мутаций. Они могут использовать для этого как минимум четыре разные стратегии. Вот тут-то и начинается самое интересное. Дело в том, что все эти четыре способа защиты полиплоидных прокариот от вырождения подозрительно напоминают те или иные аспекты эукариотического секса. Подобные адаптации могут развиться в ходе эволюции, даже если не приносят немедленной пользы клетке, за счет «отбора второго порядка на эволюционную перспективность», реальность которого была продемонстрирована в эксперименте Ленски (см.: В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011).
Первый способ — «циклы плоидности». Можно периодически сбрасывать уровень плоидности, например, делясь чаще, чем происходит репликация хромосом. Если заниматься этим достаточно интенсивно, то в популяции будет постоянно присутствовать (или периодически возникать) заметная доля моноплоидов, у которых все вредные рецессивные мутации проявляются в фенотипе и поэтому эффективно вычищаются отбором.
Второй способ — внутригеномная рекомбинация, то есть перетасовка генетической информации между хромосомами. Есть два основных варианта такой рекомбинации. Первый вариант называется «генная конверсия». В этом случае фрагмент одной хромосомы копируется в гомологичный участок другой, причем аллели, находящиеся на второй хромосоме, «затираются» аллелями первой. Интенсивная генная конверсия ведет к унификации копий генома. Любая новая мутация либо быстро затирается и исчезает, либо распространяется на все хромосомы, переходит в гомозиготное состояние и становится видимой для отбора. Моделирование показывает, что генная конверсия, если ее интенсивность существенно превышает темп мутагенеза, может спасти полиплоидную популяцию от вырождения.
Полиплоидные галофильные и метаногенные археи активно используют генную конверсию, предположительно, именно для того, чтобы унифицировать свои хромосомы и тем самым защититься от вырождения. Эту стратегию, вероятно, с той же целью применяют и пластиды растений (они тоже полиплоидные и не имеют митоза). Второй способ внутригеномной рекомбинации — кроссинговер. В этом случае гомологичные участки двух хромосом не затирают друг друга, а меняются местами. Кроссинговер не может ни уничтожить вредную мутацию, ни перевести ее в гомозиготное состояние. Поэтому сам по себе он бесполезен для полиплоидных микробов, но в сочетании с ГПГ он дает сильный положительный эффект (см. ниже).
Третий способ — интенсивный генетический обмен (ГПГ) между близкородственными клетками. Моделирование показывает, что горизонтальный перенос генов (такой, как на рис. 2) хорошо защищает полиплоидов от вырождения, особенно если он осуществляется с высокой частотой. Полиплоидные археи действительно меняются друг с другом генами изо всех сил. Эволюционный эффект интенсивного близкородственного ГПГ в целом такой же, как у эукариотического секса (см.: В эволюции бактерий горизонтальный генетический обмен играет ту же роль, что и половое размножение у высших организмов, «Элементы», 09.04.2012). Интересно, что кроссинговер, бесполезный для популяций, не практикующих ГПГ, оказывается очень полезен микробам, периодически заимствующим гены друг у друга. Это связано с тем, что ГПГ в сочетании с кроссинговером позволяет эффективно конструировать хорошие хромосомы из плохих, тем самым защищая популяцию от «храповика Мёллера» (см.: Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009).
Польза, приносимая ГПГ, имеет ту же природу, что и польза, приносимая половым размножением: оба процесса позволяют отбору отделять полезные аллели от вредных, эффективно закрепляя первые и выбраковывая вторые (см.: Половое размножение помогает отбору отделять полезные мутации от вредных, «Элементы», 01.03.2016). При этом ГПГ тем полезнее, чем чаще он происходит. При высоком темпе мутирования это особенно актуально. Однако у прокариотического ГПГ есть «встроенный конструктивный дефект», не позволяющий этому процессу достигать оптимальной (то есть высокой) частоты. Дефект кроется в асимметричности («нечестности») прокариотического ГПГ, которую хорошо иллюстрирует рис. 2. На рисунке видно, что чужой (донорский) аллель B заместил и уничтожил аллель b в геноме реципиента. Ситуация, когда свои аллели систематически замещаются чужими, может оказаться эволюционно нестабильной. Чтобы понять это, нужно подумать о судьбе генов, влияющих на интенсивность (частоту) захвата чужой ДНК и замещения собственных аллелей чужими. Допустим, у такого гена есть два аллеля: один способствует ГПГ, другой — препятствует. Какой из них победит в конкуренции? Моделирование показывает, что аллели, препятствующие ГПГ, могут распространяться в генофонде и вытеснять аллели, способствующие ГПГ, несмотря на всю пользу, которую получают от ГПГ отдельные организмы и популяция в целом. Ведь аллели, способствующие ГПГ, будут то и дело «затираться» конкурирующими алелями, которые блокируют ГПГ. А вот в обратную сторону замещение происходить не будет — аллели, блокирующие захват чужой ДНК и замещение ими фрагментов своей хромосомы, не будут затираться как раз потому, что они блокируют ГПГ. В результате аллели, снижающие частоту ГПГ, будут вести себя как эгоистичные гены, наращивая свою частоту в генофонде несмотря на то, что это вредно для особей и популяции в целом.
