«В поисках памяти». Главы из книги

25. О мышах, людях и психических заболеваниях
Подобно тому как в девяностые наши исследования эксплицитной памяти вновь привлекли мое внимание к проблемам, которые еще в колледже привели к увлечению сихоанализом, в начале нового тысячелетия возможность исследовать связанные с возрастом нарушения памяти у мышей неудержимо влекла к вопросам, которые интересовали меня во время резидентуры по психиатрии. Этот возобновившийся интерес к психическим заболеваниям был вызван несколькими факторами.
Во-первых, биологические исследования памяти, которыми я занимался, достигли того этапа, когда я мог обратиться к проблемам, связанным со сложными формами памяти и с ролью избирательного внимания в ее работе, и это побуждало меня к поиску новых модельных объектов для изучения психических заболеваний. Меня также привлекало открытие того, что некоторые формы психических заболеваний, в частности посттравматические стрессовые расстройства, шизофрения и депрессия, сопровождаются той или иной разновидностью нарушений памяти. Теперь, когда я глубже разобрался в молекулярной биологии памяти и убедился, как много могут сказать о возрастных проблемах с памятью такие модельные объекты, как мыши, можно было задуматься и о роли нарушений памяти в других психических заболеваниях и даже в биологических основах психического здоровья.
Во-вторых, за годы моей научной карьеры психиатрия основательно сблизилась с биологией. В шестидесятые, когда я проходил резидентуру в Массачусетском центре психического здоровья, большинство психиатров считало, что социальные детерминанты поведения совершенно независимы от биологических и что и те и другие действуют на разные стороны психики. На основании предполагаемых различий в их происхождении психические заболевания разделяли на две основные группы — органические и функциональные. Такое разделение, впервые предложенное еще вXIX веке, опиралось на результаты вскрытия трупов психически больных людей.
Доступные в то время методы патологоанатомических исследований мозга еще не позволяли выявлять изменения в его тонком строении. Поэтому органическими, то есть вызываемыми биологическими причинами, считались лишь те психические заболевания, которые были сопряжены с ощутимым разрушением нервных клеток и тканей мозга, например болезнь Альцгеймера, синдром Хантингтона и хронический алкоголизм. Шизофрения, различные формы депрессии и неврозы тревоги не провоцировали ни разрушения нервных клеток, ни каких‑либо других явных изменений в строении мозга, поэтому считались функциональными, то есть не вызываемыми биологическими причинами. Функциональные психические заболевания нередко воспринимались обществом как особое клеймо, потому что считалось, что они существуют «лишь в мыслях пациента». Этому представлению сопутствовало предположение, что заболевание могло проникнуть в мысли пациента по вине родителей.
Теперь мы больше не считаем, что лишь некоторые психические заболевания влияют на психику за счет биологических изменений, происходящих в мозгу. Более того, в основе всей новой науки о психике лежит принцип, согласно которому все психические процессы имеют биологическую природу: все они обеспечиваются химическими реакциями и клеточными механизмами, которые в прямом смысле происходят «у нас в голове». Поэтому любое нарушение или изменение работы этих механизмов также должно иметь биологическую основу.
Наконец, в 2001 году мне предложили написать статью о вкладе молекулярно-биологических исследований в неврологию и психиатрию для Journal of the American Medical Association в соавторстве с Максом Кауаном, моим давним другом, вице-президентом и старшим научным сотрудником Медицинского института Говарда Хьюза. Пока я работал над этой обзорной статьей, я поразился, как радикально методы молекулярной генетики и использование животных в качестве модельных объектов преобразовали неврологию. Это заставило меня задуматься, почему молекулярная биология не оказала подобного преобразующего влияния и на психиатрию.
Основная причина состоит в том, что между неврологическими и психическими заболеваниями есть несколько важных отличий. Неврология уже давно строится на знании того, какие именно области нервной системы поражает каждое заболевание. Болезни, которыми занимается неврология (инсульты, опухоли мозга и нейродегенеративные расстройства), сопровождаются вполне заметными структурными повреждениями. Исследование этих болезней научило нас, что главное для нейробиологии — местоположение. Мы уже почти сто лет знаем, что синдром Хантингтона — это расстройство хвостатого ядра, болезнь Паркинсона — черного вещества, а боковой амиотрофический склероз (БАС) — мотонейронов. Мы уверены, что каждая из этих болезней вызывает характерные нарушения движений, потому что все они затрагивают разные компоненты моторной системы.
Кроме того, выяснилось, что механизм наследования ряда распространенных неврологических заболеваний (например, синдрома Хантингтона, синдрома ломкой X-хромосомы, некоторых разновидностей БАС и ранней формы болезни Альцгеймера) сравнительно прост, и каждое из таких заболеваний, судя по всему, вызывается нарушением в единственном гене. Найти гены, ответственные за эти заболевания, было довольно легко. Когда болезнетворная мутация выявлена, можно получить мышей или мух, у которых она будет проявляться, и разобраться, каким образом данная мутация вызывает соответствующее заболевание.
Благодаря знаниям об анатомическом положении, характере и механизме работы ответственных за неврологические заболевания генов врачи уже давно диагностируют такие заболевания не только на основании симптомов, связанных с поведением. Начиная с девяностых годов помимо клинических обследований врачи могут направлять пациентов на анализы, позволяющие выявить нарушения в работе определенных генов, белков и компонентов нервных клеток, а также на томографические исследования мозга, которые позволяют устанавливать, как болезнь повлияла на определенные его участки.
Установить причины психического заболевания намного сложнее, чем определить местоположение структурных повреждений в мозгу. За сотню лет исследований мозга психически больных людей патологоанатомам не удалось найти у них отчетливых локальных повреждений, которые наблюдаются у людей, страдавших неврологическими заболеваниями. Кроме того, психические заболевания затрагивают высшие психические функции. Неврозы тревоги и разные формы депрессии представляют собой эмоциональные расстройства, а шизофрения — это расстройство мышления. Эмоции и мышление суть сложные психические процессы, обеспечиваемые работой сложных систем нейронных цепей. До недавнего времени мало что было известно о нейронных цепях, задействованных в нормальных эмоциях и мышлении.
Более того, хотя большинство психических заболеваний и имеет важную генетическую составляющую, механизм их наследования непрост, потому что их вызывают не мутации в единственном гене. Таким образом, не существует какого‑то одного гена шизофрении, как не существует и гена неврозов тревоги, депрессии и большинства других психических заболеваний. Генетические составляющие этих заболеваний определяются взаимодействием нескольких генов и среды, в которой живет и развивается человек. Эффект каждого из этих генов сравнительно невелик, но вместе они дают генетическую предрасположенность, то есть потенциал для развития болезни. Большинство психических заболеваний вызывается сочетанием такой предрасположенности и дополнительных факторов среды. Например, у однояйцевых близнецов гены одинаковы. Если у одного из них будет синдром Хантингтона, он будет и у другого. Но если у одного из них разовьется шизофрения, то у другого она может развиться лишь с вероятностью около 50%. Для этого ее должны вызвать другие, негенетические факторы, действующие на ранних этапах жизни, например внутриматочная инфекция, неправильное питание или стресс. В связи со сложностью механизмов наследования основных психических заболеваний нам пока удалось найти лишь немногие из генов, задействованных в их развитии.
