«Эволюция. Классические идеи в свете новых открытий». Главы из книги

Глава 1. Наследственность: куда катится мир? (отрывок)
Отбор — игра по правилам
Методом случайного поиска (случайных блужданий) найти в необозримой «белковой вселенной» одну конкретную точку, соответствующую оптимальному выполнению белком данной функции, совершенно нереально. К счастью, такие оптимумы, как правило, — не точки, а обширные области. А главное, поиск оптимума в ходе эволюции осуществляется вовсе не методом случайных блужданий. Он идет методом направленного отбора. Направленность возникает благодаря тому, что случайные отклонения в «неправильную» сторону, ухудшающие рабочие качества белка, отбраковываются, отменяются и забываются, тогда как случайные отклонения в «правильную» сторону запоминаются и сохраняются.
Если исходная последовательность уже находится у подножия некоей возвышенности на ландшафте приспособленности — там, где уже есть хотя бы небольшой наклон (это значит, что белок хотя бы в минимальной степени, но уже выполняет какую-то функцию), отбор загоняет последовательность на вершину горы с поразительной эффективностью. Это можно показать с помощью компьютерной программы, впервые описанной Ричардом Докинзом в книге «Слепой часовщик». Пусть в роли исходной последовательности выступает произвольный набор букв, например, такой:
Программа размножит эту последовательность в тысяче экземпляров, внося в нее случайные мутации с заданной частотой. Например, пусть каждая буква в каждой копии с вероятностью 0,05 мутирует, т. е. заменяется другой случайной буквой. В результате каждая двадцатая (в среднем!) буква у каждого потомка будет отличаться от родительской. Зададим оптимум, к которому нужно стремиться, — некую осмысленную фразу. Попробуем сначала прийти к оптимуму без помощи отбора, методом случайных блужданий. Для этого из 1000 потомков выберем одного наугад и снова размножим в 1000 экземпляров с мутациями. И еще раз, и еще.
Вот что у нас получилось. Справа указаны номер поколения и степень отличия от целевой последовательности, т. е. удаленность от оптимума:

Бесполезно! За 100 поколений — ни малейшего прогресса, никакого приближения к оптимуму. Фраза осталась такой же бессмысленной, какой и была. Случайное блуждание в пространстве последовательностей, которое мы сейчас смоделировали, практически не дает шанса найти конкретную точку в белковой вселенной.
Нет, конечно, будь у нас бесконечный запас времени, когда-нибудь мы все же наткнулись бы на оптимум. Когда, вот в чем вопрос. Мы использовали 26 букв латинского алфавита и пробел, итого 27 знаков. Длина фразы — 60 знаков. Оптимум — это одна комбинация из 2760 (≈7,6 × 1085) возможных. Вариантов больше, чем атомов во Вселенной. Примерно столько поколений нам и потребуется, чтобы случайно наткнуться на нужную точку.
К такому способу поиска вполне приложимы известные рассуждения об урагане, пролетевшем над свалкой, который ни за что не соберет из мусора «боинг-747», и об обезьяне, которая, бессмысленно стуча по клавиатуре, никогда не напишет «Войну и мир». Таким способом — и впрямь не напишет. К счастью для нас, эволюция идет вовсе не этим способом. Или, если быть совсем уж точными, не только этим способом.
Мы все-таки не зря моделировали случайные блуждания — у них тоже есть аналог среди эволюционных процессов. Именно так идет нейтральная эволюция. Так меняются последовательности (аминокислотные или нуклеотидные), от которых приспособленность организма не зависит и которые поэтому не находятся под действием отбора.
Между прочим, хоть блуждания и случайные, некие закономерности можно заметить и в этом случае. Обратите внимание, что эволюционирующая последовательность постепенно становилась все менее похожа на исходную. Фраза из поколения № 1 отличается от исходной только тремя знаками, в поколении № 2 мы видим уже семь отличий, в поколении № 4–13, в пятом поколении — 16 отличий. К 50-му поколению никакого сходства с исходной последовательностью не осталось. Но в течение первых 20–25 поколений сходство сохранялось, постоянно уменьшаясь. Поэтому мы могли по степени этого сходства примерно определить номер поколения, к которому принадлежит данная фраза. Мы могли, сравнив данную фразу с исходной и зная скорость мутирования, примерно оценить, сколько поколений разделяет эти две фразы! На этом принципе основан замечательный метод молекулярных часов, с которым мы познакомимся в следующем разделе. А пока вернемся к отбору.
Изменим немного нашу программу. Пусть теперь для размножения выбирается не любая из 1000 последовательностей наугад, а лучшая — наиболее похожая на «оптимальную». Все прочее оставим как было. Запускаем программу и получаем следующее:
Ну вот, совсем другое дело! Под действием отбора наша последовательность уверенно двигалась к оптимуму — взбиралась на пик ландшафта приспособленности — и достигла его в 89-м поколении, с чем мы ее и поздравляем.
Главный урок из этих упражнений состоит в том, что эволюция под действием отбора совсем не похожа на попытки напечатать «Войну и мир», случайным образом нажимая на клавиши. Отбор — мощная организующая сила, которая придает эволюции направленность, формируя из хаоса случайных мутаций закономерный, упорядоченный результат.
Но позвольте, не отступили ли мы от реальности, произвольно задав оптимальную последовательность — фразу, к которой нужно было стремиться? Не похоже ли это на «божественное вмешательство»?
Вовсе нет. Для любой полезной функции, выполняемой белком при данных условиях (температуре, составе среды и т. д.) действительно существует одна или несколько оптимальных аминокислотных последовательностей, которые справятся с этой функцией лучше всего. Возможно, реального белка с такой последовательностью еще нет в природе — эволюция не успела его создать, — но ведь идеальная последовательность все равно существует, подобно тому как потенциально существовал химический элемент углерод со всеми своими свойствами еще до того, как первые атомы углерода начали синтезироваться в недрах звезд, вспыхнувших в молодой Вселенной. Отбор будет двигать эволюционирующую последовательность к этому идеалу независимо от того, есть уже на свете такие белки или им еще только предстоит появиться.
В чем мы действительно погрешили против истины, так это в том, что в неявном виде ввели в модель несколько допущений о форме ландшафта приспособленности, сильно упрощающих реальность. Мы допустили, что ландшафт — это одна большая гора с гладкими склонами и единственной вершиной, причем любая случайная последовательность, с которой начинается эволюция, уже находится на склоне этой горы, так что движение «вверх» (к идеалу) повышает ее приспособленность. Нахождение на склоне означает, что исходная последовательность хоть чуть-чуть, хоть совсем плохо, но все-таки уже справляется с данной функцией.
Рассмотренная модель справедлива лишь для ситуации, когда отбору уже есть за что «зацепиться», когда эволюционирующая последовательность уже на что-то годна.
Как удается эволюционирущим последовательностям переходить с одной горы на другую и со склона на склон? Иными словами, как белок, уже приспособившийся выполнять какую-то функцию — начавший взбираться по склону одной из гор, — может приобрести другую функцию, т. е. перейти на другую гору? Как он преодолеет разделяющую их низменность?
Это важный вопрос, и мы к нему еще вернемся. Ну а пока ограничимся несколькими замечаниями.
Во-первых, действительно, чем выше белок забрался по одному склону, тем меньше у него шансов перейти на другой. Глубокие низины между горными массивами, соответствующими основным группам белковых функций — так называемым надсемействам белков, — как правило, непроходимы. Крайне редко удается преодолеть пропасть одним отчаянным прыжком — макромутацией. В подавляющем большинстве случаев такие прыжки в горной местности кончаются понятно чем. Но все же бывают и удачные приземления. И тогда всем на диво какой-нибудь фермент, миллиарды лет занимавшийся превращением одного углевода в другой, вдруг превращается в кристаллин — белок хрусталика глаза, а пищеварительный фермент трипсин — в белок-антифриз, защищающий кровь антарктических рыб от замерзания1.
Но это исключения. Как правило, эволюционное движение большого и сложного современного белка ограничено одним горным массивом — одной группой родственных функций. На больших высотах ландшафт приспособленности белков сильно разобщен, фрагментирован, так что с одного массива на другой попасть почти невозможно. Но вот на малых высотах, у подножья величественных горных массивов, простирается холмистая местность, передвигаться по которой простым и примитивным древним белкам было намного легче. Среди искусственно синтезированных коротких белковых молекул с произвольной последовательностью аминокислот можно найти молекулы, выполняющие — пусть и с низкой эффективностью — различные функции, выполняемые природными белками в клетке. Это значит, что, передвигаясь по этим низинам, можно наткнуться на предгорья какого-нибудь горного массива даже методом случайных блужданий!
Скорее всего, основные функции белков, соответствующие белковым надсемействам, были «нащупаны» еще в самом начале эволюционного становления генетического кода и синтеза белка у древних РНК-организмов2. Потом эти функции бесконечно совершенствовались и подразделялись на множество вариаций. Эволюционирующие последовательности взбирались все выше по склонам «своих» горных массивов, разбредаясь по развилкам и отрогам, и чем выше они поднимались, тем меньше оставалось у них шансов перейти с однажды выбранной горной системы на какую-то другую.
По-видимому, только для самых простых и коротких (но при этом все же полезных) белковых молекул существует реальная вероятность возникновения «из ничего», «на голом месте» — из случайной комбинации нуклеотидов, случайно закодировавших некоторую последовательность аминокислот. Такой способ формирования новых белков исчерпал себя еще в РНК-мире. С тех пор новые белки появляются только из старых — не с нуля, а путем модификации того, что есть. Это одно из главных правил эволюции. Почти все новое — это перекроенное старое.
Нейтральные мутации и генетический дрейф — движение без правил
Ландшафт приспособленности — образ яркий и полезный, но, как и всякая модель, он несовершенен. Многие аспекты эволюционного процесса с его помощью отразить трудно или невозможно. Реальный ландшафт приспособленности переменчив (впрочем, как и реальные горные системы). Если один белок в клетке изменился, это хоть немного, но изменит свойства клетки, ее поведение и внутреннюю среду — а значит, и «требования», предъявляемые отбором к другим белкам. Их ландшафты приспособленности станут немного другими. Изменение одного вида в сообществе неизбежно повлияет на факторы отбора, действующего на другие виды, и т. д.
Кроме того, трудно представить себе такой ландшафт, который точно отразил бы реальное соотношение вредных, полезных и нейтральных мутаций. Как правило, большинство мутаций нейтральны — по крайней мере у высших организмов, эукариот, у которых в геномах много участков, слабо влияющих на приспособленность. На втором месте по численности вредные мутации, на последнем — полезные. Нелегко вообразить склон такой формы, чтобы из каждой точки было больше разных путей, параллельных горизонту, чем путей, ведущих вверх или вниз. Но реальные склоны реальных ландшафтов приспособленности именно таковы.
Нейтральные мутации — это по определению такие мутации, которые не влияют на приспособленность, или, что то же самое, не подвергаются действию отбора. Нам пора познакомиться поближе с этим самым распространенным в природе классом мутаций. Забудем на время о ландшафте приспособленности и обратимся к другой модели, которая описывает процессы, происходящие с генетическими вариантами (аллелями) в популяции.
Допустим, у нас есть маленькая популяция мюмзиков из 40 особей. Для простоты примем, что мюмзики гаплоидны, т. е. имеют одинарный набор хромосом — один-единственнный экземпляр генома, а не два, как у нас с вами, диплоидных организмов. Достигнув возраста в один год, каждый мюмзик рожает нескольких детенышей, а потом сразу умирает от старости. Все детеныши одинаковые, но выжить из них могут только 40 — больше мюмзиков просто не помещается в старой железной бочке на дне пруда, где живет изучаемая популяция. Смертность детенышей случайна — ровно 40 случайно выбранных везунчиков станут взрослыми и через год произведут потомство.
Поскольку смертность случайна, приспособленность мюмзиков определяется только их плодовитостью, т. е. числом детенышей. Плодовитость зависит от генотипа. Допустим, у мюмзика есть один ген, влияющий на плодовитость. Обозначим его буквой А. Между прочим, не смейтесь: такая модель вполне годится для изучения некоторых законов популяционной генетики.
Некогда у всех мюмзиков был только один вариант (аллель) гена А. Обозначим его А1. Но потом у одной особи произошла мутация, и в результате появился второй вариант гена — аллель А2. Допустим (опять-таки для простоты), что в нашей популяции, когда мы приступили к ее изучению, у половины мюмзиков ген А был представлен первым вариантом, у другой половины — вторым. Таким образом, частота аллеля А2 равна 0,5 (q2 = 0,5) и частота аллеля А1 такая же (q1 = 0,5).
Нам нужно ответить на вопрос: как будет меняться со временем частота аллеля A2, если мутация, которая привела к его возникновению, была нейтральной?
Раз мутация была нейтральной, значит, плодовитость обладателей обоих аллелей одинакова. Допустим, они все рожают ровно по десять детенышей. Разумеется, потомство наследует родительский аллель гена А.
Может показаться, что раз аллели имеют одинаковую приспособленность, то их так и останется поровну. Это неверный ответ. На рисунке показан результат четырех запусков нашей модели. Мы видим, что частота A2 во всех четырех случаях хаотически колебалась (по-научному такие колебания как раз и называются случайными блужданиями). Блуждания продолжаются до тех пор, пока частота аллеля не упрется либо в верхнюю «точку невозврата» (q2 = 1, частота аллеля достигла 100 %, аллель зафиксировался в генофонде), либо в нижнюю (q2 = 0, частота упала до нуля, аллель элиминировался из генофонда).

Так бывает всегда. Если аллель нейтрален, его частота будет «случайно блуждать» между нулем и единицей до тех пор, пока не упрется либо в верхний, либо в нижний предел. Рано или поздно нейтральный аллель либо зафиксируется (достигнет частоты 1), либо элиминируется — исчезнет из генофонда. Третьего не дано. Таким образом, блуждания хоть и случайны, их исход предсказуем. Мы точно знаем, что дело кончится либо фиксацией, либо элиминацией. Чем больше популяция, тем в среднем дольше придется ждать исхода3, но он все равно неизбежен.
Здесь самое время сказать, что случайные, не зависящие от приспособленности изменения частот аллелей называются генетическим дрейфом. Под властью дрейфа находятся все нейтральные мутации (а в маленьких популяциях отчасти также и вредные с полезными, но об этом позже).
Можно ли вычислить вероятность того, что нейтральный аллель в конце концов зафиксируется, а не элиминируется? Да, это просто. В нашем примере аллели A1 и A2 исходно находились в равном положении, потому что у обоих была частота 0,5. Очевидно, что в такой ситуации шансы на фиксацию у них должны быть одинаковыми и равными 0,5. В половине случаев A1 зафиксируется, A2 элиминируется, в половине — наоборот.
Ну а если конкурирующих нейтральных аллелей не два, а, скажем, четыре и начальные частоты у них тоже одинаковые (0,25)? В этом случае дело кончится фиксацией одного из аллелей и элиминацией трех остальных, причем для каждого аллеля вероятность фиксации равна 0,25. Таким образом, очевидно, что вероятность фиксации нейтральной мутации в будущем просто-напросто равна ее частоте в данный момент: Pfix = q.
Если вы, дорогие читатели, еще не устали от этой примитивной математики, то позвольте познакомить вас еще с двумя простыми, интересными и полезными формулами.
Сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении? (Имеются в виду мутации уже не в одном и том же, а в разных генах.) Если мы сумеем это вычислить, то получим прекраснейший инструмент — молекулярные часы. Тогда мы сможем по количеству нейтральных генетических различий определить, когда жил последний общий предок сравниваемых организмов.
Вывод этой формулы — подлинный шедевр «биологической математики». Судите сами. Определим сначала, сколько новых мутаций появляется в популяции в каждом поколении. Будем считать для простоты, что подавляющее большинство мутаций нейтральны (это недалеко от истины). Ответ очевиден: U × N, где U — темп мутагенеза (среднее число новых мутаций у каждой новорожденной особи), N — численность популяции. Определить U можно, просто сравнивая геномы детей и родителей.
Теперь нужно понять, какая часть из этих U × N только что появившихся мутаций в итоге зафиксируется. Это и будет искомая величина — темп фиксации нейтральных мутаций в генофонде популяции за одно поколение. Обозначим ее буквой V.
Мы уже знаем, что вероятность фиксации мутации равна ее частоте: Pfix = q. Чему же равна частота только что появившейся мутации? Это совсем просто. Поскольку мутация только что появилась, она пока есть только у одной особи. Следовательно, ее частота равна 1/N. Вот, собственно, и все. Умножаем количество новых мутаций на вероятность фиксации каждой из них (т. е. на долю мутаций, которые в итоге зафиксируются) и получаем ответ: V = U × N × 1/N. Поразительно! Численность популяции волшебным образом сокращается, и величина N уходит из уравнения. Мы приходим к выводу, что темп фиксации нейтральных мутаций не зависит от численности и равен просто-напросто скорости мутагенеза: V = U. Не знаю, как вы, дорогие читатели, а мы, биологи, приходим в восторг от такой красоты.
