Способы адаптации бактерий к разным температурам оказались предсказуемыми

Команда Ричарда Ленски, микробиолога-эволюциониста, известного своим долгосрочным эволюционным экспериментом, изучила генетические различия (накопленные мутации) между популяциями бактерий, в течение 2000 поколений адаптировавшихся к пяти разным температурным режимам. Несмотря на разнообразие возможных путей адаптации большая часть мутаций оказалась специфичной, то есть они возникали в тех и только тех популяциях, которые развивались при одном и том же или при сходных температурных режимах. Однако большинство этих мутаций позже закрепились и в ходе продолжающегося долгосрочного эволюционного эксперимента. По-видимому, при разной температуре разные мутации оказываются полезнее других (но редко из полезных становятся вредными) — и естественный отбор работает именно с этими различиями в относительной полезности, закрепляя раньше мутации, оказавшиеся в данных условиях более благоприятными.
«Элементы» неоднократно рассказывали о работе лаборатории Ричарда Ленски (Richard Lenski), уже многие десятилетия занимающейся экспериментальным изучением эволюции бактерий. Эти исследования не только позволяют лучше понять закономерности приспособления этих микроорганизмов к среде (которые могут иметь и практическое значение, взять хотя бы устойчивость к антибиотикам), но и дают возможность воочию увидеть нетривиальные эволюционные процессы и их результаты, что важно для фундаментальной науки (см. Ранние этапы адаптации предсказуемы, поздние — случайны, «Элементы», 03.03.2015; В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011; Подведены итоги эволюционного эксперимента длиной в 40 000 поколений, «Элементы», 01.11.2009).
Но, кроме широко известного долговременного эволюционного эксперимента (ДЭЭ) (см. E. coli Long-term Experimental Evolution Project ), на данный момент длящегося уже 50 000 поколений, команда Ричарда Ленски проводит и более кратковременные исследования, являющиеся «отводками» основного эксперимента.
В новой работе исследователи изучали генетическую основу адаптации кишечной палочки Escherichia coli к разным температурам на протяжении 2000 поколений в нескольких десятках линий. В этой ситуации можно было ожидать как дивергенцию за счет стохастических процессов — возникновения мутаций и генетического дрейфа (см. Genetic drift), — так и параллельную эволюцию, когда разные линии приобретали бы в одной и той же среде сходные адаптации. По крайней мере, для сложно устроенных животных, у которых индивидуальное развитие существенно усложняет путь от генотипа к фенотипу, эти два варианта не обязательно противоречат друг другу: одна и та же адаптация может возникать на разной генетической основе. С этой множественностью допустимых результатов (подчеркнем, что каждый из них оказывается возможным) связан и давно интригующий ученых вопрос о предсказуемости эволюции — вернее, вопрос состоит в том, на каком уровне мы можем предсказывать то, как конкретная популяция приспособится к изменившейся тем или иным образом окружающей среде (см. D. L. Stern, V. Orgogozo, 2009. Is genetic evolution predictable?).
В эксперименте 30 отдельных линий кишечной палочки, взятые из долгосрочного эксперимента, в течение 2000 поколений развивались при пяти разных температурных режимах, по шесть линий на каждый. Часть бактерий выращивали при оптимальных 37°C (напомню, что в норме эти бактерии населяют кишечник теплокровных животных) и «терпимых» 32°C, а часть — при экстремальных для них холоде (20°C) и жаре (42°C), находящихся на нижней и верхней границах зоны толерантности (см. Реакция организма на изменение экологических факторов). Кроме того, еще один режим включал в себя чередование умеренных и экстремальных условий — температура колебалась между 32°C и 42°C. Подробнее схема эксперимента представлена на рис. 2А.