Перейдя от исследований имплицитной памяти аплизии к исследованиям эксплицитной памяти и внутреннего представления пространства у мышей, я перешел из сравнительно простой области в более сложную, задающую много важных вопросов, но дающую мало однозначных ответов о поведении людей. Принимаясь за исследование психических заболеваний, используя животных в качестве модельных объектов, я делал еще один шаг в глубины неопределенности. Кроме того, исследованиями имплицитной памяти у аплизии я занялся на самом раннем этапе работы, а к изучению эксплицитной памяти мышей подключился на интересном промежуточном звене, но в биологию психических заболеваний пришел сравнительно поздно. Многие другие ученые уже изучали психические заболевания на модельных объектах задолго до меня.
Недостаток знаний об анатомии, генетике и нейронных механизмах психических расстройств затруднял изучение этих расстройств на модельных объектах. Единственным явным исключением, которым я для начала и решил заняться, были неврозы тревоги. Сложно узнать, страдают ли мыши шизофренией, могут ли они бредить или галлюцинировать. Также сложно выявить мышей, страдающих психотической депрессией. Но все животные с хорошо развитой центральной нервной системой, от моллюсков до мышей, обезьян и людей, могут испытывать страх, то есть чувство тревоги. Кроме того, у каждого из этих животных страх имеет отчетливые, легко распознаваемые проявления. Поэтому животные не только боятся, но и позволяют нам отмечать, когда это с ними происходит. Мы можем, так сказать, читать их мысли. На это впервые обратил внимание Чарльз Дарвин в своем классическом труде «Выражение эмоций у человека и животных», опубликованном в 1872 году.
Важнейший биологический факт, отмеченный Дарвином и позволивший исследовать состояния тревоги на модельных объектах, состоит в том, что тревога (то есть страх как таковой) представляет собой универсальную инстинктивную и необходимую для выживания реакцию на что‑либо, угрожающее жизни, здоровью или социальному статусу. Тревога сигнализирует о потенциальной угрозе, которая требует некой приспособительной реакции. Как отмечал Фрейд, нормальная тревога помогает справиться с трудными ситуациями, тем самым способствуя росту личности. Есть две основные формы нормальной тревоги: инстинктивная (инстинктивный, или врожденный страх), встроенная в организм и в большей степени определяемая генами, и приобретенная («выученный» страх), к которой у организма может быть генетическая предрасположенность, но в целом приобретаемая с опытом. Как мы уже убедились, в ходе обучения легко может сформироваться ассоциация инстинктивной тревоги и безразличного раздражителя. Поскольку все способности, повышающие шансы на выживание, проявляют тенденцию к эволюционной консервативности, реакции инстинктивного и приобретенного страха сходны у всех представителей животного мира (рис. 25–1).
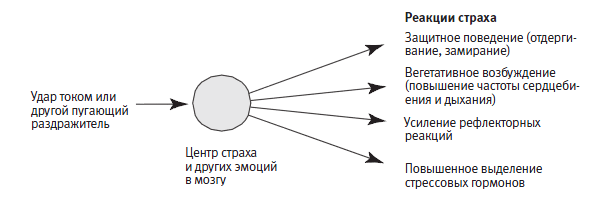
Обе формы страха могут быть и патологическими. Инстинктивная тревога имеет патологический характер, когда она чрезмерна и достаточно постоянна, чтобы парализовать способность к действиям. Приобретенная тревога имеет патологический характер, когда ее вызывают события, не представляющие реальной угрозы, как в тех случаях, когда безразличный раздражитель ассоциируется мозгом с инстинктивной тревогой. Состояния тревоги меня особенно интересовали, потому что это самое распространенное из психических расстройств: на том или ином этапе своей жизни от невроза тревоги страдают от 10% до 30% людей!
Исследуя инстинктивный и приобретенный страх у людей и подопытных животных, мы многое выяснили о поведенческих и биологических механизмах этих реакций. Одно из первых открытий из области поведенческих механизмов страха было сделано благодаря теориям Фрейда и американского философа Уильяма Джеймса, которые поняли, что страх имеет как сознательную, так и бессознательную составляющую. Оставалось неясным, как обе составляющие взаимодействуют друг с другом.
Традиционно считалось, что страх у человека начинается с сознательного восприятия какого‑либо важного события, например с того, что человек видит в своем доме пожар. Осознание этого вызывает в коре головного мозга эмоциональную реакцию (страх), запускающую сигналы, которые поступают к сердцу, кровеносным сосудам, надпочечникам и потовым железам, мобилизуя силы организма для борьбы или бегства. Таким образом, в соответствии с этими представлениями сознательная эмоциональная реакция запускает дальнейшие бессознательные, рефлекторные и вегетативные защитные реакции организма.
Джеймс отверг это представление. В опубликованной им в 1884 году и оказавшей большое влияние на развитие науки статье «Что такое эмоции?» он выдвинул идею, что когнитивный эмоциональный опыт вторичен по отношению к физиологическим выражениям эмоций. Он предположил, что, когда мы попадаем в потенциально опасную ситуацию, например видим сидящего перед нами медведя, понимание исходящей от медведя опасности еще не вызывает у нас осознанного чувства страха. Мы начнем испытывать страх лишь после того, как спасемся от медведя бегством. Вначале мы действуем инстинктивно и лишь затем запускаем когнитивные процессы, которые объясняют нам изменения, произошедшие в организме в связи с нашими действиями.
Исходя из этой идеи, Джеймс и датский психолог Карл Ланге выдвинули предположение, что мы испытываем осознанные эмоциональные ощущения уже после того, как кора головного мозга получает сигналы об изменениях физиологического состояния организма. Иными словами, сознательным ощущениям предшествуют определенные бессознательные физиологические изменения, такие как повышение или понижение давления крови, частоты сердцебиения и напряжения мышц. Поэтому при виде пожара мы испытываем страх, поскольку кора нашего мозга только что получила сигналы о том, что наше сердце заколотилось, колени задрожали, а ладони начали потеть. Джеймс писал: «Мы горюем оттого, что плачем, злимся оттого, что деремся, боимся оттого, что дрожим, а не плачем, деремся или дрожим оттого, что горюем, злимся или боимся». Согласно этому представлению, эмоции — это когнитивные реакции на информацию о состояниях организма, во многом обеспечиваемых вегетативной нервной системой. Наш повседневный опыт подтверждает, что информация, поступающая от организма, влияет на эмоциональные ощущения.
Некоторые аспекты теории Джеймса — Ланге были вскоре подтверждены экспериментальными данными. Например, выяснилось, что объективно различимые эмоции связаны с вегетативными, эндокринными и произвольными реакциями определенного характера. Более того, люди, у которых в результате несчастного случая был поврежден спинной мозг и из‑за этого в головной мозг не поступают сигналы от участков вегетативной нервной системы, расположенных ниже травмы, судя по всему, испытывают более слабые эмоции.
Однако со временем стало ясно, что теория Джеймса — Ланге объясняет лишь один из аспектов эмоционального поведения. Если бы поступающая в головной мозг информация была единственным определяющим фактором, эмоции не продолжались бы после окончания физиологических изменений. Однако наши чувства (мысли и действия, которыми мы отвечаем на эмоции) могут долгое время поддерживаться и тогда, когда нам, например, уже ничто не угрожает. И, напротив, некоторые чувства возникают у нас намного раньше, чем с организмом происходят изменения. Поэтому надо полагать, что эмоции — это не только интерпретация поступающих в мозг сигналов о происходящих в организме физиологических изменениях.