Нам остался один шаг до молекулярных часов. После того как два вида, произошедшие от общего предка, разделились, в их генофондах независимо накапливаются нейтральные мутации. За время t (измеряемое в поколениях) первый вид накопит V × t = U × t нейтральных мутаций, и второй вид накопит столько же. Совместными усилиями они накопят 2U × t нейтральных отличий друг от друга. Зная темп мутагенеза U и подсчитав число различий между геномами сравниваемых видов (обозначим его буквой D), определяем время жизни их последнего общего предка: t = D/2U. Это и есть знаменитые молекулярные часы.
Если бы темп мутагенеза и скорость смены поколений были одинаковыми у всех живых существ, все было бы совсем просто. Но они, конечно, неодинаковы, поэтому в вычисления приходится вносить поправки. А еще есть проблема насыщения: в какой-то момент — обычно по прошествии десятков, а чаще сотен миллионов лет — гены разделившихся когда-то видов оказываются настолько «переполнены» нейтральными различиями, что величина D перестает расти, хотя нейтральные мутации продолжают фиксироваться. Ниже мы увидим пример исследования, показавшего, как уровень сходства между «случайно блуждающими» последовательностями приблизился к минимально возможному и дальше снижаться уже не мог.
К счастью, разные участки генома накапливают нейтральные изменения с очень разной скоростью4. Быстро меняющиеся участки ДНК используются для датировки недавних событий, а те, что меняются медленно, хороши для датировки событий глубокой древности.
Дрейф и отбор: кто кого?
Генетический дрейф царствует над нейтральными мутациями (аллелями), отбор — над полезными и вредными. Отбор, повышающий частоту полезных мутаций, называют положительным. Отбор, отбраковывающий вредные мутации, — отрицательным, или очищающим.
Впрочем, все так хорошо и просто только в больших популяциях. В маленьких ситуация сложнее, потому что отбор и дрейф — две главные движущие силы эволюции — начинают конкурировать друг с другом за контроль над слабовредными и слабополезными мутациями.
Посмотрим, как это происходит. Воспользуемся для этого той же моделью с мюмзиками. Изменим только характер мутации, которая привела к появлению аллеля A2. До сих пор мы считали, что мутация была нейтральной. Поскольку она была нейтральной, обладатели аллелей A1 и A2 имели одинаковую плодовитость. Давайте теперь предположим, что мутация была полезной, что она повысила плодовитость мюмзиков на 5 %. Это можно смоделировать так: пусть мюмзики с генотипом A1 рожают по 20 детенышей, а мюмзики с генотипом A2 — по 21. Начальная частота аллеля A2 пусть будет по-прежнему равна 0,5. Только теперь мы рассмотрим популяции с разной численностью (N).
На рисунке (с. 54–55) показано, как будет меняться частота A2 в том случае, если его носители имеют пятипроцентное адаптивное преимущество. Мы видим, что в большой популяции (N = 5000) частота A2 неуклонно растет, приближаясь к единице. Так работает отбор в идеальных для него условиях, т. е. в больших популяциях, где влияние дрейфа на полезные и вредные аллели пренебрежимо мало. Форма у кривой довольно правильная, и это наводит на мысль, что ее можно описать какой-нибудь математической формулой. Это действительно так, но выводить формулу мы не будем, чтобы не утомить читателей (а любители математики могут сделать это самостоятельно)5.
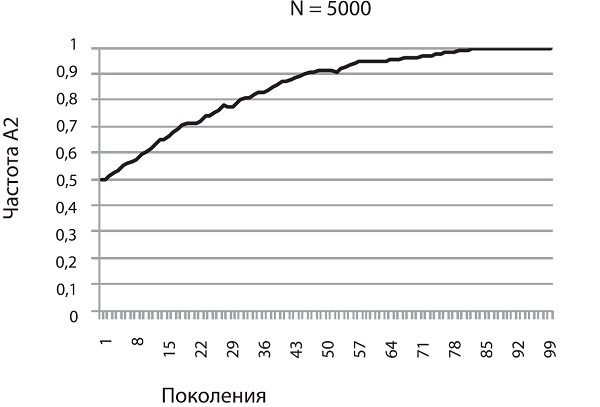
Перед нами, между прочим, важнейший эволюционный процесс — аллельное замещение, т. е. вытеснение более приспособленным аллелем менее приспособленного. Процесс идет не слишком быстро. В большой популяции, например состоящей из миллиона особей, для того чтобы зафиксировалась новая полезная мутация, дающая 5-процентное адаптивное преимущество, требуется около 560 поколений. А ведь 5 % — это серьезное преимущество. Такие мутации — редкость. Ждать, пока зафиксируется мутация с преимуществом в 1 %, придется уже 2800 поколений! Тем не менее в большой популяции отбор «чувствует» даже самую незначительную разницу в приспособленности. Это обеспечивает эффективную (пусть и медленную) фиксацию слабополезных мутаций и элиминацию слабовредных6.
На срееднем графике мы видим, какая судьба ждет такую же полезную мутацию, повышающую приспособленность на 5 %, в популяции меньшего размера (N = 200). Частота полезного аллеля и в этом случае росла и в конце концов достигла 1 (мутация зафиксировалась), но путь был труден и извилист. Пожалуй, нашей мутации могло и не повезти, ее шансы на фиксацию не были стопроцентными.

Наконец, на этом графике мы видим, что происходит с точно таким же полезным аллелем в совсем крошечной популяции (N = 30). На рисунке показаны результаты двух запусков модели. В одном случае мутация зафиксировалась, в другом — элиминировалась. Не правда ли, картинка похожа на результат работы дрейфа, а не отбора?
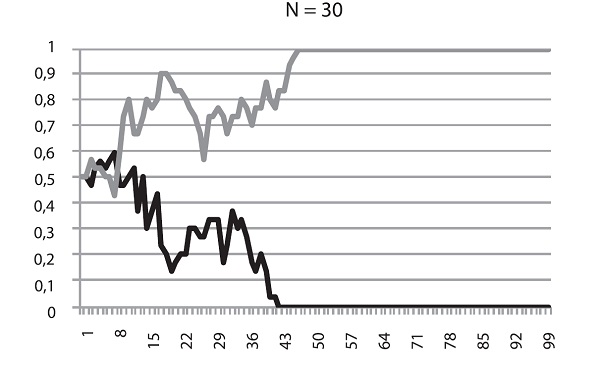
Так оно и есть. В этом заключается главный урок, который мы можем извлечь из наших экспериментов. Чем меньше популяция, тем слабее в ней власть отбора и тем могущественее дрейф. В маленьких популяциях слабополезные и слабовредные мутации начинают вести себя фактически как нейтральные. Их частоты «случайно блуждают», пока не наткнутся на верхний или нижний порог. В маленьких популяциях, где царствует дрейф, небольшие различия в приспособленности становятся невидимыми для отбора. Поэтому слабовредная мутация запросто может зафиксироваться, а слабополезная — элиминироваться.
Хорошо это или плохо? В большинстве случаев, конечно, плохо. Потеря слабополезных мутаций мешает маленькой популяции приспосабливаться к меняющимся условиям. Бесконтрольное накопление слабовредных мутаций и вовсе может поставить ее на грань вымирания. Именно в этом, между прочим, видят биологи одну из причин того, что крупные животные вымирают в среднем чаще, чем мелкие. У крупных животных, таких как носороги или слоны, популяции не могут быть такими же большими, как у мышей или насекомых. Это снижает приспособляемость крупных животных.
Но у этой медали есть и обратная сторона. У маленьких популяций больше шансов выйти из «ловушки локального оптимума», т. е. сползти с невысокого пика ландшафта приспособленности и взобраться на другой, повыше. Ведь отбор гонит организмы вверх и только вверх. Если популяция велика и малейшее различие в приспособленности «заметно» для отбора, спуск по склонам становится невозможен. Однажды взобравшись на одинокий холм, большая популяция уже никогда с него не слезет. Что касается дрейфа, то он ведет организмы по ландшафту приспособленности хаотическим образом, не замечая подъемов и спусков. Если популяция невелика и дрейф силен, у организмов есть шанс иногда двигаться не только вверх, но и немного вниз (и в сторону). Спустившись в ложбинку, организмы могут «обнаружить», что отсюда есть другой, более перспективный подъем. Если, конечно, не вымрут раньше, чем на него наткнутся.
Глава 7. Переходные формы
Проблема переходных форм беспокоила Дарвина, настораживала его сторонников и неизменно радовала оппонентов. Забегая вперед, скажем, что сейчас для экспертов-биологов эта проблема видится совсем с другой стороны, хотя все еще настораживает малоинформированных приверженцев эволюции и, как и прежде, возбуждает ее противников. В этой главе мы разберем, почему изменилось отношение биологов к переходным формам, а эмоции неспециалистов оставим за скобками.
Переходные формы — это те, что выстраиваются в ряд постепенных переходов от предков к потомкам. Если известно, какой облик имел отдаленный предок, то переходными будут любые наборы промежуточных признаков между ним и потомками. На основе филогенетического дерева можно предсказать, какие промежуточные формы могли существовать (и поэтому могут быть найдены), а какие нет. В соответствии с научным методом сбывшиеся предсказания подтверждают теорию. Например, зная строение динозавров и птиц, можно предсказать кое-какие морфологические черты переходных форм между ними. Мы прогнозируем возможность найти останки животных, подобных рептилиям, но с перьями, или останки животных, подобных птицам, но с зубами и длинным хвостом. При этом мы предсказываем, что не будут найдены переходные формы между птицами и млекопитающими, например ископаемые млекопитающие с перьями или птицеподобные ископаемые с костями среднего уха как у млекопитающих. Иными словами, эволюционное дерево покажет нам, где и что искать, а чего не может быть и чего искать не стоит.
Кроме того, к переходным относят предковые формы со смешанными признаками дочерних групп. Если дочерние группы происходят напрямую от какого-то общего предка, то в его облике соберутся некоторые (не обязательно все) примитивные признаки. Другие его признаки могут оказаться, напротив, такими же продвинутыми, как у того или иного потомка. Такую форму можно тоже называть переходной — она совмещает в себе признаки нескольких линий в их примитивном состоянии. Впрочем, подобная ископаемая форма, скорее всего, при ближайшем рассмотрении окажется не прямым предком дочерних групп, а одним из рано ответвившихся и мало измененных потомков этого предка (потому что вероятность найти чьего-либо прямого предка в очень неполной палеонтологической летописи невелика). Такая форма может служить хорошим портретным приближением к общему предку.
Переходными называют также и виды с промежуточным состоянием сложного признака (если в качестве конечного состояния рассматривать то, что наблюдается у современных животных; с точки зрения наших потомков, нынешняя форма будет промежуточной по сравнению с их обновленным, изменившимся миром).
Суть проблемы переходных форм Дарвин обрисовал так. Если эволюция идет постепенно посредством отбора все более совершенных форм, то мы, казалось бы, должны повсюду видеть ряды бесконечных плавных переходов между формами. В действительности же мы чаще видим дискретные виды, которые либо совсем не могут скрещиваться между собой, либо делают это с трудом и неохотно. В крайнем случае, если нынеживущие «усовершенствованные» виды, победив в конкуренции, вытеснили своих менее приспособленных предков, эти последние должны бы найтись в ископаемой летописи. Пусть современные виды оказались удачливее и выносливее и пусть они теперь расселились повсюду, но ведь когда-то и их предки были победителями и проживали вольготными царями на своей территории. Это значит, что их останки должны во множестве захорониться, обратиться со временем в фоссилии (окаменеть) и стать достоянием палеонтологов. Но у Дарвина в середине XIX века было мало примеров ископаемых переходных форм.
Другая часть проблемы — это постепенное образование сложного признака. Казалось бы, глаз видит только потому, что в нем все части превосходно подогнаны друг к другу (на самом деле не все и не превосходно, но это уже мелочи). Легкие вдыхают и выдыхают, потому что вся грудная клетка сконструирована как совершенный вакуумный насос. Конструкция уха легка, изящна и математически технична, и потому ухо служит нам, позволяя улавливать мельчайшие оттенки голосовых эмоций, ориентироваться в пространстве и строить звуковые гармонии. Птичье перо — легкая и прочная маховая поверхность; без набора таких совершенных, упорядоченно налегающих друг на друга поверхностей птица не полетит...
Действительно, обретение четко видящего глаза — дело, кажется, исключительно трудное. Ведь его нужно было собирать постепенно, имея в самом начале лишь набор клеток, фиксирующих свет, а в итоге получить точный оптический прибор. И на каждом этапе эволюции этот протоглаз должен был с пользой служить животному, чтобы эволюция не бросила дело на полпути.

Представляя мысленно этот путь, неволей задумаешься, зачем бы понадобились животным все эти органы в полусобранном виде. Зачем нужен полуглаз, регистрирующий лишь нечеткие тени, негодное для полета перо, плохо слышащее ухо, дыхательные пузыри, не способные как следует вдохнуть? Стоит ли эволюции тратить на эти приспособления силы и время, если конечный результат неизвестен? А если и вправду был пройден весь путь от пигментного светочувствительного пятна до совершенного глаза, то должны быть переходные формы с полузрячими недоглазами, нелетающими перьями, плохо работающими легкими и тому подобными несовершенными органами. Существовали ли такие формы?
В «Происхождении видов», где этот вопрос обозначен, на него дан четкий теоретический ответ — да, должны были существовать и существовали! — и приведены примеры. Теперь нам известно гораздо, гораздо больше превосходных примеров — переходных форм с такими как будто «недоделанными» морфологическими конструкциями, которые тем не менее прилежно служили своим обладателям.
Таким образом, проблема переходных форм распадается на три вопроса. Первый из них — почему в современном мире не так уж много постепенных переходов между видами, почему большинство видов четко отделяются один от другого. Этот вопрос мы уже обсуждали в главе 6. Второй вопрос можно сформулировать так: почему в ископаемой летописи мало вымерших переходных форм, некогда вытесненных более приспособленными конкурентами? И наконец, последний: как может постепенно сформироваться сложный орган, который кажется полезным только в своей законченной форме? Два последних вопроса мы рассмотрим ниже.
Переходных форм много
До сих пор иногда приходится слышать удивительное утверждение, что, мол, переходных форм не существует. Это мнение — незамысловатое наследие позапрошлого века или же намеренное сокрытие (прежде всего от себя самого) фактических знаний. В действительности сегодня известно огромное число переходных форм — как-никак со времен Дарвина прошло полтора столетия. И все это время охотники за древностями не переставали находить все новые и новые формы, среди которых множество переходных. Нам теперь не приходится сомневаться в их существовании и, подобно Дарвину, убеждать оппонентов, что они все же были, объясняя их редкость неполнотой свидетельств о вымершей жизни. Полтораста лет назад было известно около 200 видов ископаемых, на сегодняшний день их описано примерно 250 тыс. Открыты не только тысячи новых видов, но появилась и набрала силу целая область знаний — тафономия. Тафономия — это дисциплина о закономерностях образования местонахождений с ископаемыми. Она объясняет, где, как и почему формировались слои с окаменелостями. Родоначальником этой науки был выдающийся палеонтолог и писатель И. А. Ефремов (1908–1972). Законы тафономии помогают предсказать, в каких местах, в каких слоях и породах нужно искать новую фауну, а где не стоит и силы тратить.
В некоторых случаях ископаемые переходные формы пока обнаружить не удалось. Например, нет следов эволюции предков шимпанзе (тафономия объясняет это отсутствием условий для образования окаменелостей во влажных тропических лесах), нет достоверных следов существования ресничных червей, а этот класс объединяет более 2500 современных видов (у них нет ни скелета, ни плотных оболочек, они, вероятно, слишком мягкие и нежные, чтобы сохраниться в ископаемом состоянии). Подобные пробелы в палеонтологической летописи — вещь неизбежная, но это, конечно, не повод для предположений, что шимпанзе и ресничные черви никогда не эволюционировали и появились недавно в готовом виде.
Вот несколько замечательных примеров переходных форм, которые недавно нашлись, несмотря на вековой скепсис относительно самой возможности существования подобных существ.
На полпути к камбале
«С маленьким мятым личиком на краю...» — так можно было бы вслед за Бабелем описать камбалу. И вправду, разглядывая ее, невольно подумаешь — эк ее перекорежило! За что ж ее, бедную, так! А с другой стороны, трудно придумать существо, более пригодное для жизни на морском дне. Она совершенно плоская, один бок посветлел и притворился брюхом, другой
потемнел и стал спиной. Спинной и анальный плавники вытянулись вдоль краев, превратившись в равномерную гибкую оборку. А глаза очутились на одной стороне головы: один глаз, тот, что должен был оставаться на светлой, псевдобрюшной, стороне, переместился на другой бок, на псевдоспину.