Могут ли полиплоидные микробы обойти это препятствие, чтобы получить возможность осуществлять межорганизменный генетический обмен с высокой частотой? По-видимому, да. Для этого им нужно, во-первых, начать обмениваться не кусочками хромосомы, а целыми хромосомами, во-вторых — отказаться от асимметричной генной конверсии, исключить «затирание» одних аллелей другими и использовать для перемешивания фрагментов хромосом только кроссинговер. Умеют ли полиплоидные археи меняться целыми хромосомами, точно не известно, но это представляется вполне вероятным исходя из того, что известно о половом процессе у Haloferax.
С кроссинговером, правда, возникает еще одна проблема: кольцевые хромосомы плохо подходят для кроссинговера. При нечетном числе перекрестов они не могут нормально разойтись после рекомбинации и превращаются в одно большое кольцо. Поэтому если вы хотите часто использовать кроссинговер, вам нужно отказаться от кольцевых хромосом и заменить их линейными. Идея о том, что линейные хромосомы понадобились эукариотам именно для частого кроссинговера, а не для чего-то еще, высказывалась ранее рядом специалистов, и с ней трудно спорить, учитывая, что во всех прочих отношениях кольцевые хромосомы удобнее. Что касается многочисленных точек начала репликации, характерных для эукариотических хромосом (у прокариот обычно только одна точка начала репликации), то они могли размножиться еще у архейных предков, потому что изначально были «эгоистичными генами» (на эту крамольную идею исследователей навело изучение того же Haloferax volcanii, см.: Альтернативный способ репликации ДНК оказался эффективнее традиционного, «Элементы», 12.11.2013).
Четвертый способ, помогающий полиплоидным амитотическим микробам защититься от вырождения, — самый радикальный. Он состоит в том, чтобы изобрести митоз — механизм аккуратного и точного распределения хромосом по дочерним клеткам, гарантирующий, что каждый потомок получит ровно по одной копии каждой родительской хромосомы. Это моментально снимает все проблемы, связанные с накоплением сегрегационного груза.
После изобретения митоза все выгоды частого обмена хромосомами и кроссинговера сохраняются в полной мере. Поэтому у полиплоидных архей, научившихся аккуратно распределять хромосомы по дочерним клеткам, не было оснований отказываться от хромосомного обмена. Но со временем это закономерно привело к новому конфликту, для разрешения которого пришлось изобрести мейоз.
5. Изобретение митоза приводит к диверсификации хромосом, что порождает новые проблемы, для решения которых нужно изобрести мейоз
Неизбежным следствием изобретения митоза полиплоидными археями — предками эукариот должна была стать быстрая диверсификация хромосом. Митоз снимает проблему сегрегационного груза, и поэтому избыточные копии генов, расположенные на разных хромосомах, получают небывалую эволюционную свободу. В нашей модели они просто начинают деградировать, свободно накапливая вредные мутации, так что скоро у каждого гена остается только одна неиспорченная копия, расположенная на любой из хромосом. Каждая хромосома при этом становится уникальной и незаменимой, потому что те гены, которые остались неиспорченными на ней, безнадежно испорчены на всех остальных хромосомах (рис. 5).