Теорию Джеймса — Ланге существенно переработал невролог Антонью Дамазью, в работах которого доказывается, что эмоциональные ощущения — это, по сути, более высокого порядка представление реакций организма, которое может быть устойчивым и продолжительным. Благодаря работам Дамазью наука приблизилась к консенсусу по вопросу о механизме возникновения эмоций. Теперь считается, что первый этап их появления — бессознательная, имплицитная оценка раздражителя, за которой следуют физиологическая реакция и, наконец, сознательное ощущение, которое может быть как продолжительным, так и непродолжительным.
Чтобы непосредственно определить, в какой степени первоначальное эмоциональное ощущение зависит от сознательных, а в какой — от бессознательных процессов, ученые прибегли к исследованию внутреннего представления эмоций с помощью тех же методов клеточной и молекулярной биологии, которые используются для изучения сознательных и бессознательных когнитивных процессов. Это удалось сделать, совместив результаты исследований, проведенных на модельных объектах, с теми, что дали исследования, проведенные на людях. В итоге за последние два десятилетия были довольно точно определены нейронные проводящие пути, лежащие в основе эмоций. Бессознательная составляющая эмоций, изученная преимущественно на модельных объектах, включает работу вегетативной нервной системы и регулирующего ее гипоталамуса. Сознательная составляющая, которую изучали на людях, включает оценочную работу коры головного мозга, осуществляемую поясной извилиной. Ключевую роль в работе обеих составляющих играет миндалевидное тело — группа собранных вместе ядер, расположенная в глубине каждого полушария. Миндалевидное тело, судя по всему, осуществляет координацию сознательного чувственного опыта и физиологических проявлений эмоций, в особенности страха.
Исследования, проведенные на людях и грызунах, показали, что нейронные системы, в которых хранятся бессознательные, имплицитные, эмоциональные воспоминания, отличаются от систем, где формируется память о сознательных, эксплицитных, чувственных состояниях. Повреждения миндалевидного тела, задействованного в работе памяти, связанной со страхом, нарушают возникновение эмоциональных реакций в ответ на раздражители, вызывающие такие реакции у здоровых людей. Повреждения гиппокампа, задействованного в работе сознательной памяти, напротив, сказываются на способности запоминать контекст, в котором происходило воздействие того или иного раздражителя. Сознательные когнитивные системы дают нам выбор возможных действий, а бессознательные механизмы эмоциональной оценки ограничивают этот выбор действиями, уместными в данной ситуации. Эта концепция хороша тем, что позволяет связать исследования эмоций с изучением механизмов работы памяти. Уже доказано, что в бессознательных проявлениях эмоциональных воспоминаний задействована имплицитная память, а в сознательном припоминании чувственных состояний — эксплицитная, значит, для него необходимо участие гиппокампа.
Одна из характерных особенностей страха состоит в том, что в результате обучения он легко ассоциируется с безразличными раздражителями. У человека, в мозгу которого возникла подобная ассоциация, безразличный раздражитель может действовать как эффективный стимул к формированию долговременных эмоциональных воспоминаний. Приобретенный страх составляет главный компонент посттравматических стрессовых расстройств, а также социальных фобий, агорафобии (боязни открытого пространства) и боязни сцены. В случае с боязнью сцены и некоторыми другими формами страха ожидания какое‑то будущее событие (например, выход на сцену) ассоциируется с перспективой того, что что‑то пойдет не так (например, выступающий забудет текст). Посттравматические стрессовые расстройства возникают вслед за событиями, вызывающими сильнейший стресс, такими как опасная для жизни борьба, пытки, изнасилование, жестокое обращение или стихийные бедствия. Эти расстройства проявляются в повторно испытываемом страхе, нередко вызываемом чем‑то, что напоминает о полученной психической травме. Одна из характерных особенностей и этих расстройств, и приобретенного страха в целом состоит в том, что воспоминания о полученной травме могут десятилетиями оставаться в силе и легко всплывать под действием различных стрессовых обстоятельств. Даже после однократного столкновения с угрозой миндалевидное тело может сохранять память об этой угрозе на протяжении всей жизни организма. Как у него это получается?
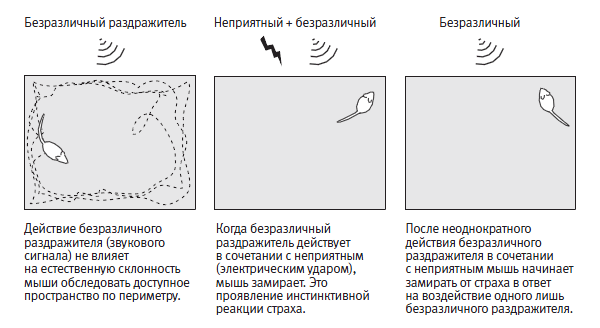
Начало моих исследований приобретенного страха у мышей было в некотором роде естественным продолжением экспериментов с аплизией. В случае с аплизией животное, у которого вырабатывается классический условный рефлекс, учится ассоциировать два раздражителя: безразличный (легкое прикосновение к сифону) и другой, достаточно сильный, чтобы вызывать инстинктивный страх (удар током в заднюю часть тела). У мышей удар током по ногам, подобно удару в заднюю часть тела у аплизии, вызывает инстинктивные реакции страха: отдергивание, прижимание к субстрату и замирание. Воздействие безразличного раздражителя, такого как простой звуковой сигнал, не вызывает таких реакций. Однако после неоднократного воздействия звукового сигнала в сочетании с ударом тока мышь обучается ассоциировать эти два раздражителя. Она запоминает, что звуковой сигнал предшествует удару током. В итоге звук сам по себе начинает вызывать у нее реакцию страха (рис. 25–2).
Хотя система нейронных цепей, лежащая в основе приобретенного страха у мышей, намного сложнее, чем соответствующая система у аплизии, о ее устройстве уже кое‑что известно благодаря исследованиям Джозефа Леду из Нью-Йоркского университета и Майла Дэвиса, работающего сейчас в Университете Эмори. Они установили, что и у грызунов, и у людей в механизмах как врожденного, так и приобретенного страха задействована определенная нейронная цепь, связанная с миндалевидным телом. Кроме того, они выяснили принципиальную схему поступления информации о безусловных и условных раздражителях в миндалевидное тело и запускания миндалевидным телом сигналов, вызывающих реакции страха.
Когда звуковой сигнал сопровождается ударом тока по ногам, информация о звуке и ударе вначале передается по двум разным путям. Звук (условный раздражитель) вызывает активацию сенсорных нейронов так называемой улитки — отдела внутреннего уха, отвечающего за восприятие звуков. Аксоны этих нейронов ведут к группе нейронов таламуса, связанных со слухом. Нейроны образуют два проводящих пути: прямой, ведущий непосредственно в боковое ядро миндалевидного тела, вообще не заходя в кору, и непрямой, ведущий сначала в слуховую кору, а затем в боковое ядро миндалевидного тела (рис. 25–3). Эти два пути, передающие информацию о звуке, заканчиваются синаптическими связями с пирамидальными нейронами, представляющими главный тип клеток бокового ядра.
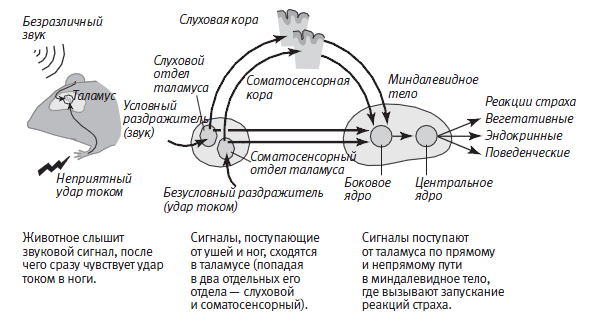
Болевое воздействие удара тока (безусловного раздражителя) вызывает активацию проводящего пути, ведущего к другой группе нейронов таламуса, осуществляющей обработку болевых сигналов. Эти нейроны тоже образуют как прямой, так и непрямой пути, ведущие к пирамидальным клеткам бокового ядра. Непрямой путь в данном случае проходит через соматосенсорную кору.