Личинки всех камбалообразных гораздо больше похожи на нормальных рыб: глаза у них, как и положено, располагаются по бокам на разных сторонах головы. В ходе индивидуального развития одна из глазниц постепенно смещается сначала на верхний край головы — на макушку, а затем на противоположную сторону.
Дарвин и его последователи предполагали, что изменения в строении черепа, наблюдающиеся во время роста малька, воспроизводят эволюцию группы. Дарвин допускал, что в данном случае мог работать «ламарковский» эволюционный механизм: предкам камбал, повадившимся лежать на боку, приходилось постоянно косить глаза, и прижизненные изменения, выработавшиеся у рыб в результате такой «тренировки», передавались потомству. В начале ХХ века, однако, стало ясно, что результаты тренировки наследоваться не могут. Рассчитывать можно только на естественный отбор. Иными словами, предки камбал, у которых «нижний» глаз от рождения был чуть-чуть сдвинут к макушке, должны были оставлять больше потомства, чем те, у которых череп был строго симметричен, а «нижний» глаз смотрел прямо в песок. Это предположение многим казалось неправдоподобным.
Если эволюция шла путем отбора небольших наследственных изменений, то глаз должен был перемещаться с одного бока на другой мельчайшими шажками. А какое преимущество могло дать предкам камбал небольшое смещение глаза, если он все равно оставался на нижней стороне головы и, следовательно, не мог ничего видеть, кроме песка? Такое строение казалось неадаптивным.
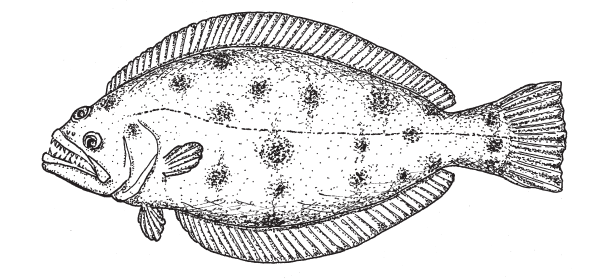
«Смещенный» глаз у псеттода находится на верхнем ребре головы. Другие примитивные признаки псеттода — спинной плавник, не доходящий до головы, и колючки в плавниках.
Самыми примитивными представителями камбалообразных до сих пор считались псеттоды (род Psettodes с всего двумя видами). У них «смещенный» глаз находится на верхнем ребре головы. Но все-таки этот глаз уже не зарывается в песок, когда рыба лежит на дне. А у той рыбы, которая, как предполагала постепенная эволюция, должна была существовать до псеттода, «брюшной» глаз бессмысленно смотрел бы в ил.
Поэтому камбалы были головной болью эволюционистов. Мало того что гипотетические переходные формы представлялись неадаптивными, так еще и не было никаких свидетельств их существования в прошлом. Была лишь упрямая уверенность, что они должны существовать, если теория верна. И вот в эоценовых отложениях (55,8-40,4 млн лет назад) Италии и Франции все же нашлись эти долгожданные и выстраданные в спорах примитивные камбалообразные. Их открыл и описал Мэтт Фридман из Чикагского университета. Первая из двух примитивных камбал была прежде неизвестна и при описании получила название Heteronectes chaneti. Вторую палеонтологи уже знали (это род Amphistium с двумя видами: paradoxum и altum), но Фридману впервые удалось точно реконструировать строение ее черепа при помощи рентгеновской томографии (Friedman, 2008).
Эти две рыбы имеют то самое строение черепа, которое скептикам казалось невозможным: череп резко асимметричен, одна глазница уже сместилась наверх, но оба глаза еще находятся по разные стороны головы. И один из них — да-да — смотрит в песок. Это явно не ошибка и не результат посмертных изменений ископаемых костей. Среди фактов, подтверждающих это, можно упомянуть асимметричное строение лобных костей у Heteronectes. Правая лобная кость крупная, прямоугольной формы, левая же превратилась в узкую изогнутую пластинку, ограничивающую верхний край сместившейся вверх глазницы. Род Amphistium известен по множеству экземпляров, и строение черепа у всех примерно одинаковое. Во всех случаях череп асимметричен, одна из глазниц смещена вверх, но остается на своей стороне черепа.
Все найденные экземпляры Heteronectes и Amphistium как минимум на порядок крупнее мальков современных камбалообразных на той стадии, когда глаза у них оказываются на одной стороне головы. Даже у псеттода глаз оказывается на верхнем ребре головы уже при длине тела 13 мм, тогда как длина типового экземпляра Heteronectes — 142 мм. Следовательно, не может быть и речи о том, что найденные переходые формы представляют собой мальков, у которых миграция глазницы еще не завершилась. Об этом же свидетельствуют и другие признаки, в том числе полное окостенение черепа и отсутствие возрастных изменений в положении глазницы у Amphistium в процессе роста от 103 до 200 мм. Несомненно, эти рыбы — взрослые и глазницы у них находятся в своем окончательном положении. Кстати, теперь мы можем констатировать, что Дарвин был прав и постепенные преобразования черепа в индивидуальном развитии современных камбал действительно воспроизводят ход их эволюции7.
Не только строение черепа, но и другие признаки Heteronectes и Amphistium показывают, что это самые примитивные из известных камбалообразных. В их скелете есть архаичные признаки, характерные для предков камбал — древних представителей группы окунеообразных. Некоторые из этих признаков не сохранились ни у кого из современных камбалообразных, другие сохранились только у псеттода (например, шипы в спинном и анальном плавниках).
Любопытно, что среди экземпляров Amphistium есть как «левые», так и «правые» особи. То же самое наблюдается у псеттода, однако у более продвинутых камбалообразных такие вариации крайне редки: каждый вид обычно представлен или только «левыми», или только «правыми» рыбами. Это пример жесткой зарегулированности онтогенеза. В начале эволюционного развития онтогенезу доставалось больше свободы в случайных вариациях, но в дальнейшем онтогенез стабилизировался и для случайности не осталось места (подробнее об этой общей закономерности эволюции онтогенеза мы поговорим в главе 8).
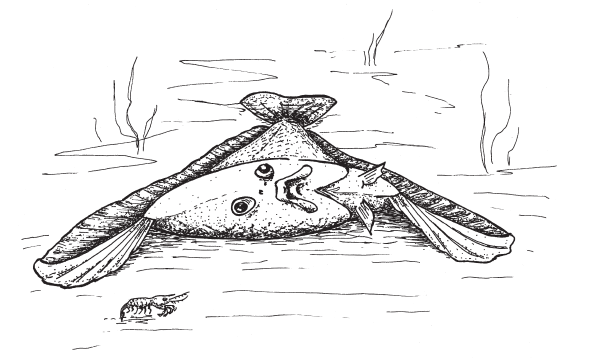
Опираясь на лучи спинного и анального плавников, древние камбалообразные приподнимали голову над песком и осматривали «нижним» глазом поверхность дна.
Ископаемые рыбы, описанные Фридманом, опровергают утверждение о невозможности и нежизнеспособности переходных форм между камбалами и нормальными рыбами с симметричным черепом. Однако хотелось бы еще и понять, как же все-таки древние камбалообразные использовали свой нижний глаз, который должен был, по идее, глядеть прямо в песок. Фридман отмечает, что подсказку можно найти в поведении некоторых современных камбалообразных, которые время от времени приподнимают свое тело над поверхностью дна, опираясь на лучи спинного и анального плавников. Heteronectes и Amphistium тоже могли «отжиматься» таким образом, поскольку спинной и анальный плавники у них были очень мощными. Приподнимая голову над песком, эти рыбы своим смещенным нижним глазом осматривали поверхность дна перед собой в поисках добычи.
Heteronectes и Amphistium не были прямыми предками современных камбалообразных. Они родственники этих предков, точнее — малоизменившиеся потомки тех рыб, что дали начало всем камбалообразным, современным и вымершим. Впрочем, то же самое можно сказать о большинстве ископаемых переходных форм. Вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, а даже если его и найдут, то еще попробуй докажи, что это точно сам предок, а не его троюродный племянник. Поэтому «по умолчанию» среди биологов принято такую возможность вообще не рассматривать. В результате любая переходная форма автоматически интерпретируется как «боковая веточка», а не «участок ствола». Но если веточка отходит от ствола недалеко, как в данном случае, называть ее «переходной формой» вполне корректно.
Первые камбалообразные появились в палеоцене (65,655,8 млн лет назад) и быстро разделились на множество эволюционных линий. В эоцене, когда жили Heteronectes и Amphistium, уже существовали более продвинутые камбалообразные с глазами на одной стороне головы. Таким образом, удивительные рыбы, описанные Фридманом, уже в эоцене были «живыми ископаемыми», т. е. мало изменившимися потомками общего предка группы. Если бы эволюционист воспользовался машиной времени и отправился в эоцен, он бы с удовлетворением наблюдал целый ряд переходных форм: тут и протокамбалы с глазами на разных сторонах головы, тут и псеттоды с глазом на макушке, тут и продвинутые камбалы с глазами на одном боку.
Что это за тип такой?
В начале xx века палеонтологи еще могли допускать, что жизнь на Земле началась в кембрийском периоде (542-488 млн лет назад). Это было связано с тем, что первые ископаемые фауны внезапно, будто бы из ниоткуда, обнаруживались в низах кембрийских слоев. Кембрийские животные являли взору биолога уже совсем готовые морфологические типы. Если это были членистоногие, то уже с изящными суставчатыми ножками, сегментированным панцирем и фасетчатыми глазами — готовые, настоящие членистоногие. Если моллюски — то моллюски готовые, с характерными раковинками, если брахиоподы — то тоже готовые, с раковинками, с отпечатками ловчих рук и мускулатуры, с прикрепительными ножками.
И Дарвин, и его последователи понимали, что таким продвинутым созданиям должен был предшествовать долгий предварительный этап развития. Но где же он? Под кембрийскими слоями лежали слои протерозоя, в которых теретически и должны были сохраниться остатки более древней и примитивной жизни. Но там, в протерозойских слоях, никто ничего интересного не находил. Поэтому загадка кембрийского взрыва — так называли внезапное появление разнообразных животных в нижних кембрийских слоях — долго казалась неразрешимой.
В течение ХХ века решение появилось. «Затерянный мир Дарвина», фауны докембрийских отложений, начали постепенно обнаруживаться. Самая известная из них получила в России название вендской, в других частях мира ее чаще называют эдиакарской. Оказалось, что в позднем докембрии уже существовали разнообразные мягкотелые многоклеточные организмы. Захоронения многоклеточных докембрийского возраста найдены в Китае, Австралии, Канаде, в Европе и в России (на Белом море). Так что кембрию пришлось довольствоваться более скромной ролью: из «начала начал» он превратился в «один из этапов».
И все же кембрийский период был революционным временем становления важнейших групп животных. Революцией было приобретение твердого скелета сразу многими группами организмов, как многоклеточными, так и одноклеточными, и поэтому они стали в изобилии сохраняться в ископаемой летописи. В кембрийское время фиксируется появление всех известных типов животного царства (членистоногие, моллюски, брахиоподы, хордовые, иглокожие и т. д.) и еще многих существ, которых не удается отнести ни к одному из них. Эти последние обладают смешанной морфологией. У них имеются признаки разных типов и классов: моллюсков и кольчатых червей, членистоногих и головохоботных червей, хордовых, полухордовых и иглокожих, ракообразных и хелицеровых. А есть и такие, строение которых вообще не дает подсказки о возможном родстве. Таких называют «проблематиками», не указывая таксономический тип.
В докембрийские времена проблематиков и животных со смешанной морфологией было еще больше, а о некоторых из них ученые даже спорят, животное это или лишайник. И дело тут не в том, что остатки редки или плохо сохранились или ученые не прилагают достаточных усилий, чтобы разобраться с ними. Нет, это объективная трудность, которую эволюция ставит перед биологами. Ведь в начальные периоды эволюции многоклеточных, до того как стабилизировались признаки основных филогенетических ветвей животного царства, существовали общие предки этих ветвей. И они, естественно, обладали смешанным набором признаков разных филогенетических линий. Из-за этого их невозможно уверенно соотнести ни с одной из них и уложить в прокрустово ложе современной систематики. Разные исследователи по-разному трактуют эти формы, то выделяя их в отдельные типы, то относя к известным типам в качестве «стволовых» (предковых) групп, то сближая с общими предками нескольких известных типов. Именно эти существа без определенной таксономической прописки дают наилучшее представление о переходных формах. Тех, что отмечают путь от предковых видов с широким диапазоном изменчивости к таксонам или группам видов, взявшим за основу своего плана строения только часть имевшихся возможностей.
Природа, сначала будто бы посмеявшись над морфологами и таксономистами и заставив их страдать от неопределенности, сама предложила подсказку. Подсказка предстала в виде серии лагерштеттов — местонахождений ископаемых с уникальной сохранностью. В лагерштеттах ископаемые сохраняют мельчайшие детали строения, и не только твердых частей, но и мягких. Это отпечатки покровов, мускулатуры, пищеварительных органов, кровеносной и выделительной систем — всего того, что так ценно для анатома и позволяет реконструировать вымерший организм, определить его место в системе животного мира. Лагерштетты, как это ни удивительно, особенно характерны как раз для кембрийских и ордовикских слоев, когда происходило становление современных типов и классов и когда жили все эти удивительные промежуточные формы. Большинство установленных ранних переходных форм известны именно из лагерштеттов.
Халкиерия
Халкиерия (Halkieria) — животное 5-6 см длиной, встречающееся в отложениях раннего и среднего кембрия (500–540 млн лет назад), сочетает признаки целых трех типов — кольчатых червей, моллюсков и брахиопод. Неудивительно, что халкиерию относили попеременно к разным таксономическим типам. История ее открытия и реконструирования весьма поучительна.
Halkieria — загадочное животное, встречающееся в отложениях раннего и среднего кембрия (500–540 млн лет назад) — сочетает признаки кольчатых червей, моллюсков и брахиопод и, возможно, является близким родственником общего предка этих животных.
Имя халкиерия изначально было дано гипотетическому ископаемому, от которого нашлись только плоские шипы. Считалось, что само животное было втиснуто в узкую щель шипа и должно было как-то удерживаться на субстрате в обтекающем его потоке воды. Эта реконструкция была не слишком убедительна, поэтому в конце концов решили, что шип — это не целый скелет животного, а только часть. Шипы халкиерии обычно находили вместе с плоскими чешуйками, причем у тех и других был сходный орнамент. Подсчитав соотношение чешуек и шипов, животное стали изображать в виде червя, сплошь покрытого чешуями и шипами. Это было все же лучше, чем живущий в струях воды непонятный монстр.
Но вот в одном из лагерштеттов Гренландии (место называется Сириус-Пассет) обнаружили отпечаток целого животного, у которого — о ужас! — были шипы халкиерии. И вот она, реальная халкиерия, — многосегментный червяк, слегка похожий на брюхоногого моллюска — слизня, покрытый рядами чешуй нескольких типов, а спереди и сзади пристроились вдобавок щитки, напоминающие раковинки брахиопод. Такую химеру, конечно, никакой самый смелый зоолог не мог себе представить. Зато теперь мы знаем, как примерно мог выглядеть общий предок моллюсков, брахиопод и кольчатых червей.

Перед вами — упрощенное дерево типов животного царства. На нем показано положение важнейших ископаемых форм, проливающих свет на происхождение и родственные связи современных типов.
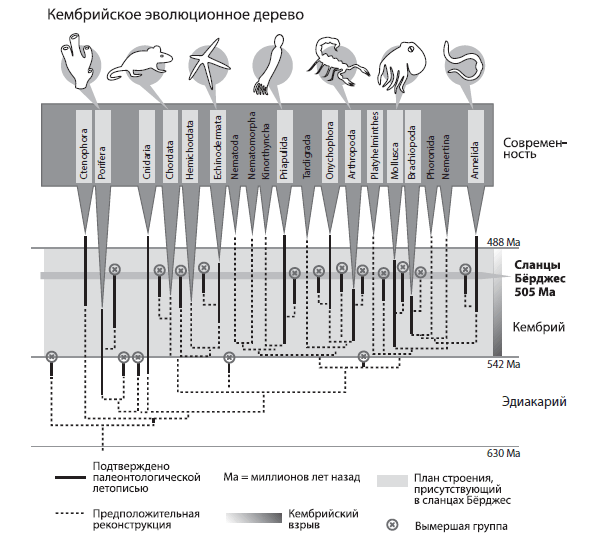
Появление типов животных в палеонтологической летописи и по филогенетическим реконструкциям; здесь показано положение на временной шкале знаменитого лагерштетта сланцы Бёрджес, первого местонахождения с остатками фауны мягкотелых и скелетных животных уникальной сохранности, открытого в 1909 году.