Рис. 5. Геном произвольно выбранной клетки из популяции 6-плоидных микробов, которые изобрели митоз, после 1000 поколений эволюции. Колонки соответствуют генам. Числа обозначают «качество генов». Зеленым цветом выделены лучшие аллели в каждом локусе. Видно, что каждый ген сохранился в более или менее неиспорченном состоянии только на одной хромосоме, а каждая хромосома имеет свой уникальный набор неиспорченных генов. Рисунок из обсуждаемой статьи в Biology Direct
Наша модель не предусматривает приобретение генами новых функций (неофункционализацию) или специализацию на одном из аспектов исходной функции (субфункционализацию). Однако это именно то, что наверняка будет происходить в подобной ситуации у реальных живых организмов. Многие избыточные гены будут потеряны или безнадежно испорчены прежде, чем в них возникнет полезная мутация, но многие поделят функции или выработают новые.
Таким образом, изобретение митоза фактически превращает полиплоидный организм в моноплоидный, обладающий несколькими разными хромосомами, с высоким уровнем генетической избыточности. Здесь уместно вспомнить, что множественные хромосомы и генетическая избыточность — характерные черты эукариот, происхождение которых долго оставалось загадочным. Наша гипотеза дает им внятное историческое объяснение.
По мере того, как хромосомы специализируются и становятся уникальными, унаследованные от предков способы свободного хромосомного обмена и рекомбинации становятся все менее выгодными, а затем и вовсе начинают приносить вред. Действительно, если все ваши хромосомы уникальны и незаменимы, вы уже не можете просто передать в другую клетку по цитоплазматическому мостику одну-две случайно выбранные хромосомы и получить какие-то хромосомы взамен. Необходимо сделать обмен хромосомами и рекомбинацию более избирательными, чтобы в них участвовали только очень похожие (гомологичные) хромосомы. Моделирование подтверждает эти рассуждения (рис. 6). Кроме того, хорошо бы еще позаботиться о том, чтобы каждая хромосома принимала участие в рекомбинации с оптимальной частотой.
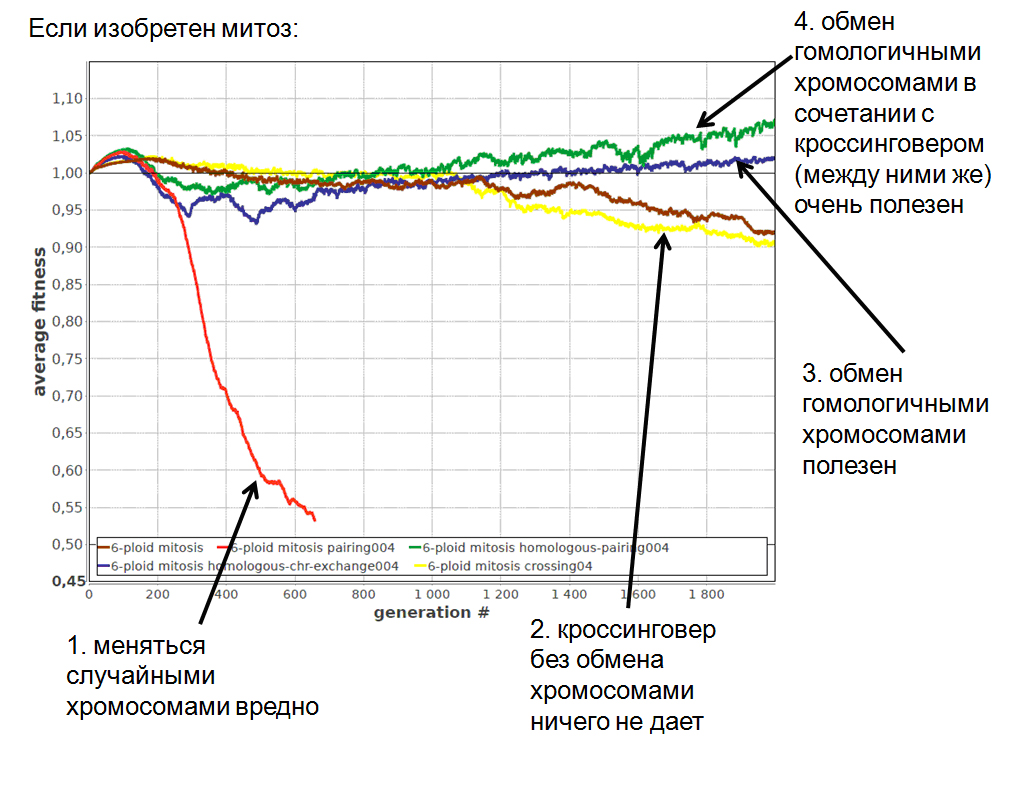
Рис. 6. Моделирование подтверждает, что после изобретения митоза меняться случайными хромосомами и перемешивать их фрагменты становится невыгодно, потому что этот унаследованный от предков способ рекомбинации вступает в конфликт с процессом специализации хромосом. Обмен самыми похожими хромосомами при этом остается выгодным, а кроссинговер усиливает полезный эффект от такого обмена. Коричневая линия — популяция 6-плоидов, имеющих митоз; остальные линии подписаны на рисунке. Изображение из обсуждаемой статьи в Biology Direct
Очевидное решение проблемы состоит в развитии клеточного слияния (которое, возможно, уже имелось у предков эукариот, ведь его наличие предполагается у Haloferax) и спаривания гомологичных хромосом двух клеток с кроссинговером и последующим их аккуратным распределением по дочерним клеткам. Важно, чтобы в ходе клеточного деления, следующего за попарной рекомбинацией хромосом, каждая дочерняя клетка получила строго по одной хромосоме из каждой гомологичной пары. Уже имеющийся механизм митоза является идеальной заготовкой (преадаптацией) для эволюции такого клеточного деления. Итак, мы получили сингамию и мейоз.