Открытие двух отдельных путей (проходящего через кору и не заходящего в нее) непосредственно доказывало, что бессознательная оценка пугающего раздражителя предшествует сознательной — в соответствии с предсказаниями теории Джеймса — Ланге. Вызывая активацию более быстрого прямого проводящего пути, не заходящего в кору, пугающий раздражитель может заставить наше сердце колотиться, а ладони — потеть еще до того, как активация более медленного пути позволит нам осознать, например, что где‑то рядом раздался выстрел.
Боковое ядро не только служит местом встречи для информации об условном (звуке) и безусловном (ударе током) раздражителях, но и вызывает мобилизацию адаптивных реакций, осуществляемую через связи нейронов этого ядра с гипоталамусом и поясной извилиной коры. Гипоталамус играет ключевую роль в физиологических проявлениях страха, запуская реакцию «борьбы или бегства» (повышенная частота сердцебиения, потоотделение, сухость во рту и напряжение мышц). Работа поясной извилины связана с сознательной оценкой страха.
Так каков же механизм работы приобретенного страха у мышей? Включает ли он изменения синаптической силы в проводящих путях под действием условного раздражителя, как это происходит у аплизии? Чтобы найти ответ, ряд ученых, в том числе я и мои коллеги, работали со срезами миндалевидного тела мышей. Результаты предшествующих исследований показали, что стимуляция как прямого, так и непрямого проводящего пути с частотой, близкой к той, что использовали Блисс и Лемо в своих экспериментах с гиппокампом, приводит к усилению этих путей, представляющему собой разновидность долговременной потенциации. Мы исследовали эту разновидность биохимическими методами и установили: хотя она немного отличается от той, что происходит в гиппокампе, зато почти ничем не отличается от долговременного усиления синаптических связей, задействованного у аплизии в сенсибилизации и выработке классического условного рефлекса (двух форм приобретенного страха). В обоих случаях работает молекулярный сигнальный путь, в котором участвуют циклический АМФ, протеинкиназа A и регуляторный белок CREB. Эти открытия в очередной раз показывали, что долговременное усиление синапсов у аплизии и различные формы долговременной потенциации у млекопитающих входят в одно семейство молекулярных процессов, позволяющих надолго усиливать синаптические связи.
В 2002 году моим сотрудником стал Майкл Роган, ранее работавший у Джозефа Леду, и мы с ним перешли от изучения срезов мозга мышей к экспериментам с интактными животными. Мы исследовали реакцию нейронов миндалевидного тела мышей на звуковой сигнал и обнаружили то же, что Роган и Леду выявили ранее про крыс, а именно: приобретенный страх усиливает эту реакцию (рис. 25–4). Такое явление напоминало долговременное усиление синаптических связей, которое мы наблюдали в срезах миндалевидного тела. Тогда сотрудничавший с нами Вадим Большаков из Гарварда предположил, что если синапсы в миндалевидном теле интактной мыши усилены приобретенным страхом, то электрическая стимуляция синапсов в срезах миндалевидного тела той же мыши не будет приводить к дальнейшему существенному усилению этих синапсов. Так и оказалось. То есть изменения, происходящие в миндалевидном теле живых мышей в результате обучения, похожим образом проявляются в тех же точках, что и изменения, вызываемые электрической стимуляцией срезов миндалевидного тела.
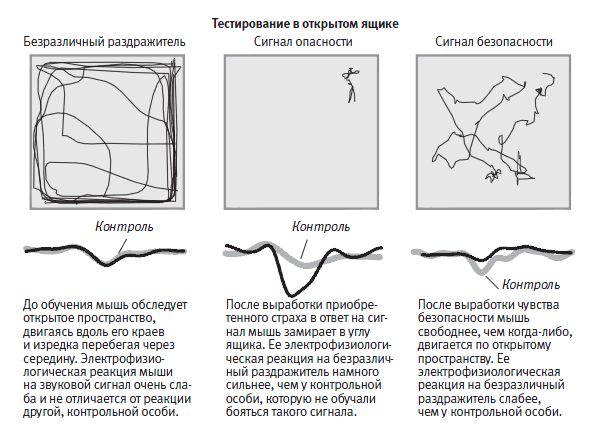
Затем мы воспользовались хорошо отработанным методом поведенческого тестирования на приобретенный страх. Мы помещали мышь в большой открытый ярко освещенный ящик. Мыши — ночные животные, поэтому они боятся яркого света и в норме будут носиться по дну ящика вдоль стенок, лишь изредка забегая в его середину. Такая форма защитного поведения представляет собой компромисс между потребностью животного в обследовании окружающей среды и в избегании хищников. Когда мы включали звуковой сигнал, мышь продолжала бегать вдоль стенок ящика как ни в чем не бывало. Но после того как мы неоднократно сопровождали звуковой сигнал ударом тока, мышь обучалась ассоциировать эти два раздражителя. Теперь, услышав звук, она переставала бегать вдоль стенок, изредка забегая в середину, и сидела, прижавшись ко дну, в одном из углов, обычно в позе замирания (рис. 25–2).
Разобравшись в особенностях анатомии и физиологии приобретенного страха, мы почувствовали, что можем изучить молекулярные основы этого явления. Вместе с постдоком Глебом Шумяцким я занялся поиском генов, которые экспрессировались бы только в боковом ядре миндалевидного тела — уже исследованном нами участке. Мы выяснили, что в пирамидальных клетках экспрессируется ген, который кодирует определенный нейромедиатор — так называемый гастрин-высвобождающий пептид. Пирамидальные клетки используют этот пептид в качестве возбуждающего нейромедиатора в дополнение к глутамату и совместно с ним, выделяя эти медиаторы из пресинаптических окончаний, ведущих к клеткам-мишеням бокового ядра. Затем мы обнаружили, что клетки-мишени относятся к особой популяции тормозных интернейронов, в мембране которых содержатся рецепторы к гастрин-высвобождающему пептиду. Как и все тормозные интернейроны бокового ядра, клетки-мишени выделяют нейромедиатор ГАМК. Клетки-мишени могут передавать сигналы обратно на пирамидальные клетки и, активируясь, выделяют в синаптические щели ГАМК, тем самым тормозя работу пирамидальных клеток.
Обнаруженная нами нейронная цепь представляет собой так называемую систему с отрицательной обратной связью: возбуждающий нейрон активирует тормозный, который, в свою очередь, тормозит активировавший его возбуждающий нейрон. Не может ли такая система служить для контроля над страхом? Чтобы это выяснить, мы тестировали генетически модифицированных мышей, у которых был отключен ген рецептора, реагирующего на гастрин-высвобождающий пептид, а значит, вышеописанная система с обратной связью не могла работать. Из нашего предположения следовало, что повышенная возбудимость таких мышей могла приводить к усиленному бесконтрольному страху.