Кембрийская и вендская (эдиакарская) часть дерева изобилует тупиковыми веточками — вымершими формами, близкими к предкам одного или нескольких типов современных животных. Имеются веточки, отходящие не от мощных стволов сегодняшних типов, а от пунктирных, базальных частей дерева. Таких больше всего в венде и начале кембрия.
Большинство среднекембрийских переходных форм известны из лагерштетта, который называется сланцы Бёрджес (Burgess Shale). Открытый на юго-западе Канады Чарлзом Уолкоттом в 1909 году, это самый старый, самый известный и самый исследованный лагерштетт в мире.
Вот одна из характерных переходных форм, найденных в этом местонахождении. Это существо, как и халкиерия, сочетает в себе признаки кольчатых червей, брахиопод и моллюсков. Его описали Саймон Конвей Моррис, палеонтолог из Великобритании, внимательный, дотошный и смелый морфолог, и Жан-Бернар Карон, специалист из Канады, создавший виртуальный музей по сланцам Бёрджес (Conway Morris, Caron, 2007).
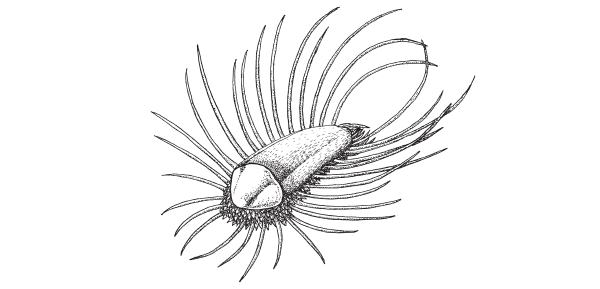
Orthrozanclus reburrus — морское животное, жившее 505 млн лет назад, — по-видимому, близкий родственник общего предка моллюсков и кольчатых червей. Из Conway Morris, Caron, 2007.
Причудливое создание длиной 6-10 мм получило название Orthrozanclus reburrus. Мягкое тело животного было покрыто сверху и с боков твердыми, но не минерализованными склеритами — етинками и шипами разнообразной формы и длины, располагавшимися в несколько рядов. Кроме того, передний конец тела был прикрыт небольшой раковиной. Животное вело донный образ жизни и ползало на брюшной стороне подобно слизню. Шипы и щетинки выполняли защитную и, возможно, сенсорную функцию. Назначение раковинки неясно. Может быть, она служила опорой для мышц ротового аппарата.

Wiwaxia — еще одно загадочное кембрийское животное: то ли моллюск, то ли родственник первых кольчатых червей.
Проблематичный Orthrozanclus reburrus несет признаки двух других проблематичных кембрийских групп — халкиериид (см. выше) и виваксиид, которые разными исследователями трактуются то как примитивные моллюски, то как кольчатые черви, то как брахиоподы. По современным представлениям, подкрепляемым сравнением ДНК, эти три типа являются родственными и объединяются в надтип Lophotrochozoa. Их важнейший общий признак — наличие особой плавающей личинки, трохофоры. К этому же надтипу относятся мшанки, эхиуриды, немертины и некоторые другие группы.
Шипы и щетинки у Orthrozanclus почти такие же, как у виваксиид, а раковинка — как у халкиериид. Несмотря на некоторые существенные различия (например, у халкиерии, в отличие от Orthrozanclus, кроме передней раковинки есть еще и задняя, а склериты минерализованы), авторы полагают, что их находка доказывает близкое родство и единое происхождение халкиериид, виваксиид и Orthrozanclus. Все эти формы они предлагают объединить в единую группу «халваксиид».
Положение халваксиид на эволюционном дереве надтипа Lophotrochozoa пока не может быть определено однозначно. Рассматриваются два возможных варианта: либо эта группа близка к предкам моллюсков, либо она ближе к общим предкам кольчатых червей и брахиопод. Впрочем, по большому счету, разница между этими гипотезами невелика, поскольку общие предки моллюсков, брахиопод и кольчатых червей сами были весьма близки друг к другу. Так что нет ничего удивительного, что нашлись вымершие животные с признаками всех трех типов: напротив, это яркий пример сбывшегося предсказания эволюционной теории.
«Микро» или «макро»?
Мы вплотную подошли к вопросу, почему биологам стало неинтересно доказывать, что переходные формы действительно существуют. И дело не только в том, что они существуют в предсказанном множестве и порядке, но и в том, что интерес к ним сместился теперь в другую плоскость. Сегодня нас интересует прежде всего, как происходили крупные изменения и возникали адаптации к совершенно новым условиям — например, при освоении суши и пресных вод первыми беспозвоночными, а затем и позвоночными, при освоении воздушной среды насекомыми и птицами, возвращение млекопитающих в океан. Можно ли проследить становление крупных групп по ископаемым? Дают ли окаменелости представление о маршрутах эволюционного творчества?
Если речь идет об обычном видообразовании, нетрудно представить себе те мельчайшие шажки, которыми век за веком шла эволюция и совершенствовались адаптации. В случае выхода на сушу или изобретения полета такие шажки вообразить труднее. Чтобы выживать на суше животным пришлось полностью перестроиться: заставить плавательный пузырь дышать, заставить сердце гнать кровь от пузыря прямо к голове, заставить тело опираться на ноги, а эти ноги заставить переступать, почки — беречь воду, уши и глаза — слышать и видеть в воздушной среде. И все это происходило постепенно, и каждое промежуточное звено было адаптировано к своей среде, без планирования и перспективного взгляда в будущее... да, представить себе такой маршрут шаг за шагом на первый взгляд непросто.
Вот в середине девона (393 млн лет назад) перед нами лопастеперые рыбы, а вот уже в верхнем девоне (383 млн лет) первые наземные четвероногие (тетраподы). Между ними всего в 10 млн лет (что не так уж много для геологической истории) уместилась целая адаптивная революция четвероногих обитателей суши. Действительно, изменения, требуемые для внедрения животного в принципиально новую среду, на первый взгляд кажутся более внушительными, чем выработка устойчивости к новому яду или паразиту. Размышления о масштабе событий заставляли биологов различать микро- и макроэволюцию (вплоть до идей о том, что это принципиально разные процессы, управляемые разными механизмами).
Но по мере развития биологии (включая экспериментальное изучение эволюции и наблюдения за случаями быстрой адаптивной радиации в природе) становилось все яснее, что макроэволюция — это череда тех же микрособытий, только длиннее (см. главу 2). Это суммарный результат множества последовательных микроэволюционных преобразований. Чтобы окончательно доказать этот вывод, важно изучать ископаемые переходные формы, показывающие возможность постепенных перестроек, из которых шаг за шагом складывается революционное изменение организма. Именно такие переходные формы являются сейчас объектом пристального внимания палеонтологов.
Чем тщательнее изучается эволюционный ряд, тем яснее становится постепенный ход изменений. Ведь если у исследователя имеются только начальное и конечное звенья цепи, то пропасть между ними кажется огромной, непреодолимой. И тогда перед лицом колоссальности задачи придумывают специальный термин «макро-» и пытаются изобрести особый макромеханизм для преодоления пропасти. Но если получше поискать и пристальнее присмотреться, то от начального до конечного варианта начинает просматриваться путь из последовательных «микро»-шагов. И здесь главным подспорьем, конечно, стали новые прекрасные находки палеонтологов. Многие важнейшие находки, заполнившие иллюзорные пропасти, — это находки в лагерштеттах. И не только кембрийских, но и мезозойских, сформированных не только морскими осадками, но и пресноводными.
Разумеется, разбираясь с макромасштабом, нельзя опираться на прямые наблюдения. Мы можем, как это показано в главах 5 и 6, наблюдать появление новых видов, но не новых семейств и отрядов — тех уровней классификации, которые, как правило, маркируют выход в новую адаптивную зону. Вы спросите почему — и правильно сделаете. Это закономерный вопрос. А ответ на него очень простой: не можем по определению. Дело вовсе не в том, что носители нового признака, сопоставимого с теми, по которым устанавливают крупные таксоны (семейства, классы и т. д.), не могут появиться на наших глазах. Это как раз пожалуйста. Но мы никогда не признаем такую форму представителем нового крупного таксона. Биологи не станут выделять в особый таксон существо, появившееся только что на наших глазах, как бы сильно оно ни отличалась от своих предков. Одна из причин в том, что неизвестны перспективы новой формы и ее новообретенного признака. Неизвестно, сможет ли на базе этого новшества появиться серия различных по морфологии и экологии видов. А может, новшество бесперспективно, его носители вскоре вымрут, и тогда их нужно классифицировать не как родоначальников крупного таксона, а как уродцев. Нравится нам это или нет, но крупные таксономические ранги биологи присваивают группе только в ретроспективе, когда группа уже «набрала силу», накопила запас изменчивости, разделилась на роды и виды, продемонстрировала свои отличные от других групп эволюционно-экологические возможности — одним словом, доказала всей своей историей, что заслуживает высокого ранга. Даже если в результате макромутации возникнет необычная форма с измененным планом строения, потребуются миллионы лет, чтобы выяснить, можно ли эту форму считать родоначальником нового крупного таксона или это был просто уродец, обреченный на вымирание, — аберрантный представитель родительского таксона, от которого так и не произошло ничего заслуживающего внимания.
Хорошей иллюстрацией может быть появление нового отряда мшанок Fenestellida. Эти мшанки отличаются от других формой колоний и высокой специализацией зооидов. Первый род этих мшанок появился в силуре. И в течение последующих 25 млн лет несколько видов мшанок с характерной фенестеллидной морфологией существовали одновременно со своими прародителями, скромно довольствуясь акваторией морей, омывающих Лаврентию (будущая Северная Америка). Только в начале каменноугольного периода эти несколько видов породили целый букет перспективных форм, которые расселились по всему миру. На базе «предложенной» первым силурийским родом морфологии они породили множество родов и семейств, составивших отряд Fenestellida. Если бы палеонтологи начали работать с силурийскими видами, то они, не зная дальнейшей истории, присоединили бы единичные виды с необычным фенестеллидным признаком к предковому отряду, выделив в отдельный род, но не более того. Но как только к ним в руки попадает все последующее разнообразие, то сразу появляется целый новый отряд. Так кто же возник в силуре и существовал без изменений целых два периода — новый род или отряд? Это вопрос, требующий формального решения и перспективного взгляда. Любая новая, отклоняющаяся форма может стать прародителем большого таксона, а может бесславно вымереть.
Так или иначе, начало крупного таксона — это всегда уклоняющийся от предковой морфологии вид, который получил какое-то преимущество, открывшее перед его потомками новые возможности. Это первый макроэволюционный шаг.
Вперед, в воздушный космос
Рассмотрим, как из рыб получились четвероногие животные, ставшие завоевателями суши: как рыбы отправились в наземный космос. Это событие датируется концом девонского периода, 385–360 млн лет назад. Первые четвероногие были не первыми существами, покинувшими водную среду: на суше в конце ордовика уже поселились растения, по-видимому, еще в кембрии в почвах появилась богатая фауна беспозвоночных, а еще раньше обитателями суши стали бактерии и грибы. Так что рыбы, пожертвовав гармонией со своей привычной водной средой и превратившись в медлительных, неповоротливых четвероногих, пришли в населенный и гостеприимный мир. Если в воде их подстерегали хищники, а за пищу приходилось конкурировать со многими голодными едоками, то на суше крупных хищников не было и привлекало обилие доступной пищи. Поэтому рискованный и трудный выход на сушу был выгодным предприятием. Это была гигантская незанятая экологическая ниша. И эволюция поспешила заполнить ее, как только представилась возможность.

Прототакситы (Prototaxites), конусовидные сооружения девонского периода высотой от 6 до 9 м. Как выяснилось, это гигантские вымершие грибы. Богатая грибная флора существовала гораздо раньше, не меньше миллиарда лет назад. Об этом свидетельствуют, в частности, исследования Константина Наговицына из Института нефтегазовой геологии и геофизики, выделившего разнообразные и необычные ископаемые грибы из протерозойских пород возрастом 1 млрд лет. Нужно помнить, что высшие растения могли начать освоение суши только «с ведома» своих непременных спутников — грибов и бактерий. Кто же еще, если не они, станет фиксировать для растений атмосферный азот и высвобождать необходимые растениям вещества из захороненной в почве органики? Ведь без этого невозможна жизнь наземного растения. Освоение суши растениями было подготовлено предшествующим заселением суши грибами и бактериями. Копия оригинального рисунка Ричарда Бизли (© Richard Bizley, bizleyart.com)
Возможность, по-видимому, обеспечили условия девонского периода. Палеоэкологическая интерпретация местонахождениий с девонскими переходными формами рисует нам мелкие, слабопроточные пресные водоемы субтропического или тропического пояса. Предполагается пониженное содержание кислорода в атмосфере и в воде. Возможно, из-за сильных периодических обмелений позднедевонским лопастеперым рыбам пришлось приспосабливаться к жизни в условиях, где вода больше не служит опорой для тела, потому что слишком мелко, где приходится дышать без помощи жабр атмосферным воздухом.
Эти водоемы облюбовали различные рыбы — и их было много, включая крупных и мелких хищников. Не случайно в девонских морях 17 % родов рыб приобрели панцирь — мирные обитатели обзаводились средствами защиты. Это было время расцвета рыб — помимо бесчелюстных, появившихся не позднее ордовика, в девонских морях плавали хрящевые и костные рыбы, разделившиеся еще в силуре. А костные рыбы в девоне были уже представлены двумя группами — лучеперыми и лопастеперыми. Эти группы различаются, помимо прочего, строением плавников: у лопастеперых плавник сидит на мясистом основании, скелет которого составляют удлиненные кости, плавниковая лопасть поддерживается серией симметричных костных пластин. У лучеперых плавниковые лопасти асимметричны, а кости основания плавника укорочены.
Именно одна из групп лопастеперых с мощными грудными и брюшными плавниками, напоминающими лапы — рипидистии, — дала начало наземным позвоночным. К лопастеперым относятся современные кистеперые (латимерия) и двоякодышащие (рогозуб, протоптер, чешуйчатник). Генетический анализ показал, что из современных рыб ближайшими родственниками тетрапод являются двоякодышащие.
Эволюционные изменения, происходившие с рипидистиями — предками первых тетрапод (земноводных), были постепенными. Из позднедевонских отложений известно более десятка представителей, сочетавших признаки земноводных и рыб. Они выстраиваются в относительно стройный ряд ископаемых форм: эустеноптерон (Eusthenopteron), пандерихт (Panderichthys), тиктаалик (Tiktaalik), элпистостега (Elpistostega), ливониана (Livoniana), эльгинерпетон (Elginerpeton), вентастега (Ventastega), метаксигнат (Metaxygnathus), акантостега (Acanthostega), ихтиостега (Ichthyostega), тулерпетон (Tulerpeton) и грирерпетон (Greererpeton). В этом ряду постепенно убывают рыбьи и накапливаются тетраподные черты, но разные системы органов совершают этот переход не одновременно, какие-то приходят к тетраподному состоянию быстрее, какие-то медленнее. Вслед за знаменитым шведским палеозоологом Эриком Ярвиком этих созданий часто называют «четвероногими рыбами».
Основное, что потребовалось изменить, чтобы получить право называться «настоящими четвероногими», — это конечности. Впрочем, четвероногость и хождение как таковое тоже возникли задолго до самих четвероногих. Ловко ходить по дну на четырех плавниках, приподнимая тело над грунтом, умели многие лопастеперые рыбы. Так что преадаптации для хождения имелись еще на «рыбьей» стадии. Но, чтобы ходить эффективно по суше, где тело весит гораздо больше, желательно все же иметь специализированную конечность. Нужно было сформировать подвижный сустав лапы и открепить пояс передних конечностей от черепа. Ведь у рыб пояс грудных плавников жестко крепится к задневисочной кости черепа, а это ограничивает движения и головы, и плавников.
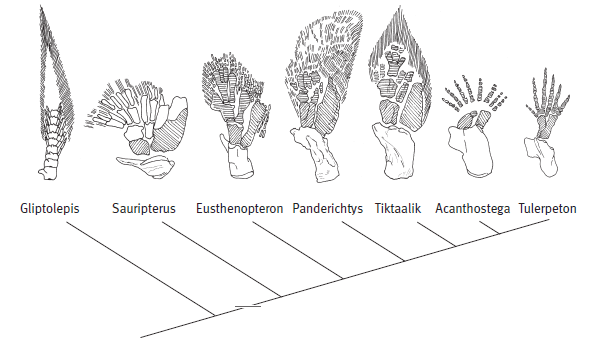
Схема изменения скелета передних конечностей у «четвероногих рыб».