Изучение молекулярных механизмов мейоза привело специалистов к выводу, что мейоз наверняка развился на основе митоза (см.: На пути к разгадке тайны мейоза — популярный синопсис статьи Ю. Ф. Богданова «Эволюция мейоза одноклеточных и многоклеточных эукариот. Ароморфоз на клеточном уровне»). Также есть веские аргументы в пользу того, что эволюция мейоза началась с развития механизма спаривания гомологичных хромосом, и что смысл этого новшества был не в том, чтобы повысить интенсивность рекомбинации, а в том, чтобы ограничить ее, запретив рекомбинацию между непохожими хромосомами (A. S. Wilkins, R. Holliday, 2009. The evolution of meiosis from mitosis). Наша гипотеза хорошо согласуется с этой идеей и объясняет, откуда взялась потребность в такой адаптации.
6. Предлагаемый эволюционный сценарий
Итак, мы предлагаем следующий сценарий происхождения эукариот:
1) Предки эукариот были полиплоидными археями, не имевшими митоза. Они жили в мутагенной среде на мелководьях во времена Великого кислородного события. В таких условиях быть полиплоидом полезно в краткосрочной перспективе, но чревато вымиранием в долгосрочной.
2) Отбор способствовал выработке адаптаций, уменьшающих негативные эффекты полиплоидности при сохранении ее преимуществ. В результате был выработан набор средств, снижающих риск вырождения полиплоидов в мутагенной среде: унификация хромосом путем генной конверсии; интенсивный ГПГ между родственниками (спаривание с образованием цитоплазматических мостиков, возможно — временное слияние клеток); циклы плоидности (периодические редукционные деления).
3) Поскольку эти относительно простые «полумеры», видимо, не решили проблему полностью, в дальнейшем развивались еще более эффективные средства защиты от генетической деградации: обмен целыми хромосомами в сочетании с рекомбинацией; переход от конверсии к кроссинговеру, поскольку он эволюционно стабилен и позволяет осуществлять генетический обмен часто; замена кольцевых хромосом линейными.
4) Наконец, был изобретен митоз — аккуратное распределение хромосом при делении, так что каждая дочерняя клетка стала гарантированно получать ровно одну копию каждой родительской хромосомы. Это сразу сняло проблему сегрегационного груза. Однако обмен хромосомами в сочетании с кроссинговером по-прежнему был весьма полезен, так что у предков эукариот не было оснований от него отказываться.
5) Специализация и диверсификация хромосом, являющаяся неизбежным следствием изобретения митоза полиплоидами, постепенно стала вступать в конфликт со старыми способами безвыборочного генетического обмена и рекомбинации. Эти способы «устарели», и отбор способствовал их модернизации. В результате развились механизмы, обеспечивающие обмен, спаривание и рекомбинацию только очень похожих (гомологичных) хромосом. В конечном счете это привело к развитию сингамии и мейоза, то есть настоящего эукариотического секса.