В соответствии с нашим предположением в боковом ядре миндалевидного тела этих мышей мы наблюдали резко усиленную долговременную потенциацию наряду с достоверно повышенной и более продолжительной памятью, связанной со страхом. Примечательно, что такой эффект затрагивал только приобретенный страх: те же мутантные мыши демонстрировали в разных других тестах нормальный врожденный страх. Этот факт согласуется с тем, что известно о принципиальных отличиях приобретенного страха от врожденного. Так комплексный подход с использованием методов клеточной биологии и молекулярной генетики позволил нам найти нейронную цепь, которая играет важную роль в осуществлении контроля над приобретенным страхом. Это открытие могло привести к разработке препаратов, которые позволят бороться с приобретенным страхом, связанным с такими синдромами, как посттравматические стрессовые расстройства и фобии.
А как обстоят дела с противоположностью страха — чувством безопасности, уверенности в себе и счастья? Не могу не вспомнить первое предложение «Анны Карениной», романа Льва Толстого о трагических последствиях любовной связи, неприемлемой в глазах общества: «Все счастливые семьи похожи друг на друга, каждая несчастливая семья несчастлива по‑своему». Это утверждение, в котором больше литературной, чем научной силы, предполагает, что тревога и депрессия могут принимать много разных форм, в то время как позитивные эмоции (чувство благополучия, безопасности и счастья) имеют общие черты.
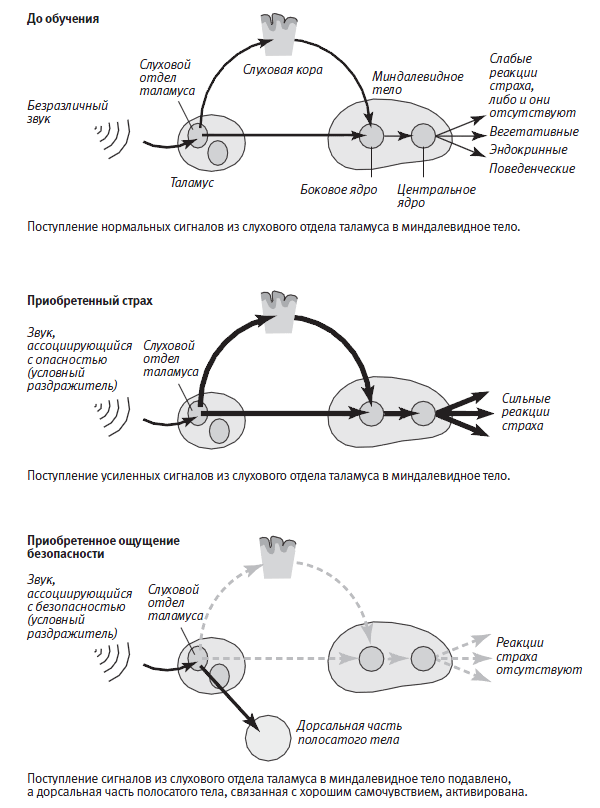
Чтобы это проверить, мы с Роганом исследовали нейробиологические свойства приобретенного чувства безопасности, которое можно считать разновидностью чувства счастья. Мы рассуждали следующим образом. Когда звуковой сигнал действует в сочетании с ударом тока, животное запоминает, что этот сигнал предвещает удар тока. Так что если звуковой сигнал и удар тока никогда не будут действовать в сочетании, животное запомнит, что сигнал не предвещает удара, а предвещает, напротив, безопасную ситуацию. Проведя эксперимент, мы получили в точности такой результат, какой и предсказывали: когда попавшая в новую среду мышь, на которую до того воздействовали звуковыми сигналами и ударами тока, никогда не сочетавшимися друг с другом, слышала тот же звук, она переставала демонстрировать защитное поведение. Она расхаживала в центре открытого пространства как у себя дома, не проявляя никаких признаков страха (рис. 25–5). Исследовав боковое ядро миндалевидного тела мышей, обученных подобным образом, мы обнаружили эффект, противоположный долговременной потенциации, а именно долговременное подавление нейронной реакции на звук, предполагавшее, что поступление сигналов в миндалевидное тело было резко ограничено (рис. 25–4).
Затем мы задались вопросом, дает ли такое обучение подлинное чувство безопасности, то есть настоящее ощущение уверенности в себе, или же просто снижает базовый уровень тревоги, всегда присутствующей в каждом. Чтобы отличить одно от другого, мы регистрировали сигналы в полосатом теле — участке мозга, который в норме задействован в положительном подкреплении и в обеспечении чувства благополучия. (Именно этот участок активируется кокаином и другими вызывающими привыкание наркотиками, которые вторгаются в нейронную систему положительного подкрепления и заставляют человека принимать наркотик все чаще.) Оказалось, что, когда у животного формируется приобретенный страх, то есть оно обучается ассоциировать звуковой сигнал с ударом тока, нейронная активность в его полосатом теле не меняется. Но когда животное обучается ассоциировать звуковой сигнал с безопасностью, реакция полосатого тела заметно усиливается, что соответствует связанному с безопасностью позитивному ощущению.
Наши исследования приобретенного чувства безопасности позволили в новом свете увидеть позитивные ощущения счастья и благополучия, а также негативные чувства тревоги и страха. Они указывали на существование в глубинах мозга второй системы, связанной с позитивными эмоциями, поскольку и нейроны таламуса, реагирующие на звуковой сигнал, и нейроны бокового ядра миндалевидного тела связаны с полосатым телом и передают в него информацию о счастье и благополучии. Полосатое тело, в свою очередь, связано со многими участками мозга, в том числе с префронтальной корой, которая подавляет активность миндалевидного тела. Поэтому есть все основания полагать, что приобретенное чувство безопасности, усиливая сигналы, поступающие в полосатое тело, не только увеличивает ощущение безопасности и благополучия, но и ослабляет ощущение тревоги, подавляя активность миндалевидного тела.
Судя по этим результатам, мы, возможно, стоим на пороге эпохи, когда молекулярная биология когнитивных функций и эмоций откроет новые способы повышения самооценки и ощущения благополучия. Не окажется ли при этом, например, что некоторые неврозы тревоги вызываются нарушениями работы нейронных сигналов, которые в норме обеспечивают чувство благополучия? В нашем распоряжении начиная с шестидесятых годов имеются лекарства, позволяющие облегчать ряд форм тревоги, но эти препараты помогают не при всех неврозах тревоги, лишь при некоторых, а, например, лоразепам и диазепам вызывают привыкание, поэтому за их использованием приходится очень внимательно следить. Средства, усиливающие активность нейронных цепей, которые обеспечивают ощущение безопасности и хорошее самочувствие, могут дать нам более эффективный способ борьбы с неврозами тревоги.
26. Новый способ лечения психических заболеваний
Можно ли использовать мышей в качестве модельных объектов для изучения других психических расстройств, более сложных, более серьезных и тяжелых, чем неврозы тревоги? Можно ли использовать их для изучения шизофрении — самого стойкого и разрушительного из психических заболеваний человека, которое сильнее других нуждается в новых методах лечения?
Шизофрения, как ни странно, распространена довольно широко. От нее страдает около 1% населения Земли. У мужчин она, по‑видимому, встречается несколько чаще, чем у женщин, и принимает более тяжелые формы. Еще 2–3% людей страдают шизотипичным личностным расстройством, которое нередко считают более мягкой формой той же болезни, не вызывающей явного психотического поведения.