Но если присмотреться повнимательнее, то окажется, что конечности (плавники) девонских лопастеперых рыб не так уж сильно отличались по своему плану строения от лап примитивных тетрапод. Хотя конечности сохранились не у всех ископаемых переходных форм, мы достаточно подробно знаем, что и как изменялось. Уже у эустеноптерона в переднем плавнике имелась кость, соответствующая плечевой кости тетрапод, и две кости, соответствующие локтевой и лучевой костям. Строение «дистальных элементов», соответствующих будущей кисти тетрапод, у эустеноптерона и пандерихта еще оставалось относительно неупорядоченным.
Далее в этом ряду поместился тиктаалик, обнаруженный в 2006 году американскими палеонтологами Эдвардом Дешлером, Нилом Шубиным и Фаришем Дженкинсом на острове Элсмир (арктическая Канада). «Тиктаалик» на языке эскимосов означает «большая пресноводная рыба, живущая на мелководье». И вправду, тиктаалик — это плоская, покрытая крупной чешуей рыба с крокодильей головой, на которой сверху сидят глаза, впереди две ноздри и большая зубастая пасть. У этой рыбы, как и у других представителей интересующего нас ряда переходных форм, часть черт сходна с лопастеперыми, а другие признаки сближают ее с четвероногими. Рыбьи признаки — это чешуя, плавниковые лучи, почти такие же, как у кистеперых, сложная нижняя челюсть и небные кости. Тетраподные признаки — укороченный череп, отделенная от пояса передних конечностей и потому относительно мобильная голова, наличие локтевого и плечевого суставов (Daeschler et al., 2006).
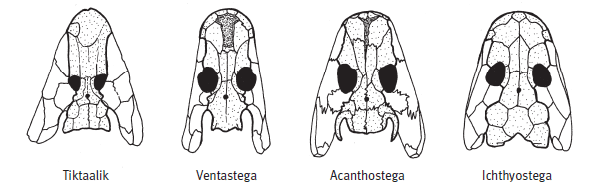
Черепа «четвероногих рыб» и первых тетрапод. Из статьи Ahlberg et al., 2008. Nature. V. 453. P. 1199–1204.
По сравнению с пандерихтом у тиктаалика скелет передних конечностей приобрел чуть более оформленный вид, так что становится уже более-менее понятно, откуда взялись у потомков тиктаалика косточки пясти и пальцев. Тиктаалик и пандерихт уже могли сгибать свой передний то ли плавник, то ли лапу в суставе, который у их потомков назовут локтевым.
А еще тиктаалик опробовал одно небольшое, но важное нововведение — он почти избавился от жаберной крышки, которая еще была у пандерихта (к функциям жаберной крышки мы еще вернемся).
Вместе с жаберной крышкой потерялась и жесткая связь между поясом передних конечностей и черепом. Голова освободилась от передних конечностей (или конечности от головы). То и другое стало более подвижным. Теперь можно было начинать учиться нормально ходить. У тиктаалика ребра сплюснулись, соединения позвонков окостенели. За счет этого он стал хуже сгибаться, зато тело приобрело устойчивость, что очень важно для ходячего, а не плавающего образа жизни.
Утрата жаберной крышки оказалась на редкость полезна в дальнейшем. Сочленение остатков жаберной крышки с головой вроде бы стало рудиментарным довеском для наземного, дышащего легкими существа. Но оно не исчезло вовсе, а начало медленное и крайне важное для всех наземных четвероногих путешествие внутрь черепа, превращаясь потихоньку в крошечные слуховые косточки. В действительности слуховые косточки начали формироваться еще до утери жаберной крышки. Как было показано Пером Альбергом и Мартином Бразо из Уппсалы, пандерихт, а не тиктаалик первым приспособил под примитивное стремечко (слуховую косточку) одну из костей сочленения жаберного аппарата с черепом (гиомандибулу). Эта кость была у него тонкой, изящной, она прилегала к дыхательному отверстию (первой жаберной щели), которое расширилось у пандерихта. У рыб это отверстие называется брызгальцем, оно имеется у эмбрионов тетрапод, но по ходу развития эмбриона становится полостью среднего уха и евстахиевой трубой. Пандерихт дышал воздухом, насосный аппарат жаберной крышки (см. ниже) ослабился, и части его переориентировались на выполнение другой функции — восприятие звука. От рыб, близких к пандерихту, стремечко унаследовали все тетраподы, включая нас (Brazeau, Ahlberg, 2006).
У вентастеги, которая по строению черепа занимает промежуточное положение между тиктааликом и акантостегой, от конечностей, к сожалению, почти ничего не сохранилось. Широкая форма рыла, строение черепной коробки сближают вентастегу с акантостегой, а форма и пропорции покровных костей черепа — с тиктааликом.
Широкая челюсть вентастеги была усажена мелкими острыми зубами. На длинном теле — около метра-полутора в длину — имелось две пары коротеньких конечностей с пальцами (сколько пальцев — неизвестно) и хвост с плавником, который поддерживали плавниковые лучи длиной около 7 см. Вентастега жила в солоноватоводных мелких прибрежьях и, имея внушительные размеры, охотилась на рыб. Как и у пандерихта с тиктааликом, у вентастеги часть бывшего сочленения жаберного аппарата с черепом была преобразована в слуховую косточку — стремечко. Куда относить вентастегу — еще к рыбам или уже к тетраподам, т. е. древнейшим амфибиям, — вопрос формальный, но если у нее действительно были пальцы, то, пожалуй, второй вариант предпочтительнее.
Следующие в ряду — акантостега и ихтиостега, которых принято считать уже «настоящими» тетраподами. Их конечности представляют собой всего лишь еще один небольшой шажок в сторону дифференциации дистальных частей скелета (стопы и кисти). У них окончательно оформились три крупные кости конечности (в передней лапе это плечо, локтевая и лучевая кости), тогда как многочисленные косточки кисти пока еще были мелкими и изменчивыми. Но у акантостеги уже точно были настоящие пальцы, а плавниковых лучей, которые еще сохранялись у тиктаалика, у нее не было. При этом число пальцев у акантостеги варьировало от пяти до восьми: признак еще не «стабилизировался». Изменчив он и у ихтиостеги. Более стабильное число пальцев характерно для тулерпетона (шесть пальцев); также у него имелись уже совсем «тетраподные» локтевая, лучевая и пястные косточки. У грирерпетона число пальцев стабилизировалось — теперь их будет всегда пять.
Кроме конечностей выход на сушу потребовал глубокого преобразования дыхательной и кровеносной системы. Опять же кажется, что переход от дыхания водой к дыханию воздухом невозможен мелкими улучшениями конструкции — нужно какое-то масштабное преобразование сразу обеих систем. Но это, как выясняется, не так. Подобно постепенному преобразованию конечностей так же постепенно происходил и переход от жаберного дыхания к легочному. И это можно увидеть на том же ряду ископаемых четвероногих рыб.
Итак, нужно сформировать легкие и, соответственно, два круга кровообращения и трехкамерное сердце. Такое строение кровеносной и дыхательной систем, судя по палеонтологическим данным, было чуть ли не у всех девонских лопастеперых рыб. Сохранилось оно и у современных двоякодышащих рыб. В качестве органа воздушного дыхания лопастеперые стали использовать особое впячивание передней части пищевода. Рыба заглатывает воздух, который отправляется в этот пузырь, оплетенный кровеносными сосудами, — это наипростейшее легкое.
У двоякодышащих пузырь выполняет две функции: примитивного легкого и примитивного гидростатического органа. У большинства (но не у всех) костных рыб раздувшееся выпячивание пищевода специализировалось для выполнения второй функции и превратилось в «настоящий» плавательный пузырь. Но у девонских рыб этот вырост пищевода совмещал функции легкого и регулятора плавучести.
В девонских водоемах обретение дополнительного органа дыхания было более чем оправдано: содержание кислорода в атмосфере было понижено, а доля углекислого газа, наоборот, повышена по сравнению с современностью. Поэтому в воде, особенно в стоячей, рыбам было трудно дышать одними жабрами. И они приобрели дополнительный орган для дыхания атмосферным воздухом. Так что для выхода на сушу уже велась «подготовка» исподволь -легкие для дыхания воздухом и прилагающаяся к ним модифицированная система кровообращения уже были припасены у девонских лопастеперых рыб. Разумеется, отбор поддерживал их как адаптации для жизни в бедной кислородом воде. О том, для чего они потом пригодятся, ни рыбы, ни отбор знать не знали.
Появление дыхательного пузыря -примитивного легкого -создало предпосылки для еще одного новшества: эффективного воздушного насоса. У костных рыб воду через жабры гонит жаберная крышка: рыба закрывает рот и поднимает среднюю часть крышки, увеличивая объем жаберной полости (ее мягкая оторочка плотно прижата к телу). Соответственно, давление в жаберной полости уменьшается и, когда рыба открывает рот, туда устремляется вода. Затем рыба закрывает рот, жаберная крышка опускается, оторочка отходит от края и вода вытекает наружу, омывая жабры.
Все это возможно в плотной водной среде, когда требуется обеспечить сквозной ток воды через жабры. В разреженной воздушной среде такой насос будет крайне неэффективным, ведь помимо разной плотности среды у дыхательного насоса и задачи другие. Нужно не гнать воздух сквозным потоком, а вентилировать слепой воздушный мешок. Но эту задачу первые тетраподы (земноводные) толком так и не решили. Это удалось лишь рептилиям. А у земноводных принцип нагнетания воздуха в легкие примерно тот же, что и у дышащих воздухом рыб. Они используют преобразованный аппарат жаберных дуг для расширения глоточной полости и, соответственно, для вдоха. Отработанный воздух из легких выдыхается за счет сокращения мускулатуры легких (а у двоякодышащих рыб он выходит за счет более высокого давления в воде по сравнению с воздушной поверхностью над водой, куда рыба выставляет голову для вдоха-выдоха).
Особенности жаберного аппарата, приспособленного для вдоха-выдоха за счет движения глотки, а не жаберной полости, хорошо отражаются в скелете и современных, и ископаемых девонских лопастеперых. Для более эффективного выведения углекислого газа из организма лопастеперые используют жабры, а земноводные, утратившие жабры, используют кожу («кожное дыхание»). Это создает массу ограничений, из-за которых земноводные так и не смогли далеко уйти от воды.
Но в воздушной среде жаберная крышка все равно не работала в качестве воздушного насоса, поэтому от нее пришлось отказаться, и это открыло наземным обитателям дополнительные возможности -косточки жаберной крышки ушли внутрь черепа. И начали формировать среднее ухо. Жаберная крышка частично редуцировалсь у тиктаалика. Это означает, что тот способ дыхания, которым продолжали пользоваться другие рыбы, стал ему больше не нужен. Тиктаалик дышал в основном воздухом при помощи легких и, возможно, кожей.
Раз у тиктаалика редуцировалась жаберная крышка, это означает, что ему пришлось научиться глотать без использования всасывающего жаберного насоса — рыбы втягивают частицы пищи с током воды. Тиктаалик глотал только за счет движения глотки и головы. У акантостеги, наследницы тиктаалика, еще сохранялись внутренние жабры, а у ихтиостеги они уже редуцировались (наружные жабры у личинок амфибий сохраняются и сегодня). Следовательно, последователи тиктаалика тоже глотали за счет движений головы и челюстей. В дальнейшем по мере редукции остатков жаберного аппарата у древних амфибий происходило постепенное развитие шейного отдела.
Таким образом, в строении дыхательной и кровеносной систем при переходе от рыб к тетраподам не было резких скачков, а большинство важных изменений произошло еще на «рыбьем» этапе. Каждое новшество в дыхании и кровообращении не создавалось на пустом месте, а лишь улучшало требуемые функции на основе развития имеющихся приспособлений.
Плавники превращаются в пальцы: взгляд генетика
Недавно было обнаружено семейство генов, необходимых для развития плавников у рыб и получивших название actinodin (and). У наземных позвоночных этих генов нет. Если отключить у рыбы гены and, то, во-первых, в парных плавниках не развиваются эластичные плавниковые лучи — лепидотрихии (но в хвостовом плавнике они развиваются нормально), во-вторых — меняется характер работы ряда ключевых генов-регуляторов (в том числе hoxd13a и sonic hedgehog), которые есть не только у рыб, но и у четвероногих и от которых зависит передне-задняя полярность зачатка конечности (будь то плавник или нога). Похожие изменения в работе этих генов-регуляторов у четвероногих, возникающие в результате некоторых мутаций, приводят к полидактилии (многопалости).
По-видимому, потеря генов and древними четвероногими привела к утрате лепидотрихий в парных конечностях, однако в хвостовом плавнике лепидотрихии могли сохраниться. Именно так обстоит дело у древнейших четвероногих, обнаруженных в ископаемой летописи, таких как акантостега и ихтиостега. У этих животных уже не было плавниковых лучей на парных конечностях, но они еще сохранялись в плавниковой складке, окаймлявшей хвост. У более ранних переходных форм, таких как тиктаалик, плавниковые лучи на парных конечностях имелись.
Логично предположить, что утрата генов and и вызванная ею дестабилизация работы регуляторных генов у древнейших четвероногих создала предпосылки для развития у них большого числа (до восьми) почти одинаковых пальцев. В дальнейшем у четвероногих восстановился исходный, «нормальный» характер работы генов-регуляторов, причем теперь их работа уже не зависела от утраченных генов and. Результатом этого могло стать автоматическое сокращение числа пальцев до пяти. Так изучение генетики современных животных помогает понять молекулярные механизмы событий, произошедших сотни миллионов лет назад (Zhang et al., 2010).
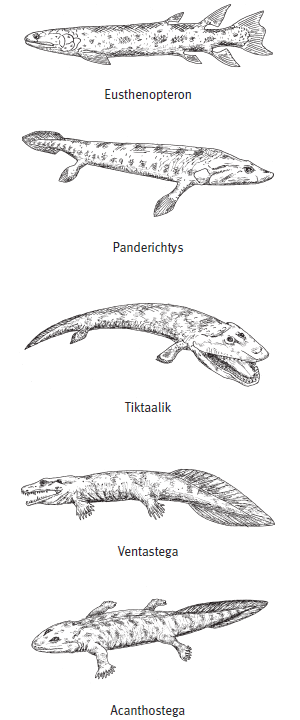
Реконструкции внешнего облика некоторых позднедевонских «четвероногих рыб». Сверху вниз: эустеноптерон, пандерихт, тиктаалик, вентастега, акантостега.
Мы рассмотрели ряд переходных форм, связывающих лопастеперых рыб с примитивными четвероногими. Совершенно непонятно, где в этом ряду находится искомый великий «макропереход». Приобретение выростов пищевода, в которые можно набирать воздух? Нет, это просто адаптация рыб к плохо аэрируемым водоемам. Аналогичные органы воздушного дыхания (например, в виде модификаций ротовой полости) возникали и в более поздние времена у разных рыб, оказавшихся в схожих условиях. Приобретение костей поясов конечностей у эустеноптерона? Едва ли, ведь это тоже была чисто «рыбья» адаптация к большей подвижности плавников. Может, назначить этот рубеж по приобретению подвижного сустава у лап, т. е. где-то между эустеноптероном и тиктааликом? Но ведь это просто адаптация рыб к ползанию по дну мелких луж и переползанию из одного пересохшего водоема в другой, развивавшаяся к тому же долго и постепенно. Еще есть вариант — присвоить «макро»-титул тиктаалику, у которого передние конечности отделились от черепа и стали свободными лапками... хотя и это всего лишь улучшенный вариант адаптации для передвижения по дну на мелководье. Утрата плавниковых лучей и приобретение пальцев? При большом желании можно, конечно, объявить эту небольшую модификацию дистальных участков конечностей тем самым эпохальным «превращением рыбы в амфибию». Действительно, именно в этом месте непрерывного ряда переходных форм специалисты проводят формальную грань между рипидистиями и тетраподами. Но не лучше ли честно признать, что палеонтологические данные показывают просто длинный ряд постепенных маленьких изменений, каждое из которых само по себе никак не тянет на «макрособытие»?
Всюду в этом ряду мы видим последовательные адаптации, служившие их обладателям верой и правдой в конкретных условиях. Ни одна из них по отдельности не заслуживает статуса «макроизменения». Сопоставимые по масштабу эволюционные преобразования происходят и в современной природе вокруг нас, и в лабораторных экспериментах, и в повседневной работе селекционеров. И уж конечно, если бы первые тетраподы вымерли, так и не дав начало большой и разнообразной группе животных, разумные осьминоги ни за что не выделили бы ихтиостегу, акантостегу и их родню в особый класс — считали бы их самое большее отдельным тупиковым семейством лопастеперых. И были бы правы.
Нестройными рядами — в сухопутное будущее
Завершая разговор о переходных формах между рыбами и тетраподами, нужно уточнить, что, располагая находки в один ряд, мы, конечно, сильно упрощаем реальность. В палеонтологии это типичная ситуация: пока переходных форм найдено мало, их удобно раскладывать в ряд, но чем больше находок, тем кустистее становится эволюционная схема. Вообще эволюция чаще похожа на куст, чем на ниточку, и многочисленные параллелизмы — ее типичнейшая черта (о параллельной эволюции и ее генетических основах мы говорили в главе 4).