6) Одновременно должно было происходить совершенствование механизмов выбора брачного партнера, потому что когда вы подходите к межорганизменной рекомбинации столь серьезно, спариваться с кем попало становится опасно. Подобно тому, как хромосомы стали спариваться для обмена участками только с очень похожими хромосомами, клетки должны были начать спариваться только с клетками, имеющими такой же хромосомный набор. Результат — появление «биологических видов» с хорошо перемешиваемыми и в меру изолированными генофондами. Но пока механизмы выбора партнеров были еще несовершенны, эукариоты могли нахватать много генов от неродственных линий (что они и сделали, см.: Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016).
7. Испытание на прочность
Всякая гипотеза, чтобы получить признание, должна пройти проверку временем и новыми фактами. Это — дело будущего. Впрочем, несколько испытаний наша гипотеза уже благополучно прошла в процессе подготовки к печати.
Во-первых, в 2015 году, когда работа над моделью уже была в разгаре, появилось сообщение о том, что у архей обнаружена строгая корреляция между полиплоидностью и наличием гистонов (S. K. Spaans et al., 2015. The chromosome copy number of the hyperthermophilic archaeon Thermococcus kodakarensis KOD1). То, что у некоторых архей есть гистоны, было известно давно, и этот факт всегда считался важным аргументом в пользу того, что предками эукариот были именно археи. Новые данные показывают, что этот аргумент приложим только к полиплоидным археям. Связь между гистонами и полиплоидностью, по-видимому, объясняется тем, что гистоны помогают упаковывать множество копий генома в одной маленькой прокариотической клетке.
Во-вторых, из нашей гипотезы вытекает одно очень специфическое проверяемое следствие. Если эукариоты произошли от полиплоидных архей, которые приобрели митоз, это обязательно должно было привести к массовому появлению новых семейств близкородственных генов (паралогов) у базальных эукариот (подобно тому, как это происходит у эукариот в результате полногеномных дупликаций, но только в большем масштабе). Как выяснилось, сравнительная геномика подтверждает это предсказание: еще в 2005 году Евгений Кунин и его коллеги показали, что на заре эволюции эукариот был период массового приобретения новых семейств паралогичных генов (K. S. Makarova et al., 2005. Ancestral paralogs and pseudoparalogs and their role in the emergence of the eukaryotic cell).
В третьих, один из рецензентов указал нам на несоответствие, состоящее в том, что все известные полиплоидные археи относятся к эвриархеям, тогда как эукариоты ближе к другой группе архей — кренархеям (к которым относятся, в том числе, и ближайшие родичи эукариот — локиархеи). Все кренархеи, чья плоидность известна, являются либо моноплоидами либо, самое большее, диплоидами. Плоидность локиархей неизвестна, ведь этих микробов никто даже не видел — известны только их геномные последовательности. Но, поскольку среди кренархей полиплоидов не обнаружено, предположение о полиплоидности локиархей выглядит маловероятным.
На помощь пришли гистоны. Поскольку было показано, что плоидность у архей коррелирует с наличием гистонов, можно посмотреть, нет ли гистонов у локиархей. В оригинальной статье с описанием локиархей гистоны не упоминаются. Однако в декабре 2015 года появилось сообщение об идентификации в геномах локиархей нескольких гистонов, да не каких-нибудь, а самых замечательных — переходных по своей аминокислотной последовательности между гистонами эвриархей и эукариот (B. Henneman, R. T. Dame, 2015. Archaeal histones: dynamic and versatile genome architects).
Так что пока всё сходится, и сам я сейчас процентов на 95 уверен, что эукариоты действительно произошли от полиплоидных архей, которые изобрели митоз.
Источник: Alexander V. Markov and Ilya S. Kaznacheev. Evolutionary consequences of polyploidy in prokaryotes and the origin of mitosis and meiosis // Biology Direct. 2016. V. 11. P. 28 (открытый доступ).
Журнал Biology Direct использует уникальную систему открытого рецензирования: мало того, что рецензенты не анонимны, так еще и их рецензии публикуются вместе со статьей. Поэтому заинтересованный читатель может по приведенной ссылке ознакомиться не только с текстом статьи, но и с реакцией на нее ведущих специалистов по биоинформатике и геномике, галоархеям, биохимии и происхождению жизни.