Шизофрения характеризуется тремя типами симптомов: позитивными, негативными и когнитивными. Позитивные симптомы наблюдаются в течение не меньше шести месяцев и представляют собой странное и иногда очень причудливое поведение, сопровождаемое нарушениями психических функций. Эти симптомы особенно заметны во время психотических эпизодов — фаз заболевания, во время которых пациент не способен правильно трактовать действительность. В это время пациент не может реалистично оценивать свои убеждения и плоды восприятия и сравнивать их с тем, что на самом деле происходит в окружающем мире. Характерным признаком неспособности трактовать действительность служат иллюзии (ненормальные убеждения, противоречащие фактам, которые не меняются под действием доказательств их неразумности), галлюцинации (плоды восприятия, не вызываемые внешними раздражителями, например внутренние голоса, комментирующие поступки человека) и нелогичное мышление (утрата нормальных связей или ассоциаций между идеями — так называемое ослабление или крушение ассоциаций, в тяжелых формах приводящее к бессвязности мыслей и речи).
Негативные симптомы шизофрении — это отсутствие некоторых форм нормального социального и межличностного поведения, сопровождающееся самоизоляцией, бедностью речи и утратой способности испытывать и выражать эмоции (так называемым уплощением аффекта). Когнитивные симптомы включают расстройства внимания и нарушения одной из форм эксплицитной кратковременной памяти — так называемой рабочей памяти, играющей ключевую роль в осуществлении исполнительных функций типа организации распорядка дня или планирования и выполнения какой‑либо последовательности действий. Когнитивные симптомы шизофрении носят хронический характер и сохраняются даже во время непсихотических периодов. Бороться с такими симптомами особенно сложно.
В промежутках между психотическими эпизодами пациенты проявляют преимущественно негативные и когнитивные симптомы: они ведут себя эксцентрично, изолируются от общества и демонстрируют низкий уровень эмоциональной активации, обедненные социальные потребности, бедность речи, пониженный объем внимания и недостаток мотивации.
Большинство исследователей шизофрении давно отметило, что весь спектр ее симптомов невозможно изучать на таком модельном объекте, как мыши. Позитивные симптомы у мышей сложно наблюдать, потому что мы не знаем, как выявлять их иллюзии и галлюцинации. Столь же сложно наблюдать у них негативные симптомы. Но после новаторских экспериментов на обезьянах, поставленных Патрисией Голдман-Ракич из Йельского университета, я и мои коллеги Элинор Симпсон, Кристоф Келлендонк и Джонатан Полан решили выяснить, нельзя ли использовать мышей в качестве модельных объектов для изучения молекулярных основ некоторых аспектов когнитивных симптомов шизофрении. Мы полагали, что сможем моделировать такой ключевой компонент когнитивных симптомов, как нарушение рабочей памяти. Рабочая память уже была хорошо описана, и мы знали, что для ее функционирования необходима префронтальная кора — часть лобной доли, обеспечивающая работу самых сложных из наших психических процессов. Кроме того, мы полагали, что, если сможем разобраться в этих когнитивных нарушениях, будем лучше представлять и то, как функционирует префронтальная кора в нормальных психических состояниях.
Исследования префронтальной коры начались еще в 1848 году, когда Джон Харлоу описал ставший впоследствии знаменитым случай с пациентом по имени Финеас Гейдж, работавшим прорабом на строительстве железной дороги. В результате взрыва его голову пробил металлический прут, прошедший сквозь префронтальную кору. Гейдж выжил, и его общий интеллект, восприятие и долговременная память не пострадали, но у него произошли изменения личности. До получения травмы он был добросовестным и трудолюбивым, а после запил и стал ненадежным работником, постоянно менявшим работу. Последующие исследования людей с повреждениями префронтальной коры подтвердили, что этот участок мозга играет ключевую роль в оценочной деятельности и долговременном планировании.
В тридцатых годах психолог Карлайл Якобсен из Йельского университета начал исследования функций префронтальной коры обезьян и получил первые свидетельства того, что она задействована в работе кратковременной памяти. Сорок лет спустя британский специалист по когнитивной психологии Алан Бэддли описал форму кратковременной памяти, которую назвал рабочей, потому что она суммирует сиюминутные ощущения за сравнительно краткий период и связывает их с закрепленными воспоминаниями о предшествующем опыте, что необходимо для планирования и осуществления сложных форм поведения. Вскоре после этого Хоакин Фустер из Калифорнийского университета в Лос-Анджелесе и Патрисия Голдман-Ракич связали результаты, полученные Якобсеном в его исследованиях префронтальной коры, с результатами, которых достиг Бэддли в исследованиях рабочей памяти. Они выяснили, что удаление префронтальной коры у обезьян приводит к нарушению не всей кратковременной памяти, а лишь тех ее функций, которые Бэддли описал под названием рабочей.
Открытие того, что префронтальная кора задействована в планировании и осуществлении сложных форм поведения (функций, которые нарушаются при шизофрении), заставило исследователей заняться изучением префронтальной коры пациентов, страдающих шизофренией. Результаты функциональной томографии мозга показали, что метаболическая активность в префронтальной коре таких пациентов ниже нормы, даже если они не заняты никакой особенной психической деятельностью. Когда здоровые испытуемые получают задания, для выполнения которых нужна рабочая память, метаболическая активность в их префронтальной коре сильно повышается. У шизофреников при этом повышение метаболической активности более слабое.
Учитывая, что у шизофрении есть генетическая составляющая, нет ничего удивительного, что умеренные нарушения рабочей памяти наблюдаются также у 40–50% родственников первой степени родства (родителей, детей, братьев и сестер) пациентов, страдающих шизофренией, хотя у них нет клинических симптомов шизофрении. Более того, у тех же родственников больных наблюдаются аномалии в работе префронтальной коры, что лишний раз подчеркивает значение этого участка мозга для генетической предрасположенности к шизофрении.
Тот факт, что когнитивные симптомы шизофрении напоминают нарушения поведения, наблюдаемые у подопытных животных, у которых фронтальные доли коры были хирургическим путем отделены от остального мозга, заставил нас задаться вопросом, каковы молекулярные основы нарушений рабочей памяти, связанных с префронтальной корой.
Значительную часть того, что мы знаем о биологии шизофрении, удалось выяснить в поисках лекарственных препаратов для борьбы с этим заболеванием. В пятидесятых годах французский нейрохирург Анри Лабори выдвинул идею, что тревога, которую многие пациенты испытывают перед операцией, может быть вызвана тем, что их организм в большом количестве выделяет гистамин. Это сходное с гормонами вещество выделяется в состоянии стресса и вызывает расширение кровеносных сосудов и понижение давления крови. Лабори доказывал, что избыток гистамина может участвовать в возникновении некоторых неблагоприятных побочных эффектов обезболивания, таких как тревожное возбуждение, шок и скоропостижная смерть. В ходе поисков препарата, который подавлял бы действие гистамина и успокаивал пациентов, Лабори вышел на хлорпромазин, недавно разработанный французской фармацевтической фирмой Rhoˆne-Poulenc. Успокоительное действие хлорпромазина произвело на Лабори такое впечатление, что он задумался, нельзя ли использовать этот препарат для успокоения возбужденных пациентов, страдающих психическими расстройствами. Французские психиатры Жан Деле и Пьер Деникер занялись разработкой этой идеи и выяснили, что высокие дозы хлорпромазина действительно успокаивают возбужденных и агрессивных пациентов с симптомами шизофрении.
Впоследствии выяснилось, что хлорпромазин и близкие ему препараты можно использовать не только как транквилизаторы, эффективно, но не чрезмерно успокаивающие пациентов, но и в качестве антипсихотических средств, успешно подавляющих психотические симптомы шизофрении. Эти препараты, первые эффективные средства борьбы с серьезным психическим заболеванием, произвели революцию в психиатрии. Кроме того, они привлекли внимание психиатров к вопросу о механизме действия подобных средств.