На первый взгляд, последовательные изменения от рыб к четвероногим хорошо соотносятся с порядком положения переходных форм во временном ряду: раньше всех жили существа с преобладанием рыбьих признаков, затем с течением времени рыбьи черты терялись, сменяясь тетраподными. Пандерихты жили 385–383 млн лет назад, тиктаалик — 380–375, вентастега — 374–365, акантостега и ихтиостега — 370–365, грирерпетон — 359 млн лет назад.
Однако найдены и другие «четвероногие рыбы», нарушающие эту стройную картину. Например, Metaxygnathus, от которого найдена только нижняя челюсть. Его возраст — 374 млн лет. Он должен бы, судя по геологическому возрасту, иметь морфологию, близкую к тиктаалику, но его морфологическое место — после вентастеги. Очевидно, у Metaxygnathus развитие челюстного аппарата шло быстрее, чем у других, параллельно развивавшихся форм. При этом остальные его органы могли меняться медленее (хотя об этом мы можем пока только гадать). Неодинаковая скорость эволюции разных систем органов свойственна для стартового периода многих крупномасштабных эволюционных преобразований (выход рыб на сушу, происхождение членистоногих, птиц, млекопитающих, цветковых растений и т. д.). На этом этапе могут сосуществовать формы, у которых, например, система дыхания уже продвинутая, а ног еще нет, а могут быть и такие, у которых ноги уже вполне ходячие, а легкие еще рыбьи.
Разная скорость эволюции морфологических структур в разных линиях создает мозаику продвинутых и примитивных признаков, которую невозможно разложить в одну аккуратную линеечку. Можно видеть лишь общее направление изменений: как потихоньку вымирают промежуточные формы с преобладанием рыбьих черт, а выживают с преобладанием тетраподных.
В позднем девоне (385-359 млн лет назад) существовали представители разных линий рыб, устремившихся в своей приспособительной гонке к полуводному, полуназемному образу жизни. Позднедевонские окаменелости помогают реконструировать общее направление этой гонки, но не маршрут каждой из линий. То, что известно палеонтологам, — скудные фрагменты явно богатой фауны — пунктирно прорисовывает ход разных эволюционных линий, но не показывает связи между ними, их начала и концы. Поэтому нельзя утверждать, что все они появились и вымерли в позднем девоне, ведь нам известны только некоторые из штрихов пунктира.
Скорее всего, история четвероногих рыб началась не в позднем, а в среднем девоне (397-385 млн лет назад) или даже раньше. В связи с этим можно ожидать находки переходных форм с рыбьими признаками, немного разбавленными признаками тетрапод, в слоях среднего девона. О том, что позвоночные, возможно, приступили к освоению суши раньше позднего девона, свидетельствуют окаменевшие следы, оставленные каким-то шагавшим по суше животным. Эти следы найдены в Польше в слое возрастом 395 млн лет. В определении возраста слоя, в котором найдены эти следы, как будто нет оснований сомневаться. Слой сформировался в прибрежной морской обстановке. Может быть, это была лагуна или заливаемая равнина. На поверхности слоя видны даже ямки от дождевых капель и трещины от высыхания. Сами следы разные по размеру и морфологии; есть цепочки следов, есть и одиночные отпечатки. Самый большой след — 26 см в ширину; для сравнения, ихтиостега длиной около 2,5 м оставляла 15-сантиметровый отпечаток. В цепочке следов различаются отпечатки передних и задних лап — передние меньше задних.
Реконструкция шага по этим следам позволяет заключить, что животное двигалось, изгибая тело из стороны в сторону, примерно как саламандра. Следов волочения хвоста нет. Значит, уже были сформированы крестец и пояс задних конечностей, приподнявших тело над землей. Все это признаки четвероногого хождения (Nied wiedzki et al., 2010). Эксперты спорят об интерпретации этих признаков — некоторые рыбы тоже любят выходить погулять на воздух и оставляют на суше следы: вспомним, например, чудесную рыбку — илистого прыгуна, который отлично приспособился к прогулкам на свежем воздухе. Но даже если окончательно исчезнут сомнения в тетраподном происхождении тех древних следов, это будет лишь указывать на мозаичный характер эволюции на ранних этапах становления тетрапод. Мы тогда убедимся, что в одной из линий рипидистий четвероногое хождение совершенствовалось быстрее, чем в других. Возможно, это происходило в ущерб более сбалансированной морфологии (см. раздел «Отбор на эволюционную перспективность» в главе 4).
Сомики показали, как рыбе пообедать на суше
Хотя в лаборатории невозможно воспроизвести такие длинные последовательности мелких изменений, как «выход рыб на сушу», но все же лабораторные наблюдения могут многое прояснить. Чтобы понять, как рыбы могли питаться на суше, не имея к этому никакого навыка, было предпринято любопытное исследование. В качестве объекта был выбран представитель клариевых сомиков Channallabes аpus, обитающий в болотистых водоемах Африки. Выбор был неслучаен: все клариевые сомики способны передвигаться на суше, а некоторые представители этого семейства переползают из одного озера в другое, двигаясь подобно угрю. Для дыхания они используют специальное «легкое» — видоизмененный участок глотки за жабрами, пронизанный кровеносными сосудами. Могут они дышать и лишенной чешуи кожей. Таким образом, сомики приспособились вести двойную жизнь — в воде и на суше, подобно позднедевонским четвероногим рыбам.
Но могут ли они питаться на суше? Ведь питание у всех рыб построено на едином механизме захвата добычи — они ее всасывают. Раздвигая жабры, они создают в жаберной полости пониженное давление, затем открывают рот — и туда устремляется поток воды с пищевыми объектами, кому какие по нраву. А как быть тем рыбам, которые, подобно клариевому сомику, проголодались, будучи на суше? Ведь на воздухе такой механизм захвата пищи не сработает.
Наблюдения показали, что сомик тем не менее и на суше не голодает. Чтобы схватить добычу, он выбрасывает голову вперед, сбивает насекомое (а наземные беспозвоночные составляют основу его рациона) и, нагибая голову, прижимает добычу к земле и хватает ее. Правда, всосать и проглотить прямо на месте он может лишь мягкую добычу — слизня или червя. Жестких тварей вроде жуков приходится все же тащить в воду.
Размышления о способе наземного питания у современных сомиков привели ученых к выводу, что у рыб, начавших в девоне осваивать сушу, были «преадаптации» к питанию в воздушной среде. На пути «из воды на воздух» животные использовали любые возможности, заложенные в рыбий план строения.
Возможность питаться на суше поначалу могла обеспечиваться гибкой шеей и выработкой специальной позы «голова к земле». Точно так же, между прочим, питается илистый прыгун, обитатель мангровых зарослей, известный любитель свежего воздуха. У тиктаалика и ихтиостеги тоже имелся характерный изгиб шейного отдела позвоночника. Не исключено, что древние наземные позвоночные в первую очередь учились не рот пошире открывать, а, подобно сомикам, изящно изгибать шею. Этот способ захвата добычи был знаком рыбам, так что завоевателям суши не пришлось изобретать на первых порах ничего нового.
Современные экспериментаторы: илистый прыгун и удильщик
Некоторые ныне существующие костистые рыбы уже в кайнозое стали предпринимать новые «попытки» освоить сушу, порой весьма успешные. У этих современных рыб предпосылок для выхода на сушу куда меньше, чем у их девонских предшественников. Бывшие легкие у них уже «истрачены» на плавательный пузырь, а задние конечности (брюшные плавники) пришли в негодное для превращения в ноги состояние. Тем не менее илистый прыгун ловко ходит по суше, опираясь на грудные плавники и помогая себе хвостом, и дышит воздухом при помощи нового, довольно «кустарного» и малоэффективного органа воздушного дыхания, который сформировался у него в ротовой полости. Эти удивительные рыбы способны питаться на суше и даже размножаются на берегу в башенках из глины, на дне которых они устраивают лужицу, где и развивается икра.
Мы не знаем, дадут ли илистые прыгуны начало новой волне сухопутной экспансии рыб. Может, и дадут, хотя сейчас у них на суше многовато конкурентов. У девонских четвероногих рыб конкурентов на берегу не было, зато пища (наземные и почвенные беспозвоночные) была уже изобильна. Если все же от храбрых илистых прыгунов произойдет новая большая сухопутная группа, зоологи будущего придадут ей высокий ранг, а на ее родоночальников — нынешних илистых прыгунов — будут смотреть уже совсем другими глазами: не как на экстравагантного аутсайдера, а как на дальновидного экспериментатора.
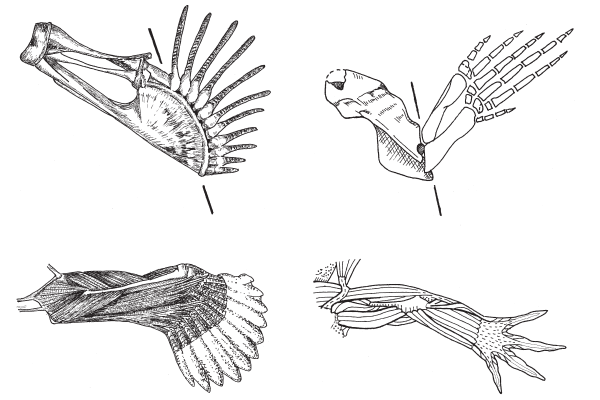
Схема строения передних конечностей мелководного удильщика Antennarius (слева), древней амфибии Tulerpeton (справа вверху) и аксолотля (справа внизу). Линией показано «главное место сгиба»: у тулерпетона это локтевой сустав, у удильщика — нечто совсем другое, но аналогичное по функции. Рисунок А. Н. Кузнецова.
Илистые прыгуны — далеко не единственные рыбы, которые в наши дни в той или иной мере «воспроизводят» то давнее макроэволюционное событие — выход на сушу. Еще один пример — мелководные удильщики антеннарииды. Они не выходят на сушу, однако грудные плавники у них приобрели поразительное сходство с лапами четвероногих. У них есть даже нечто вроде пальчиков!
Удильщики при помощи своих лап ползают по дну и держатся за камни, чтобы их не болтало волнами. Как видим, лапы могут быть полезны и под водой (первые тетраподы тоже, скорее всего, большую часть жизни проводили в воде). Хотя лапы удильщиков внешне и по своим движениям очень похожи на лапы тетрапод, скелет у них другой. Такое сходство называют аналогичным, т. е. появившимся независимо на разной основе, в противовес гомологичному сходству, которое демонстрируют родственные виды и роды.
Динозавры осваивают воздух
Современный мир изобилует летающими существами — насекомые, птицы, летучие мыши; есть и другие, которые хоть и не настоящие летуны, но уже и не совсем наземные жители — древесные лягушки, белки, шерстокрылы, ящерицы — «летучие драконы»... Существует даже аэропланктон — мельчайшие животные и бактерии, приспособленные переноситься на большие расстояния воздушными массами. Человек тоже от них не отстает — поднимается в воздух при помощи технических средств. Таким образом, у многих групп животных прослеживается тенденция к освоению воздушного пространства.
Полет требует согласованных и множественных перестроек тела. Чем они сбалансированнее и глубже, тем искуснее получившийся летун. В этом разделе мы разберем, как полетели птицы, потому что их эволюционная история замечательно прояснилась в последние годы благодаря серии блестящих находок различных переходных форм.
Вскоре после публикации «Происхождения видов» был обнаружен первый скелет археоптерикса — знаменитой промежуточной формы между рептилиями и птицами. Он был захоронен в позднеюрских отложениях Германии, возраст которых, по последним данным, чуть менее 150 млн лет. У археоптерикса было развито оперение (типичная птичья черта), а по строению скелета он мало отличался от мелких динозавров-теропод. У него были когти на передних конечностях, зубы и длинный хвост. Характерных «птичьих» особенностей скелета было немного (крючковидные отростки на ребрах, вилочка). Археоптерикс — классический пример переходной формы. Но это ископаемое было лишь одиноким указателем на длинной дороге, ведущей к птицам. Ясно, что при постепенном становлении птичьих черт подобные промежуточные формы были гораздо разнообразнее. Но вот сохранились ли они? Удастся ли палеонтологам их обнаружить?
Удалось. Дальнейшие поиски в юрских и меловых слоях открыли богатую фауну примитивных переходных полудинозавров-полуптиц. Ниже показана филогенетическая схема, отражающая родственные связи большинства известных на сегодня переходных форм, сочетающих признаки птиц и динозавров.
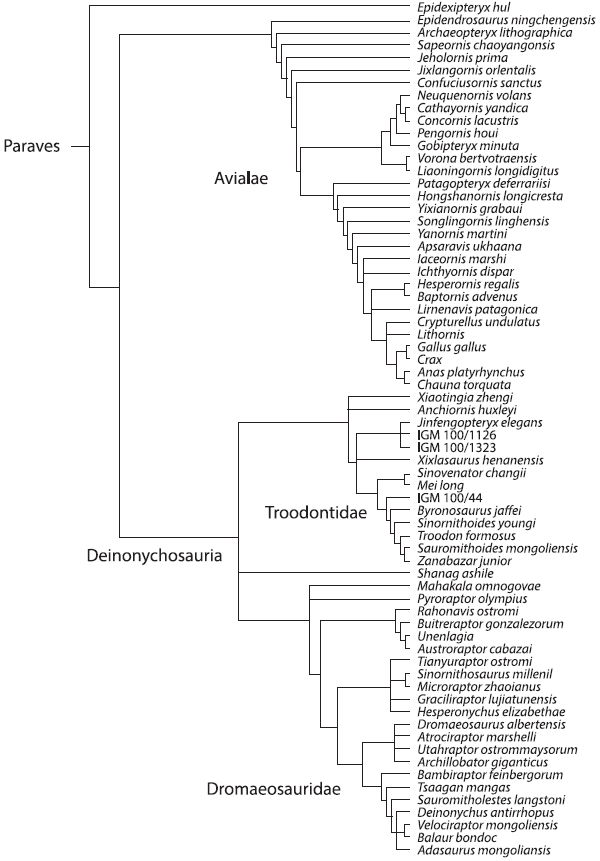
В силу правил современной кладистической систематики факт происхождения птиц от динозавров (а не от общих с динозаврами предков) требует рассматривать птиц как подгруппу динозавров, а для «собственно» динозавров теперь применяют громоздкий термин non-avian dinosaurs (нептичьи динозавры).
Началом становления птиц следует считать юрскую эпоху, когда многие хищные динозавры начали эволюционировать «в птичью сторону». Представителей той линии динозавров, в которой происходило это параллельное «оптичивание», объединяют под общим назанием Paraves. Сами современные птицы на схеме представлены четырьмя видами — курицей (Gallus) и гокко (Crax), уткой (Anas) и паламедеей (Chauna). Они занимают скромное место на одной из веточек среди множества ископаемых видов, формируя вместе с ними обширную группу Avialae, «сестринскую» по отношению к другой большой кладе оперенных динозавров — дейнонихозаврам. Дейнонихозавры подразделяются на троодонтид и дромеозаврид. Археоптерикс занял на схеме вполне логичное место одного из самых примитивных представителей всей разнообразной группы мезозойских летунов. В конце мелового периода, 65,5 млн лет назад, все они вымерли, кроме небольшой группы, которая пережила меловой кризис и дала начало современным птицам.
Самым примитивным представителем Paraves на сегодняшний день считается эпидексиптерикс. Он близок к общему предку первых летающих динозавров. Его нашли в 2008 году в юрских (от 168 до 152 млн лет назад) отложениях Внутренней Монголии (Китай). Это животное размером с голубя. По оценке исследователей, эпидексиптерикс весил примерно 160 г — меньше, чем другие птицеподобные динозавры. От своих родственников он отличался прежде всего коротким хвостом: у него всего 16 хвостовых позвонков, причем последние десять образуют нечто, напоминающее пигостиль примитивных птиц, хотя эти позвонки и не сливаются в единое целое. Длина хвоста составляет 70 % длины туловища, тогда как у ближайшего родственника — эпидендрозавра — хвост был втрое длиннее туловища.
Название «эпидексиптерикс» (от греч. epidexi - «показывать» и pteryx - «перо», «крыло») динозаврик получил неспроста. К хвостовым позвонкам прикреплялись четыре необычайно длинных пера с центральным стержнем и опахалом из неветвящихся бородок. Туловище, включая конечности, покрывали мелкие перья без стержня, похожие на пух других динозавров и примитивных птиц. Перьев, пригодных для полета, у эпидексиптерикса не было, и летать он наверняка не мог.