См. также:
1) На пути к разгадке тайны мейоза. Популярный синопсис статьи Ю. Ф. Богданова «Эволюция мейоза одноклеточных и многоклеточных эукариот. Ароморфоз на клеточном уровне» // Журнал общей биологии. 2008. Т. 69. № 2. С. 102–117.
2) А. В. Марков, А. М. Куликов, 2009. Происхождение эукариот как результат интеграционных процессов в микробном сообществе.
3) Для эукариот более важны гены, полученные от архебактерий, «Элементы», 15.10.2010.
4) Альтернативный способ репликации ДНК оказался эффективнее традиционного, «Элементы», 12.11.2013.
5) Новооткрытый микроб заполняет брешь между прокариотами и эукариотами, «Элементы», 12.05.2015.
6) Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016.
7) Обнаружены одноклеточные организмы с ядром, но без митохондрий, «Элементы», 18.05.2016.
-
-
Да-с - мощно задвинулЪЪ АлександрВладимирычЪ!
На Нашей памяти - это первый случай на Элементах, когда авторы пишут синопсис собственной статьи в серьёзном международном журнале. Если эта традиция будет продолжена - будет действительно круто.
Тогда прочие отечественные СМИ, например, уже вполне могут ссылаться прямо сюда как на источник.
-


>полиплоидами являются, по-видимому, все галоархеи (к которым относится Haloferax), метаногены и термоплазмы.
Т.е. все эвриахеи? Но эукариоты вроде как ближе к кренархеотной ветви?..
>Второй способ внутригеномной рекомбинации
Лучше поменять на "вариант", поскольку слово "способ" уже применено к самой "внутригеномной рекомбинации" - читатель может запутаться.
Ксатати: а каким способом пользуются дейнококки и нейссерии? Первым ("циклами плоидности")?
>Но со временем это закономерно привело к новому конфликту, для разрешения которого пришлось изобрести мейоз.
Прям диалехтика какая-то, простиг-споди! :~]
-
Статья написана неплохим языком, настолько неплохим, что впору поставить задачу подправить его до хорошего литературного. Ритмичность фраз, пропорциональность и длина предложений - всё настолько неплохо, что кто-то должен специально сесть за доводку. Рассказы Ю.Нагибина и М.Тарковского вам в помощь!
Спасибо за понятия "очищающий отбор" и "сегрегационный груз", утащу их себе в культурологию. -
По первому вопросу: см. в самом конце новости. Во-первых, у локиархей, ближайших современных родичей эукариот, обнаружены гистоны, да не просто гистоны, а промежуточные между гистонами эвриархей и эукариот.
Во-вторых, у архей обнаружена строгая корреляция между полиплоидностью и наличием гистонов. Поэтому есть основания надеяться, что локиархеи - полиплоиды, хотя все кренархеи с экспериментально установленной плоидностью - моноплоиды или диплоиды.
С остальным согласен. Про дейнококков и нейссерий - не знаю. Нейссерии активно используют генную конверсию для борьбы с иммунной системой, может и для унификации копий генома тоже.-
Да, дочитал - теперь вижу. И про нейссерий похоже так и есть: пока писал камент спешил и сам вспомнить деали исходной статьи не успел.
Но вот локиархеи как таковые (как и большинство прочей прокариотической "темной материи" некультивантов) у меня вызывают наибольшие сомнения - уж очень у них всё гладко и богато, а культур или просто какой специфичной микроскопии - нету.
Нужны независимые подтверждения другими группами. И другими методами.
-
-
В такой форме Ваш тезис выглядит как оксюморон.
А если его упростить, то будет "эукариотность - это одна из форм жизни клеточных организмов". Что уже будет практически тавтологией. Или просто банальным школьным определением.
Так что полезной информации из него читателю вряд ли удастся выловить.
Кроме того: хоть родство их и несомненно и уже может быть прослежено достаточно плавно, архитектурные различия сейчас настолько велики, что трудно представить что-то более различное в мире клеточной жизни. Поэтому тезис и по факту понимания не добавляет.