Первые сведения о механизме действия хлорпромазина были получены в ходе исследований одного из его побочных эффектов — синдрома, напоминающего болезнь Паркинсона. В 1960 году Арвид Карлссон, профессор фармакологии из Гетеборгского университета в Швеции, впоследствии разделивший со мной Нобелевскую премию, сделал три замечательных открытия, которые позволили разобраться в принципиальных вопросах, связанных с болезнью Паркинсона и шизофренией. Во-первых, он открыл дофамин и показал, что это вещество играет в мозгу роль нейромедиатора. Во-вторых, он выяснил, что если опустить концентрацию дофамина в мозгу подопытного животного ниже определенного порогового уровня, то у животного разовьется подобие болезни Паркинсона. Исходя из этих данных Карлссон доказывал, что болезнь Паркинсона может быть результатом понижения концентрации дофамина в участках мозга, задействованных в регуляции моторики. Он и его коллеги проверили это предположение и выяснили, что, давая пациентам препараты дофамина, можно остановить развитие симптомов болезни Паркинсона.
В ходе этих исследований Карлссон отметил, что, когда пациентам давали слишком большие дозы дофамина, у них возникали психотические симптомы, напоминающие проявления шизофрении. Это наблюдение заставило его предположить, что первопричиной шизофрении служит перепроизводство дофамина и что терапевтическое действие антипсихотических средств связано с тем, что они блокируют работу дофаминовых рецепторов, ослабляя дофаминовую передачу в каких‑то ключевых нейронных путях и тем самым уменьшая эффект перепроизводства дофамина. Впоследствии предположение Карлссона получило экспериментальные подтверждения. Еще одно свидетельство в пользу этой идеи было получено, когда выяснилось, что в качестве побочного эффекта у пациентов, которых лечили антипсихотическими средствами, нередко возникали симптомы, напоминающие проявления болезни Паркинсона, что тоже говорило в пользу предположения о подавлении этими препаратами работы дофамина в мозгу.
Карлссон считал, что чрезмерная активность производящих дофамин нейронов ответственна за все симптомы шизофрении: позитивные, негативные и когнитивные. Он предположил, что избыток дофамина в проводящем пути, ведущем в гиппокамп, миндалевидное тело и другие связанные с ними структуры, может вызывать позитивные симптомы, а избыток дофамина в проводящем пути, ведущем в кору, особенно учитывая множество синаптических связей, которые этот путь образует с префронтальной корой, может вызывать негативные и когнитивные симптомы. Со временем стало ясно, что все лекарства, подавляющие симптомы шизофрении, действуют преимущественно на один тип дофаминовых рецепторов — рецепторы D2. Соломон Снайдер из Университета Джонса Хопкинса и Филип Симан из Торонтского университета обнаружили сильную связь между эффективностью антипсихотических препаратов и их способностью блокировать рецепторы D2. В то же время выяснилось, что антипсихотические препараты помогают только против позитивных симптомов шизофрении. Они ослабляют или даже устраняют иллюзии, галлюцинации и некоторые типы расстройств мышления, но не оказывают существенного влияния на негативные и когнитивные симптомы болезни. Объяснить это избирательное действие было трудно.
В 2004 году работы ряда исследователей показали, что одним из факторов генетической предрасположенности к шизофрении служит аномально высокая концентрация рецепторов D2 в полосатом теле — области мозга, которая, как мы уже убедились, обычно задействована в обеспечении хорошего самочувствия. Повышенная концентрация рецепторов D2, способных связывать дофамин, приводит к усилению дофаминовой передачи. Симпсон, Келлендонк, Полан и я решили исследовать роль этого фактора генетической предрасположенности в возникновении когнитивных нарушений, связанных с шизофренией. Для этого мы получили генетически модифицированных мышей, у которых работал ген, обеспечивающий перепроизводство рецепторов D2 в полосатом теле. Оказалось, что у таких мышей действительно наблюдаются нарушения рабочей памяти, в соответствии с предсказаниями гипотезы Карлссона.
Нам хотелось узнать, почему препараты, блокирующие работу рецепторов D2, не могут подавить когнитивные симптомы шизофрении. Для этого мы провели еще один эксперимент, воспользовавшись генетическими методами, которые были разработаны нами десятью годами раньше. Когда мышь становилась взрослой, мы отключили внедренный в нее ген, вызывающий перепроизводство рецепторов дофамина, и обнаружили, что это не привело к устранению нарушений рабочей памяти. Иными словами, исправление молекулярного дефекта в мозгу взрослого организма не исправляло наблюдаемого когнитивного дефекта.
Результат заставлял предположить, что переизбыток рецепторов D2 в процессе развития вызывает в мозгу изменения, которые сохраняются и у взрослого организма. Возможно, именно из‑за таких изменений антипсихотические препараты не помогали бороться с когнитивными симптомами шизофрении. Перепроизводство рецепторов D2 в полосатом теле начинает действовать уже на ранних стадиях развития, задолго до появления первых симптомов болезни, возможно, за счет того, что вызывает устойчивые и необратимые изменения дофаминовой системы в какой‑то другой части мозга. После этого нарушения работы префронтальной коры и структур полосатого тела, задействованных в когнитивных симптомах шизофрении, уже нельзя исправить, понизив концентрацию рецепторов D2 до нормального уровня.
К настоящему времени нам удалось найти по крайней мере одно изменение, происходящее в префронтальной коре в результате перепроизводства рецепторов D2: подавление активации другого типа дофаминовых рецепторов — D1. Результаты экспериментов, проведенных ранее Патрисией Голдман-Ракич, заставляли предположить, что подавление активации рецепторов D1 вызывает снижение концентрации циклического АМФ, что приводит к нарушению рабочей памяти.
Эксперименты доказали, что генетически модифицированных мышей можно с успехом использовать в качестве модельных объектов для изучения сложных психических заболеваний. Эти объекты позволяют нам разбивать заболевание на более простые и лучше поддающиеся анализу молекулярные компоненты. С помощью мутантных мышей мы можем не только исследовать генетические факторы, приводящие к шизофрении, но и, манипулируя средой развития мышей в утробе матери и на ранних стадиях после рождения, изучать взаимодействия генов и среды, которые могут вызывать развитие болезни.
Депрессия, еще одно распространенное заболевание, разрушительно влияющее на психическое здоровье, была впервые описана в V веке до н. э. древнегреческим врачом Гиппократом, который полагал, что настроение определяется равновесием четырех «соков» организма: крови, слизи, желтой желчи и черной желчи. Считалось, что депрессию вызывает избыток черной желчи. Само слово «меланхолия», которым древние греки называли депрессию, означает черную желчь. Хотя Гиппократово объяснение депрессии и кажется нам фантастическим, лежащее в его основе представление о том, что психические заболевания являются отражением физиологических процессов, получает в наши дни всеобщее признание.
Клинические признаки депрессии можно лаконично описать словами Гамлета: «Каким докучным, тусклым и ненужным мне кажется все, что ни есть на свете!» 1 Если не лечить депрессию, отдельный ее эпизод обычно длится от четырех месяцев до года. Для нее характерны плохое настроение, которое сохраняется большую часть времени изо дня в день, а также сильная душевная боль, неспособность испытывать удовольствие и общая потеря интереса к окружающему. Депрессия нередко сопровождается бессонницей, потерей аппетита, веса, подавленным половым влечением и замедленным мышлением.