Эти длинные хвостовые перья служили исключительно «для красоты», т. е. для привлечения брачного партнера. Самцы многих современных птиц красуются перед самками длинными хвостовыми перьями, больше ни на что не годными и только мешающими летать. Наряд эпидексптерикса доказывает, что красота брачных нарядов была важна для «птичьей» ветви динозавров уже в самом начале их истории (Zhang et al., 2008).
Перья нужны для красоты, тепла и полета. По-видимому, сначала они выполняли две первые задачи и только позже пригодились для третьей. Да и полет начинался не сразу: маховые перья развивались постепенно. Увеличение несущей поверхности передних конечностей повышало стабильность бегущего или прыгающего динозаврика. Начальные этапы эволюции маховых перьев, возможно, были связаны со стабилизацией быстрого бега по неровной поверхности. При этом большие перья постепенно смещались на задний край конечности; строение самих перьев и мест их крепления постепенно совершенствовалось.
Оперенные примитивные предки птиц дали начало группе дромеозаврид. Эти животные распрощались с поверхностью земли, предпочитая деревья. Недавние исследования показали, что даже прославленный велоцираптор не столько бегал по равнинам, сколько лазил по деревьям, и его строение было приспособлено именно к этому образу жизни. Выяснилось, что велоцирапторы были покрыты перьями, что подразумевает теплокровность (Turner et al., 2007). Раньше их считали (эта версия увековечена фильмом «Парк юрского периода) скоростными равнинными стайными хищниками, наделенными страшным оружием — зубастыми челюстями и четырьмя гигантскими когтями (по одному на каждой из четырех конечностей). Но оказалось, что устройство и геометрия когтей на задних лапах велоцираптора сравнимы с когтями некоторых лазающих по деревьям птиц и млекопитающих. Это сравнение со всей определенностью переместило велоцираптора с земли на дерево. Велоцирапторы использовали колоссальные когти на ногах не только для того, чтобы убивать толстокожую жертву. При лазании по стволу они опирались на этот коготь. Он должен был быть крупным, чтобы удерживать на вертикальном стволе довольно массивное животное — эти динозавры весили около 15 кг.
Скорее всего, у велоцирапторов был «птичий» механизм фиксации когтей. Когда птица сидит на ветке, ее когти обхватывают ветку и фиксируются в этом положении без затрат энергии. Это позволяет птице спать сидя на дереве и при этом не падать. Опорой для древесного хищника при посадке на стволе также служил жесткий хвост, составленный сросшимися позвонками и окостеневшими сухожилиями. Перья, древесный образ жизни, да еще и в стае — ну чем не птичка!
Анхиорнис (Anchiornis huxleyi) принадлежит к другой части филогенетического дерева — к базальным (т. е. примитивным, ранним) троодонтидам. Анхиорнис жил раньше археоптерикса — 155–151 млн лет назад. Когда его нашли и описали (а это произошло в 2009 году), он сразу занял почетное место среди самых удивительных и неожиданных животных. Это летающее создание величиной с ворону имело контурные маховые перья и на передних, и на задних конечностях. Превосходные отпечатки безошибочно передают строение скелета и перьев, так что в интерпретациях нет сомнений — на ногах у анхиорниса были крупные перья, пригодные для полета.
Это не первый найденный четырехкрылый динозавр — такие были известны и раньше, например микрораптор, живший 125 млн лет назад. Поначалу многие эксперты трактовали четырехкрылость как некий курьез, экстравагантный, но незначительный поворот эволюции Paraves.
В 2005 году был описан еще один динозавр с крупными перьями на ногах — педопенна (что переводится как «пероног»), жившая одновременно с археоптериксом или, возможно, чуть раньше. Она известна только по остаткам оперенных задних конечностей. Педопенну интерпретировали как базального представителя Paraves (на схеме она не показана)
Судя по распределению четырехкрылых форм по эволюционному дереву, четырехкрылость была свойственна первым представителям всех трех ветвей Paraves: троодонтидам, дромеозавридам и Avialae. Из этого следует, что современные птицы произошли от четырехкрылых предков. Между прочим, у примитивных Avialae, включая археоптерикса, действительно были на ногах контурные перья, хотя и не такие крупные.
В отличие от тех современных птиц, у которых имеется более или менее развитое оперение на ногах, у микрораптора, педопенны и анхиорниса контурные перья задних конечностей располагались упорядоченно и образовывали большие плоские поверхности, которые, несомненно, влияли на аэродинамические свойства животного. Как именно использовались эти дополнительные плоскости — для планирования, маневрирования или, допустим, торможения, — точно не известно. Едва ли четырехкрылые динозавры могли активно махать своими «задними крыльями» — скелет их задних конечностей не приспособлен для таких движений, — но они могли разворачивать ноги таким образом, чтобы перья помогали планировать. Четырехкрылый полет в любом случае не слишком эффективен.
Анхиорнис, судя по скелету, был не очень хорошим летуном (микрораптор летал получше). Задние конечности анхиорниса, если забыть о перьях, больше похожи на ноги профессионального бегуна. Но крупные перья на ногах едва ли помогали ему быстро бегать — скорее, наоборот, должны были мешать. У современных быстро бегающих птиц прослеживается тенденция к утрате оперения на ногах. Может быть, анхиорнис еще не успел «решить», что для него важнее — летать или бегать, и строение его задних ног является результатом компромисса между разнонаправленными векторами отбора. В ходе дальнейшей эволюции ранних птиц крылья и ноги более четко поделили функции: первые для полета, вторые для бега.
Анхиорнис по многим чертам скелета похож на троодонтид, к которым его и отнесли, но некоторые признаки (например, очень длинные передние конечности) сближают его с дромеозавридами и Avialae. В результате формальные диагнозы (списки отличительных признаков) трех групп Paraves становятся более расплывчатыми. Это типичная ситуация, неприятная для систематиков, но абсолютно неизбежная при изучении базальных представителей крупных таксонов (об этом мы говорили при обсуждении кембрийских типов). Понятно, что чем ближе мы подбираемся к общему предку всех Paraves, тем менее четкими становятся различия между тремя эволюционными линиями, входящими в состав этой группы. «Переходные формы» размывают эти различия. Дарвин был прав, рассуждая о стирании границ между таксономическими группировками.
Кроме того, мозаика птичьих и динозавровых черт, столь широко представленная у Paraves, еще раз напоминает о параллельной эволюции. Начало эволюции каждой крупной группы, как правило, представлено несколькими линиями, в каждой из которых своим чередом, со своими особенностями и со своей скоростью шло приспособление к новой среде или новому образу жизни. В случае «оптичивания» динозавров это линии дромеозаврид, троодонтид и Avialae.
Птицы, динозавры и их гены
Признать, что птицы являются динозаврами — а именно этого требует современная систематика, — психологически довольно трудно. К тому же есть и «нединозавровые» гипотезы происхождения птиц. Правда, к сегодняшнему дню у них почти не осталось сторонников (об одной из них мы рассказали в книге «Рождение сложности»). Мог бы помочь генетический анализ, но ДНК динозавров не сохранилась (по крайней мере, пока не выделена и не прочитана). Но все же подобраться к генам динозавров ученые смогли (Organ et al., 2009). Они косвенным образом оценили размер геномов различных динозавров. Для этого они воспользовались тем обстоятельством, что в ископаемых костях, если они достаточно хорошо сохранились, на срезах бывают видны маленькие полости, в которых при жизни животного помещались клетки костной ткани — остеоциты.
Известно, что размер генома во многих группах живых существ положительно коррелирует с размером клеток. Срезы костей 26 видов современных четвероногих, взятых для исследования, обнаружили линейную зависимость между размером генома и средним объемом остеоцита. Найденная зависимость позволила ученым с приемлемой точностью оценить размеры геномов 31 вида динозавров и ископаемых птиц.
У современных птиц размер генома необычно мал — от 0,97 до 2,16 млрд пар нуклеотидов, в среднем 1,45. Для сравнения, у жабы 6,00, крокодила 3,21, коровы 3,7, кошки 2,9, мыши 3,3, человека 3,5. А что же динозавры? Геномы птицетазовых динозавров (вопреки своему названию не родственных птичьей ветви), имели средний размер около 2,5 млрд пар нуклеотидов, что сопоставимо с современными рептилиями. Геномы теропод (хищных двуногих динозавров), в том числе самых древних, живших задолго до появления птиц, были значительно меньше — в среднем 1,78 млрд п. н. Из девяти исследованных видов теропод только у одного (овираптора) размер генома оказался за пределами диапазона, характерного для современных птиц.
Таким образом, общий предок всех динозавров, скорее всего, имел типичный для наземных позвоночных большой геном. Это состояние сохранилось у птицетазовых динозавров, а также у рептилий, доживших до наших дней. У ящеротазовых динозавров (к которым относятся тероподы) еще на заре их истории (в триасе) произошло радикальное сокращение генома. Птицы, таким образом, унаследовали маленький геном от своих предков — динозавров-теропод, а не приобрели его позже как адаптацию к полету.
Тем не менее между размером генома и полетом все-таки есть связь. Об этом свидетельствуют два обстоятельства. Во-первых, у нелетающих птиц (таких как страусы) геном больше, чем у летающих. Во-вторых, у летучих мышей геном меньше, чем у других млекопитающих.
Еще одно генетическое исследование, пролившее свет на раннюю эволюцию птиц, было выполнено в 2011 году.
С точки зрения эмбриологии и сравнительной анатомии три пальца крыла у птиц соответствуют пальцам II, III и IV исходной пятипалой конечности. Это противоречит палеонтологическим данным: ископаемые находки демонстрируют постепенную утрату пальцев IV и V в эволюционном ряду, ведущем от первых триасовых динозавров к археоптериксу и другим вымершим ящерохвостым птицам. Три оставшихся у них пальца — это пальцы I, II и III. Значит, если современные (веерохвостые) птицы являются потомками ящерохвостых, то пальцы их крыльев должны быть пальцами I-II-III, а не II-III-IV. Получается, пальцы — всего лишь пальцы! — не соглашаются происходить от динозавров. И этого достаточно, чтобы заронить сомнения в справедливости динозавровой теории. Но стараниями генетиков этот конфликт между эмбриологией и палеонтологией был разрешен (Wang et al., 2011).
Зачатки крыльев (слева) и ног (справа) цыпленка на ранней (вверху, E) и более поздней (внизу, L) стадиях развития. Длина масштабной линейки — 1 мм. Пальцы нумеруются слева направо. Видно, что по своему положению в зачатках конечностей три пальца крыла (Fa, Fb, Fc) соответствуют пальцам ноги II, III и IV (Hb, Hc и Hd). Однако набор генов, работающих в зачатке первого пальца крыла (Fa), соответствует первому пальцу ноги (Ha), а не второму (Hb). Из Wang et al., 2011.
Они измерили активность генов в зачатках пальцев передних и задних конечностей куриного эмбриона. Всего в зачатках пальцев удалось зарегистрировать активность 14 692 генов. Анализ проводился по отдельности для каждого пальца на двух стадиях развития зачатков (ранней и поздней). Выяснилось, что по характеру экспрессии генов первые (самые внутренние) пальцы крыльев и ног четко отличаются от остальных (внешних) пальцев, но при этом они похожи друг на друга. Первые пальцы отличаются от других по уровню экспрессии 556 генов. Это весомый аргумент в пользу того, что первый палец крыла — это на самом деле палец I, а не II, как следует из данных эмбриологии.
Редукция пальцев IV и V в эволюционном ряду от ранних динозавров к птицам. Пальцы нумеруются слева направо. (A–C) триасовые динозавры: (A) Heterodontosaurus (птицетазовый динозавр, не родственный птицам), (B) Herrerasaurus (ранний представитель теропод), (C) Coelophysis (более продвинутый представитель теропод). (D–E) юрские тероподы: (D) Allosaurus, (E) Ornitholestes. (F–G) ящерохвостые птицы: (F) Archaeopteryx, (G) меловой Sinornis. (H–I) современные (веерохвостые) птицы: (H) птенец гоацина (Opisthocomus), (I) курица (Gallus). Из Vargas A. O., 2005. Beyond Selection // Revista Chilena de Historia Natural. V. 78. P. 739–752.
Очевидно, именно работой этих 556 генов определяется «идентичность» первых пальцев с молекулярно-генетической точки зрения. Что касается второго и третьего пальцев крыла, то их программы развития, похоже, сформировались заново из фрагментов программ развития пальцев II, III и IV. Поэтому нельзя определенно сказать, каким пальцам ноги они гомологичны с молекулярно-генетической точки зрения. Но если учесть весь комплекс имеющихся данных (молекулярно-генетическую гомологию первых пальцев, взаимное расположение пальцев крыла, палеонтологические данные), то все-таки получается, что три пальца птичьего крыла происходят от пальцев I-II-III, а не II-III-IV. А это значит, что конфликт между данными эмбриологии и палеонтологии снят.
Полученные результаты подтвердили высказанную ранее гипотезу, согласно которой в эволюции динозавров-теропод имело место смещение областей работы программ развития пальцев I-II-III на одну эмбриональную позицию в дистальном направлении (от туловища), так что палец I стал развиваться там, где положено развиваться пальцу II (Wagner, Gauthier, 1999).


«Ничего особенно не трудно, если разделить работу на части»
Этим известным высказыванием мы обязаны Генри Форду. Руководствуясь им, он добился успеха. Эволюция, следуя этому правилу, не раз проделывала многотрудную работу. И тоже достигла колоссальных успехов. Мы видим совершенные конструкции идеальных летунов — буревестников, стрижей; мы видим превосходно сконструированных стремительных бегунов — лошадей, гепардов; мы знаем орлов, обладающих почти волшебной остротой зрения; удивляемся точности шарнирных креплений у роботоподобных насекомых; превозносим мозговитого супермыслителя — человека. У них у всех есть сложные органы, прекрасно подходящие для своего назначения: совершенные ноги для бега, высокотехничные суставы, эффективные мышцы, глаза и мозг. Будь они чуть по-другому устроены, сбей немного настройку их частей — и нога поплетется, коленка не согнется, глаз не увидит, а крыло не полетит. Как естественный отбор справлялся с созданием органов, которые, казалось бы, полезны только в полностью собранном виде? Правильный ответ — постепенно. И на каждом шаге этого процесса конструкция оказывалась так или иначе полезной своему обладателю. Это могло быть последовательное усовершенствование структуры, в ходе которого нужный орган все лучше служил своему хозяину. Но бывало и по-другому: «недоделанный» орган использовался совсем не так, как его применяет сегодняшний счастливый обладатель.
Рассмотрим один яркий пример. Путешествие косточек первой и второй жаберных дуг — характерных для всех челюстноротых рыб — внутрь черепа и превращение их в слуховые косточки похоже на фокус. Мы видим у примитивных рыб несколько составных жаберных (висцеральных) дуг, среди которых выделяется первая — у челюстноротых она стала верхней и нижней челюстями. Между верхней и нижней челюстью имеется челюстной сустав. Остальные жаберные дуги более или менее одинаковые. Затем обретает собственную особую роль вторая жаберная дуга8. Соединенная с черепной коробкой, она служит подвеской для жаберной крышки. Функции первых двух дуг у рыб очевидны: первая — это челюсти, вторая — дыхательный насос. Затем с переходом к воздушному дыханию надобность в жаберной крышке исчезает, и она редуцируется. А косточка сустава, при помощи которой жаберная крышка крепилась к черепу, отправляется в ссылку в среднее ухо передавать звуковые сигналы. Так появилась первая слуховая косточка — стремечко. И произошло это, как мы помним, еще у девонских «четвероногих рыб». И сама косточка, и ее сустав пригодились четвероногим рыбам в новом качестве.
Первая жаберная дуга — рыбьи челюсти — с переходом к наземной жизни тоже преобразуется. Ее окостеневшая часть в виде челюстного сустава еще кое-как служила на своем месте, но потом у млекопитающих тоже осталась не у дел — сустав обновился за счет других костей. Этот отслуживший сустав бывшей первой дуги получил новое назначение — он тоже отправился в среднее ухо. В среднем ухе, таким образом, собралась компания из трех косточек, некогда кормивших своих хозяев и помогавших им дышать, но смещенных со своих позиций более пригодными в новых условиях элементами и сочленениями. Эти три косточки — молоточек, наковальня и стремечко — при этом не исчезли, а взяли на себя новые обязанности по восприятию и передаче звуковых колебаний. Более того, они стали одним из основных признаков скелета, по которому можно отличить млекопитающих от всех других наземных тетрапод. Вот такой эволюционный маршрут у слуховых косточек: от жаберных дуг к среднему уху. Если не знать промежуточных этапов этого маршрута, то кажется, что подобный путь невероятен и невозможен. К счастью, чудесные трансформации висцеральных дуг удается проследить как по данным эмбриологии и сравнительной анатомии, так и на ископаемом материале.