И еще я подумал что рано записывать промежуточные звенья эволюции одноклеточных в вымершие. Судя по разным статьям которые читал, глубоко под земной поверхностью могут существовать разные очень консервативные сообщества организмов, а пространства для микрожизни там огромные. Вот эта например статья произвела на меня большое впечатление, тут срок не так велик, но однако же
http://elementy.ru/novosti_nauki/432540/Mikrobiota_uglenosny
Можно ли эту судьбу побороть без изобретения митоза? Например, увеличить частоту горизонтальных переносов до такого уровня, чтобы успеть сменять хромосомы до полного истощения в цитоплазме запасов продуктов А, B и C.
-
А антиоксиданты. Нет кислорода нет и антиоксидантов. Появляется кислород, и надо обзавестись антикоскидантами, а иначе свободные радикалы атакуют клетки. А это процесс не сиюминутный. И пока не обзавелись, уровень мутаций мож и повышается.
ААААаа. Я гений. осинило. Супероксидная дисмутаза же у бактерий, архей и эукариот разная. (Или нет) Мож всё происходило одновременно, и появление еукариот и супероксидной дисмутазы.
> среди кренархей полиплоидов не обнаружено
В том же абзаце пишете, что кренархеи таки бывают диплоидами. Действительно ли требуется высокая степень плоидности? Мог ли весь предлагаемый сценарий произойти на диплоидах?
Предполагаю, что именно появление мейоза привело к расцвету многоклеточных – к эдиакарию. Многоклеточные организмы не могут осуществлять ГПГ. Только секс позволяет им перемешивать гены.
-
> Предполагаю, что именно появление мейоза привело к расцвету
> многоклеточных – к эдиакарию. Многоклеточные организмы не могут
> осуществлять ГПГ. Только секс позволяет им перемешивать гены.
Вроде как, многоклеточные грибы и растения появились задолго до эдикария. Да и с ГПГ у них не всё так уж плохо (без оного не было бы плацентарных, а, значит и нас с вами), хотя до вакханалии прокариот им, в этом плане, конечно, всё равно далеко.-
Тут, кстати, интересно было бы ещё оценить как изменилась вирусная нагрузка с обретением внутренней компартментализации - всё-таки это дополнительные барьеры для инфекции. И довольно серьёзные.
И как эти изменения связаны с описанным механизмом: не могло ли половое размножение развиться в т.ч. и как замена трансдукции, который, как описано и в "Логике Случая", является ведущим механизмом ГПГ, поскольку коньюгация и трансформация распространены на порядки реже?-
На счёт трансдукции, я думаю, вряд ли. Её основной эффект - внесение в геном новой информации, в то время как половой процесс направлен, скорее, на противостоние дегенеративным явлениям в геноме и его рихтовку в плане косметических адаптивных изменений. Так что, одно едва ли может эффективно заменить другое.
-
-
Мейоз существует не только у многоклеточных. Множество видов простейших практикуют его при конъюгации. Так что появился он задолго до первых многоклеточных.
-
> но случаев его все равно известно немало.
Теперь представим, сколько известно случаев полового размножения у многоклеточных. Не менее чем 10²⁷, я полагаю. Можно ли в этой ситуации сказать «ГПГ отсутствует»?
> появился он задолго до первых многоклеточных
Согласен. Мейоз появился у одноклеточных, а затем попал к многоклеточным (возможно, посредством ГПГ). Но когда именно – надёжных сведений нет.
Взглянем на рисунок 20 на странице http://scisne.net/a-303?pg=10
Примерно 900 млн. лет назад стала захораниваться неокисленная органика. А в течение миллиарда лет до того не происходило ничего столь же заметного. Что за событие случилось 900 млн. лет назад?
Моя гипотеза: появление секса привело к разнообразию и к распространению колоний многоклеточных, например водорослевых лугов.
Да, не эдиакарий, а заметно раньше.
-
Да, легенда объясняет. Но способ раскраски неправильный.
Вот, здесь же в легенде есть утверждение: «каждый ген сохранился в более или менее неиспорченном состоянии только на одной хромосоме». Проверим это утверждение. Какие аллели считаем неиспорченными? Пусть это аллели с качеством 0.9 и лучше. Значит нужно покрасить все аллели, которые 0.9 и лучше. Что мы тогда увидим? Увидим, что в седьмом локусе проверяемое утверждение всё же нарушается.