На том или ином этапе жизни эпизоды депрессии переживает около 5% людей. В Соединенных Штатах в каждый отдельно взятый момент от депрессии страдают около 8 млн человек. Глубокие формы депрессии могут быть очень серьезными: в наиболее тяжелых случаях пациент способен прекратить питаться или соблюдать элементарные правила личной гигиены. Хотя некоторые люди испытывают за свою жизнь только один эпизод депрессии, обычно у этой болезни бывают рецидивы. Около 70% людей, испытавших один серьезный эпизод депрессии, рано или поздно испытывают по крайней мере еще один. Средний возраст, в котором развивается депрессия, составляет около двадцати восьми лет, но первый эпизод может произойти почти в любом возрасте. Депрессия способна поражать даже маленьких детей, хотя у них она обычно остается недиагностированной. Эпизоды депрессии бывают и у пожилых, причем нередко у тех, у кого раньше не было таких эпизодов, у них лечить депрессию сложнее. Женщины страдают от депрессии в два-три раза чаще, чем мужчины.
Для борьбы с депрессией было разработано несколько эффективных средств. Первое — ингибитор моноаминоксидазы (ИМАО) — первоначально было получено для борьбы с совсем другой болезнью, туберкулезом. Действие ИМАО связано с подавлением расщепления серотонина и норадреналина, благодаря чему в синапсах оказывается больше нейромедиаторов, которые могут выделяться в синаптическую щель. Врачи вскоре заметили, что пациенты, употреблявшие ИМАО, отличались на удивление хорошим настроением, хотя по‑прежнему страдали от серьезной болезни. Вскоре врачи выяснили, что ИМАО — более эффективное средство для борьбы с депрессией, чем с туберкулезом. Это открытие привело к разработке целого класса препаратов, которые сегодня успешно помогают 70% пациентов, страдающих глубокой депрессией.
Открытие антидепрессантов, последовавшее за открытием антипсихотических средств, означало для психиатрии старт новой эпохи. Теперь психиатрия перестала быть областью, которая мало что могла сделать для тяжелобольных пациентов: в ее распоряжении был набор действенных лекарственных средств, сравнимый с таковым в других областях медицины.
Эффективные средства для борьбы с депрессией действуют преимущественно на две системы мозга, в которых задействованы модуляторные нейромедиаторы серотонин и норадреналин. Однозначные данные получены по серотонину, работа которого напрямую связана с настроением человека: повышенные концентрации серотонина сопряжены с хорошим самочувствием, пониженные — с симптомами депрессии. Более того, у людей, совершающих самоубийство, концентрация серотонина обычно сильно понижена.
Самые эффективные из известных антидепрессантов — так называемые селективные ингибиторы обратного захвата серотонина. Эти препараты повышают концентрацию серотонина в мозгу, подавляя (ингибируя) работу системы молекулярного переноса, которая позволяет захватывать серотонин обратно из синаптической щели, куда его выделяет пресинаптический нейрон. На основании этого открытия была выдвинута гипотеза, согласно которой депрессию вызывает недостаток серотонина, норадреналина или обоих веществ в мозгу.
Хотя эта гипотеза и позволяет объяснить некоторые стороны реакции пациентов на антидепрессанты, она не объясняет ряд других важных явлений. В частности, почему при использовании антидепрессантов, которые всего за несколько часов подавляют обратный захват серотонина нейронами, требуется не менее трех недель на то, чтобы ослабить симптомы депрессии. Если все действие антидепрессантов связано с подавлением обратного захвата и последующим накоплением серотонина в синапсах, то как объяснить задержку реакции на эти препараты? Возможно, не менее трех недель требуется, чтобы серотонин повлиял на ключевые нейронные цепи во всем мозгу — чтобы мозг «обучился» тому, как снова стать счастливым. Кроме того, теперь мы знаем, что антидепрессанты влияют и на другие процессы, а не только на обратный захват и накопление серотонина.
Одно из важнейших открытий, касающихся депрессии, сделали Рональд Думан из Йеля и Рене Хен из Колумбийского университета. Они выяснили, что антидепрессанты также повышают способность небольшого участка гиппокампа — зубчатой извилины — производить новые нервные клетки. Подавляющее большинство нервных клеток не делится, но в этом небольшом гнезде стволовых клеток происходит деление, благодаря которому возникают новые дифференцированные нервные клетки. В течение двух или трех недель (времени, которое требуется на то, чтобы антидепрессанты заработали) некоторые из этих клеток входят в состав нейронных цепей зубчатой извилины. Функции этих стволовых клеток не ясны. Чтобы их исследовать, Хен с помощью радиации разрушал зубчатую извилину мышей, служивших модельными объектами для изучения депрессии, вызываемой стрессом. Оказалось, что антидепрессанты уже не подавляли депрессивных симптомов у мышей, лишенных этих стволовых клеток.
Такие замечательные открытия говорят о возможности того, что воздействие антидепрессантов на поведение отчасти связано именно со стимуляцией производства новых нейронов в гиппокампе. Эта идея согласуется с данными о том, что депрессия нередко приводит к серьезным нарушениям памяти. Возможно, повреждения мозга, вызываемые депрессией, способны исправляться благодаря умению гиппокампа производить новые нервные клетки. Прекрасная идея! Для ее разработки и проверки потребуются воображение и способности нового поколения исследователей психики, которое будет работать в ближайшие десятилетия.
Молекулярной биологии, очевидно, предстоит сделать для психиатрии то, что она уже начала делать для неврологии. Использование генетически модифицированных мышей в качестве модельных объектов для изучения важнейших психических заболеваний может служить этому делу по крайней мере в двух направлениях. Во-первых, благодаря тому что изучение психически больных людей позволило найти ряд генов, способствующих предрасположенности к психическим заболеваниям (например, разновидность гена рецептора D2, которая служит фактором риска для заболевания шизофренией), эти гены можно внедрять в геном мышей и использовать для проверки различных гипотез о происхождении и развитии каждого из таких заболеваний. Во-вторых, генетические исследования, проводимые на мышах, позволят изучать лежащие в основе заболеваний сложные молекулярные сигнальные пути подробнее и точнее, чем на людях. Такие фундаментальные нейробиологические исследования дадут дополнительные возможности для диагностирования и классификации психических заболеваний и рациональную основу для разработки новых лекарственных средств.
В целом мы сейчас переходим из десятилетия, посвященного исследованию тайн работы мозга, в десятилетие, посвященное поиску средств лечения нарушений его работы. За пятьдесят лет, прошедших с тех пор, как я начал заниматься медициной, фундаментальная наука и клинические исследования перестали быть двумя разобщенными дисциплинами. Некоторые из интереснейших вопросов, изучаемых сегодня нейробиологией, непосредственно связаны с актуальными проблемами неврологии и психиатрии. В результате работа на стыке обеих дисциплин перестала быть уделом немногих людей в белых халатах. Теперь перспективы медицинского применения служат одним из направляющих факторов значительной части нейробиологических исследований.
За девяностые годы, которые теперь называют «десятилетием мозга», все мы стали работать на стыке фундаментальной науки и медицины. Благодаря нашим достижениям первое десятилетие XXI века стало «десятилетием лечения заболеваний мозга». В результате такие дисциплины, как психиатрия и неврология, сближаются друг с другом. Можно предвидеть, что в недалеком будущем молодые врачи, проходящие резидентуру по каждой из этих дисциплин, будут в течение одного года обучаться вместе, подобно тому как те, кто собирается выбрать целый ряд медицинских специальностей, от кардиологии до гастроэнтерологии, в течение года занимаются в резидентуре внутренней медициной в целом.
1 Перевод М. Л. Лозинского