Ископаемые примитивные млекопитающие демонстрируют переходы от рептильной организации с одной слуховой косточкой (стремечком) к формам, у которых имеются все три, но две из них (молоточек и наковальня) еще остаются приросшими к нижней челюсти. Это память о прошлом, когда они работали в качестве челюстного сустава. Отделение молоточка и наковальни от челюсти тоже происходило поэтапно. Об этом можно было догадываться, основываясь на данных эмбриологии. У современных млекопитающих в ходе эмбрионального развития сначала слуховые косточки отделяются от боковой поверхности нижней челюсти, но остаются связаны с ней спереди посредством меккелева хряща — остатка первичной нижней челюсти. На втором этапе исчезает и это переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается. И вот нашлись такие ископаемые виды, у которых во взрослом состоянии сохраняется частичное соединение слуховых косточек с нижней челюстью. Морганукодон (Morganucodon) — примитивный представитель «маммалиаформ» (непосредственных предков млекопитающих), живший в позднем триасе около 205 млн лет назад. У морганукодона в среднем ухе уже есть все три косточки. Но молоточек и наковальня все же соединены с нижней челюстью и спереди, и с боков. А недавно были найдены еще два древних животных, у которых процесс отделения слуховых косточек продвинулся чуть дальше. Это яноконодон (Yanoconodon allini) и ляоконодон (Liaoconodon hui), примитивные млекопитающие, жившее 125-120 млн лет назад (в раннемеловую эпоху) (Luo et al., 2007; Meng et al., 2011). От них остались скелеты превосходной сохранности, в которых сохранились и кости среднего уха.
Яноконодон и ляоконодон, как и большинство мезозойских млекопитающих, были мелкими, невзрачными ночными зверьками. Как и любым ночным животным, им весьма пригодился бы тонкий слух. Скорее всего, ночной образ жизни первых млекопитающих способствовал эволюции органов слуха. У яноконодона и ляоконодона слуховые косточки демонстрируют ту стадию, когда молоточек и наковальня уже отделились от нижней челюсти с боков, но еще прикреплены к ней спереди посредством меккелева хряща — древней рыбьей челюсти. Совсем как на одной из эмбриональных стадий развития среднего уха у современных млекопитающих. Эти ископаемые звери стали настоящим подарком эволюционистам — яркой иллюстрацией постепенного становления сложных органов.
Нужно, однако, иметь в виду, что три слуховые косточки сами по себе еще не означают радикального улучшения слуха. Для того чтобы дальше совершенствовать эту функцию, нужно к механизму передачи звуковых колебаний подтянуть и способы обработки звуковой информации, а это тоже произошло не сразу. Например, у одного из примитивных млекопитающих Hadrocodium молоточек и наковальня уже отделены от нижней челюсти. Но строение внутреннего уха у Hadrocodium еще примитивное, такое же, как у Morganucodon. По-видимому, древние млекопитающие еще не были первоклассными слухачами. Слух совершенствовался постепенно по мере развития разных элементов системы восприятия, передачи и анализа звуковой информации.
Сначала чутье, потом философия
Мозг млекопитающих (и особенно человека) нередко сравнивают со сверхсложным компьютером, способным выполнять сразу множество задач. Создать подобный компьютер и довести его до такого совершенства, чтобы он работал, не «повисая» каждую минуту, и верно служил своему обладателю всю его жизнь... ну как такое сделать! Однако природа все же справилась с этим, постепенно наращивая мощности и координируя когнитивные задачи. Начав с нервной трубки первых хордовых, естественный отбор постепенно добрался до млекопитающих с крупным мозгом и развитым неокортексом (новой корой).
Мозг млекопитающих радикально отличается от мозга рептилий не только своим размером, но и строением. В частности, у млекопитающих резко увеличились обонятельные луковицы и отделы коры, связанные с обонянием, а также мозжечок.
Особенно нас занимает процесс формирования неокортекса, позволяющего нам, читателям, разбирать и воспринимать буковки, складывать их в слова, анализировать информацию, а затем, запоминая ее, участвовать в дальнейших дебатах. Так как же мы, млекопитающие, получили этот изумительно сложный орган — неокортекс, кору больших полушарий?
Непосредственных предков первых млекопитающих относят к группе маммалиаформ, которые, в свою очередь, берут начало от цинодонтов. Палеонтологи детально реконструировали этапы эволюционных преобразований зубов и скелета цинодонтов по мере их «маммализации» — постепенного эволюционного движения в сторону млекопитающих. Об эволюции мозга известно значительно меньше. Между тем очевидно, что именно развитие мозга во многом предопределило эволюционный успех млекопитающих.
Изучение мозга маммалиаформ и первых млекопитающих затруднялось, во-первых, тем, что редко удавалось найти хорошо сохранившийся череп, во-вторых — тем, что для изучения эндокрана (слепка мозговой полости, по которому можно судить о размере и форме мозга) череп, как правило, нужно было разрушить.
Американским палеонтологам удалось в некоторой мере восполнить этот досадный пробел (Rowe et al., 2011). При помощи компьютерной рентгеновской томографии, не разрушая драгоценных черепов, они получили детальные трехмерные изображения эндокранов двух маммалиаформ, живших в начале ранней юры (200-190 млн лет назад) на территории нынешнего Китая.
Изученные маммалиаформы Morganucodon oehleri (уже упоминавшееся животное с готовыми косточками среднего уха) и Hadrocodium wui - ближайшие родственники первых млекопитающих. По строению скелета они представляют собой классические переходные формы между «еще рептилиями» и «уже млекопитающими». При этом Morganucodon стоит ближе к примитивным цинодонтам, а Hadrocodium настолько близко подошел к млекопитающим, насколько это возможно, оставаясь формально за пределами группы. Исследование показало, что по строению мозга эти животные тоже занимают промежуточное положение между типичными цинодонтами и их потомками — млекопитающими.
Ранее были изучены эндокраны ранних цинодонтов Thrinaxodon и Diademodon. Выяснилось, что мозг у них был еще вполне рептильный по своему размеру и строению.
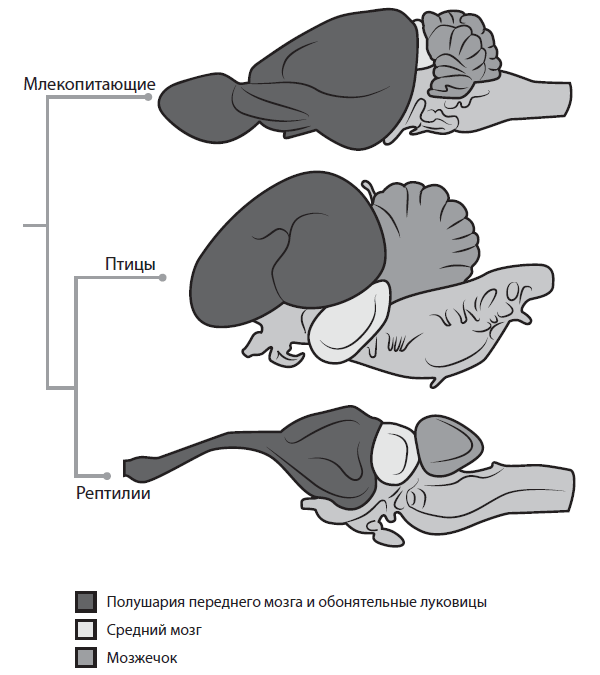
Соотношение отделов мозга у млекопитающих, птиц и рептилий.
Относительный размер мозга у позвоночных оценивается при помощи «коэффициента энцефализации» (EQ), который вычисляют по эмпирически выведенной формуле EQ = EV / (0,055[Wt]0,74), где EV — объем мозговой полости в миллилитрах, Wt — масса тела в граммах.

У базальных цинодонтов Thrinaxodon и Diademodon EQ варьирует от 0,16 до 0,23. Для сравнения, у крысы EQ = 0,53. Обонятельные луковицы у них были небольшие, а в носу не было окостеневших носовых раковин, что свидетельствует о слабом развитии обонятельного эпителия. Передний мозг был маленький и узкий, не подразделенный на отделы, без признаков наличия неокортекса. Средний мозг и эпифиз (теменной глаз) не были закрыты сверху полушариями переднего мозга. Мозжечок был шире переднего мозга, спинной мозг тонкий. Эти и другие «рептильные» особенности мозга и черепа цинодонтов свидетельствуют о том, что у них по сравнению с млекопитающими было слабое обоняние и не слишком совершенный слух, осязание и координация движений.
Мозг Morganucodon, как выяснилось, был больше похож на мозг млекопитающего. По объему он в полтора раза превосходит мозг базальных цинодонтов (EQ = 0,32). Сильнее всего увеличилась обонятельная луковица и обонятельная кора. Это со всей очевидностью указывает на более развитое обоняние. Полушария переднего мозга стали выпуклыми благодаря развитию неокортекса. Они закрывают сверху средний мозг и эпифиз, как у млекопитающих. Передний мозг морганукодона шире мозжечка, хотя мозжечок тоже заметно вырос по сравнению с базальными цинодонтами. Увеличение мозжечка говорит об улучшенной координации движений. На это же указывает и более толстый, чем у базальных цинодонтов, спинной мозг.
Развитие неокортекса у древних млекопитающих было связано в первую очередь с совершенствованием соматосенсорных функций. Значительная часть неокортекса у примитивных млекопитающих, таких как опоссум, — это так называемая соматосенсорная кора, которая отвечает за сбор и анализ сигналов, приходящих от многочисленных механорецепторов, рассеянных по всему телу. Особенно много таких рецепторов приурочено к волосяным фолликулам.
Базальный цинодонт Thrinaxodon (Южная Африка, Антарктика, ранний триас, 248-245 млн лет назад).
По мнению многих палеонтологов, волосы выполняли сначала тактильную (осязательную) функцию, а для терморегуляции стали использоваться позже, когда у предков млекопитающих начала развиваться теплокровность. У Morganucodon и Hadrocodium достоверных остатков волосяного покрова пока не обнаружено, но их близкий родственник — похожий на бобра маммалиаформ Castorocauda (в переводе с латыни — «с хвостом как у бобра») — был покрыт густым мехом, состоявшим из осевых волос и подшерстка. Это позволяет предположить, что Morganucodon и Hadrocodium тоже были покрыты шерстью. Вероятно, появление неокортекса у маммалиаформ было связано в том числе и с развитием волосяного покрова и осязания.
Таким образом, мозг примитивного Morganucodon иллюстрирует первый этап прогрессивной эволюции мозга в ходе становления млекопитающих. На этом этапе увеличение мозга было обусловлено развитием обоняния, осязания и координации движений.
Hadrocodium, более продвинутый представитель маммалиаформ и ближайший родственник «настоящих» млекопитающих, иллюстрирует второй этап развития мозга. Коэффициент энцефализации у Hadrocodium равен 0,5, т. е. Мозг увеличился еще в полтора раза по сравнению с Morganucodon и достиг размеров, характерных для некоторых настоящих млекопитающих. Мозг вырос в основном за счет обонятельных луковиц и обонятельной коры. Таким образом, второй этап прогрессивной эволюции мозга в ходе маммализации тоже был связан с развитием обоняния.
Третий этап соответствует переходу от высших маммалиаформ, таких как Hadrocodium, к настоящим млекопитающим. На этом этапе обоняние становится еще более тонким, о чем свидетельствуют изменения решетчатой кости: на ней образуются носовые раковины, поддерживающие разросшийся обонятельный эпителий.
Эти данные показывают, что потребность в тонком чутье была, по-видимому, главным стимулом развития мозга в ходе становления млекопитающих. У млекопитающих, как известно, обоняние развито намного лучше, чем у всех остальных наземных позвоночных. Скорее всего, изначально это было связано с приспособлением к ночному образу жизни, когда на зрение полагаться бессмысленно (мы говорили об этом в главе 5). В такой ситуации было выгодно совершенствовать уже имеющуюся древнюю систему обоняния: разнообразить обонятельные рецепторы и усиливать мозговые мощности для их анализа. Что ж, пусть это было обоняние, зато в качестве побочного продукта млекопитающие получили кору с массой нейронов и в конце концов стали использовать ее для осмысления произошедших перемен.
1 Эти и другие примеры радикальной смены функций мы рассмотрим в главе 5.
2 Об РНК-организмах и теории РНК-мира рассказано в книге «Рождение сложности».
3 Не вдаваясь в сложные вычисления, заметим лишь, что число поколений, которое (в среднем) должно пройти до фиксации или элиминации нейтральной мутации, имеет примерно тот же порядок величин, что и численность популяции. Если в популяции несколько тысяч особей, то и ждать придется несколько тысяч поколений, в миллионной популяции речь будет идти о миллионах поколений. Поэтому на судьбу нейтральных мутаций сильное влияние оказывают «бутылочные горлышки» — периоды сокращения численности популяции, во время которых нейтральный аллель может потеряться или зафиксироваться особенно быстро.
4 Потому что разные участки ландшафта приспособленности имеют разную «проходимость»: одни похожи на ровные плато (там нейтральная эволюция идет быстро), другие — на лабиринт узких тропинок над пропастями (там нейтральные изменения накапливаются медленно). Подробнее об этом см. ниже.
5 Ответ такой: \( q = \frac{F^x×k}{1+F^xk} \), где q — частота аллеля А2 по прошествии X поколений, F — относительная приспособленность аллеля А2 по сравнению с конкурирующим аллелем А1 (в нашем случае F = 21/20 = 1,05), \( k=\frac{q_{0}}{1-q_{0}} \), где q0 — частота аллеля А2 в начальный момент времени. В бесконечно большой популяции динамика q должна в точности соответствовать этой формуле. В маленьких популяциях соответствие будет неточным из-за генетического дрейфа.
6 Медленность элиминации слабовредных мутаций может пойти на пользу популяции. Ведь некоторые мутации, вредные здесь и сейчас, могут оказаться полезными в перспективе. Главное, чтобы это потенциальное преимущество успело реализоваться до того, как отбор элиминирует мутацию. Мы рассмотрим такие коллизии в следующих главах.
7 Это соответствует известному «биогенетическому закону» Эрнста Геккеля, который гласит, что индивидуальное развитие повторяет эволюцию. В своей жесткой и абсолютизированной форме биогенетический закон неверен (в биологии вообще опасно что-либо абсолютизировать), но в смягченном варианте он вполне справедлив.
8 Иногда эти дуги называют второй и третьей соответственно, так как самая первая дуга — губная — у современных рыб отсутствует или остается в виде рудимента.


Кембрийский взрыв
-
 28.11.2025После смерти трихоплакс мгновенно распадается на клеткиЕлена Наймарк • Новости науки
28.11.2025После смерти трихоплакс мгновенно распадается на клеткиЕлена Наймарк • Новости науки -
 22.05.2017«Летающие жирафы, мамонты-блондины, карликовые коровы...». Глава из книгиАндрей Журавлёв • Книжный клуб • Главы
22.05.2017«Летающие жирафы, мамонты-блондины, карликовые коровы...». Глава из книгиАндрей Журавлёв • Книжный клуб • Главы
-
 03.05.2017Кембрийский взрывСергей Ястребов • Библиотека • «Химия и жизнь» №10, 2016
03.05.2017Кембрийский взрывСергей Ястребов • Библиотека • «Химия и жизнь» №10, 2016
-
 21.11.2015Путь к разгадке Кембрийского взрываИнтервью с Еленой Наймарк • Библиотека • «Троицкий вариант» №23(192), 2015
21.11.2015Путь к разгадке Кембрийского взрываИнтервью с Еленой Наймарк • Библиотека • «Троицкий вариант» №23(192), 2015
-
 13.02.2015Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрываСергей Ястребов • Новости науки
13.02.2015Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрываСергей Ястребов • Новости науки
-
 18.06.2014«Эволюция. Классические идеи в свете новых открытий». Главы из книгиАлександр Марков, Елена Наймарк • Книжный клуб • Главы
18.06.2014«Эволюция. Классические идеи в свете новых открытий». Главы из книгиАлександр Марков, Елена Наймарк • Книжный клуб • Главы
-
 04.06.2014Китайские палеонтологи нашли древнейшего головохоботного червяСергей Ястребов • Новости науки
04.06.2014Китайские палеонтологи нашли древнейшего головохоботного червяСергей Ястребов • Новости науки
-
 17.11.2013Членистоногие подтверждают реальность кембрийского взрываСергей Ястребов • Новости науки
17.11.2013Членистоногие подтверждают реальность кембрийского взрываСергей Ястребов • Новости науки
-
 13.12.2011Диверсификация животных началась задолго до кембрийского взрываАлександр Марков • Новости науки
13.12.2011Диверсификация животных началась задолго до кембрийского взрываАлександр Марков • Новости науки
-
 09.09.2011Тайная мягкая жизньМихаил Федонкин • Библиотека • «Популярная механика» №7, 2011
09.09.2011Тайная мягкая жизньМихаил Федонкин • Библиотека • «Популярная механика» №7, 2011