А способ покраски, применяемый уважаемым автором, маскирует факт нарушения. Больше того, при этом способе внешний вид раскраски был бы таким же, даже если заполнить ячейки случайными числами – точно так же в каждом столбце оказалась бы по одной зелёной ячейке.
А C - value paradox, когда от количества ДНК не зависит сложность организма, а размер клетки. Если общий геном разрастался, то и размер клетки мог бы разрастатса.
А мутации еще могли быть потому, что микробы еще не научились справлятся с кислородом и свободными радикалами, и только постепенно обзаводились антиоксидантами. Упоминается что по выходу на сушу надо было искать новых антиоксидантов, так как селена и иода не хватает.
Вообще то представляю как одна хишная клетка всё захватывала и разросталась всякими митохондриями.
Мож я тут в прямом эфире вижу важнейшее открытие.
Я это назвал самой скучную в истории статью про секс;) Вообще, тот же принцип возможно можно применить и в экономике. Гос апарат это делает. Начальник счётной палаты РФ и начальница гос контроля вроде дружили. И обменивались опытом. И наши и ваши налоговые инстанции говорит одни и те же слова. А вот экономичаская сфера скорее нет. А если компании не конкурируют, то почему не обмениватся ДНК. То что человек учится от природы не в новинку. Ну и вообще, в политике по принципу гмо, с введением полезных генов.
http://asdam.livejournal.com/122018.html
вот тут закралась маленькая опечатка:
В 2010 году была предложена красивая гипотеза, связывающее воедино происхождение эукариот...
наверно надо поправить на "связывающая"
По-моему, гораздо лучшее объяснение - это происхождение ядра вместе с митозом, мейозом и прочим от наследства лизогенного поксвируса. Лизогенных вирусов и плазмид, которые поддерживают одну копию на клетку и синхронизируют репликацию с делением хозяина, известно много. Известны также их варианты, которые имеют механизмы расхождения пар копий по дочерним клеткам, похожие на митоз, а уж плазмид, которые заставляют хозяев обмениваться генетическим материалом, множество - в том числе тех, которые встраивают хромосому хозяина в себя.
Кроме того, происхождение ядра от крупного лизогенного вируса объясняет еще и особенности репликации эукариотной ДНК, происхождение ядерных пор, и собственно линейные хромосомы эукариот. Ничего этого гипотеза полиплоидии кольцевых хромосом архей как толчка к митозу и мейозу не объясняет.
-
А как правильно разделять гены так, чтобы в копии оставались все дупликаты материнской клетки? Это же непростой процесс.
Для кольцевого генома такого способа не имеется.-
Я не очень понимаю ваш вопрос. Для кольцевого генома не имеется способа разделять хромосомы так, чтобы в копии оставались все гены материнской клетки? Имеется, и не один - ведь прокариоты с кольцевыми геномами именно это и делают. То же делают многие лизогенные плазмиды.
У прокариот и плазмид есть минимум четыре типа систем разведения копий по дочерним клеткам. Как правило, в них участвуют белковые нити, участки ДНК для прикрепления этих нитей, и белки, собирающие весь механизм - в общем это довольно похоже на митоз эукариот. У некоторых прокариот и плазмид есть даже нечто вроде клеточного центра.
Или вы имеете в виду что-то другое?
Ну, а гипотеза классная. Очень красивая и стройная, на первый взгляд, во всяком случае.
Объясните, пожалуйста, почему этих ужасов не происходит у бактерий и обычных (неполиплоидных) архей.
Последние новости










Рис. 1. Схема жизненного цикла полиплоидной (в данном случае — триплоидной) бактерии, не имеющей митоза. Три кольцевые хромосомы, каждая из которых содержит весь геном и может отличаться от двух других отдельными мутациями, реплицируются (удваиваются) в произвольном порядке, пока число хромосом не станет равно шести. После этого клетка делится на две дочерние триплоидные клетки. Поскольку митоз отсутствует, каждой из дочерних клеток достаются три случайно выбранные хромосомы из шести (при наличии митоза каждой «дочке» досталось бы ровно по одной копии каждой из трех родительских хромосом). Ori — точка начала репликации. Рисунок из статьи Anna H. Chen et al., 2012. Spatial and Temporal Organization of Chromosome Duplication and Segregation in the Cyanobacterium Synechococcus elongatus PCC 7942