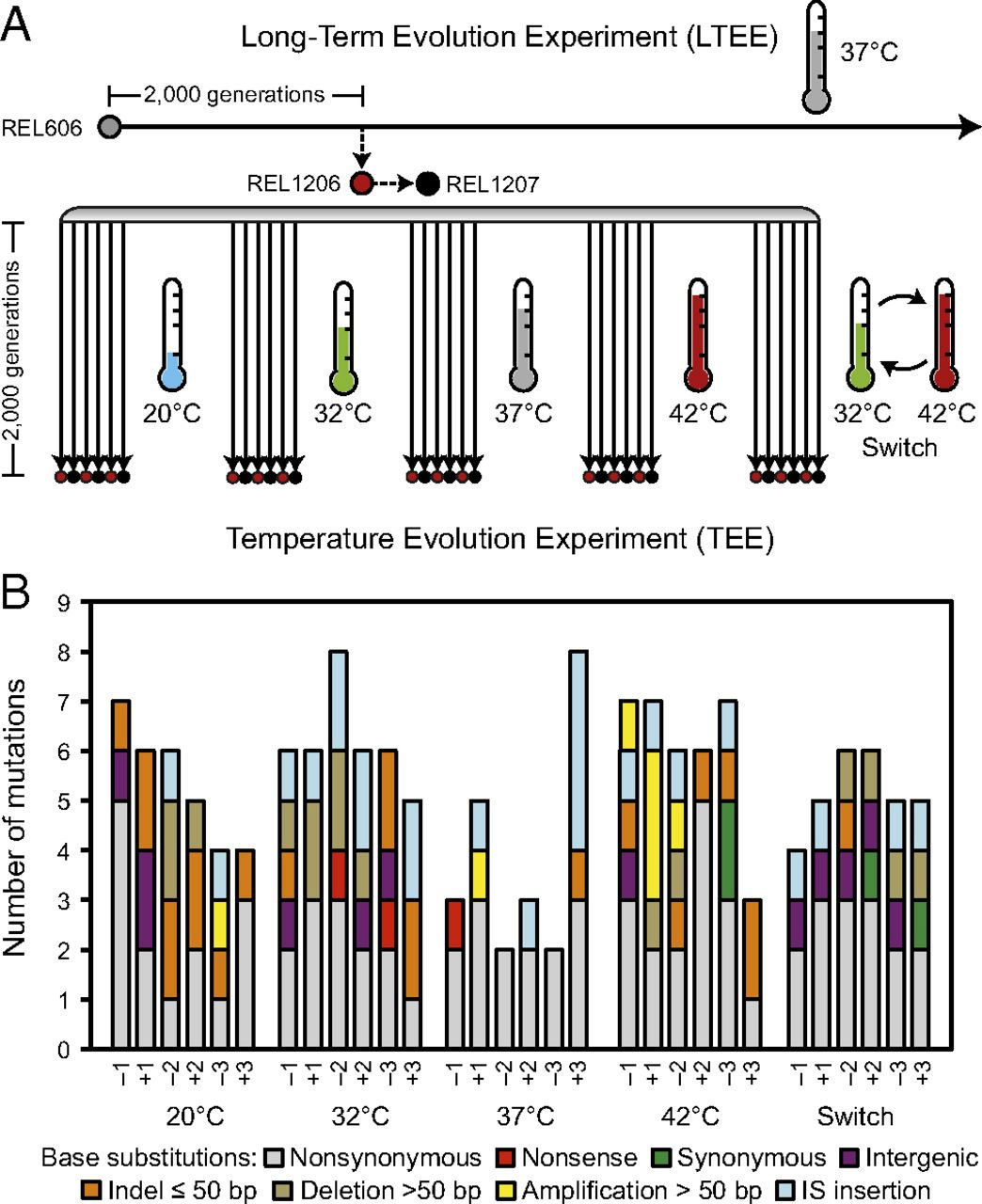
Рис. 2. Геномные изменения в кишечных палочках Escherichia coli, эволюционировавших при разных температурных режимах. А — общая схема эксперимента. Линия E. coli REL1206 была изолирована из долговременного эволюционного эксперимента после 2000 поколений эволюции при 37°C. В описываемом эксперименте шесть популяций эволюционировали в течение 2000 поколений при пяти разных температурных режимах. Три популяции в каждом из пяти вариантов опыта начинались с клона REL1206, неспособного перерабатывать арабинозу (обозначенные как −1, −2, −3) и три — с клона REL1207, способного к этому (+1, +2, +3). Эти клоны отличались одной заменой нуклеотида, которая не влияла на их приспособленность. В — распределение всех полученных в ходе эксперимента 159 мутаций по популяциям и типам генетических изменений. Рисунок из обсуждаемой статьи в PNAS
Все популяции бактерий за время эксперимента повысили скорость роста, то есть их приспособленность к среде возросла. Причем обычно эволюционировавшие бактерии оказывались более успешными по сравнению с предковыми не только в той среде, в которой их выращивали, но и на других. Но такое происходило далеко не всегда — например, популяции из 20, 32- и 37-градусных сред при 42°C росли хуже, чем предки. Впрочем, эти результаты были опубликованы уже давно (см. A. F. Bennett, R. E. Lenski, J. E. Mittler, 1992. Evolutionary adaptation to temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment; J. A. Mongold, A. F. Bennett, R. E. Lenski, 1996. Evolutionary adaptation to temperature. IV. Adaptation of Escherichia coli at a niche boundary). А в свежей работе авторы разбирались в генетической «подноготной» температурных адаптаций. Для этого они полностью отсеквенировали ДНК у представителей каждой популяции из конечного, 2000-го поколения.
Суммарно за все время эксперимента все исследуемые линии накопили 159 мутаций (напомним, что в эксперименте используется популяция E. coli, размножающаяся бесполым путем, поэтому эволюционные процессы изучают по накоплению мутаций) , при этом каждая отдельная линия накопила от двух до восьми мутаций — что интересно, только одна из этих «крайних» популяций росла не при оптимальных 37 градусах, а при «умеренных» 32-х. Но в среднем число мутаций, накопленных 37-градусными бактериями, значимо меньше, чем у всех остальных (но только если рассматривать их всех вместе; при попарном сравнении вариантов эксперимента различия в числе накопленных мутаций оказываются незначимыми — возможно, по причине малого числа популяций в каждом из вариантов опыта). Такие результаты вполне ожидаемы — ведь бактерии росли при оптимальной для них температуре, в которой, к тому же, в течение еще 2000 поколений развивались общие предки экспериментальных линий (рис. 2А). Таким образом, бактерии уже оказывались весьма адаптированными к этим условиям и на них действовал скорее очищающий отбор — отсеивающий мутации, разрушающие уже существующие адаптации, — чем движущий, приводящий к формированию новых приспособлений. Обнаруженные мутации относились к разным типам: это были синонимичные и несинонимичные замены нуклеотидов (см. Мутации: нуклеотидные замены: типы), делеции (утраты участков ДНК) и т. д. Их распределение по типам и отдельным популяциям приведено на рисунке 2В. Три из обнаруженных мутаций могут приводить к гипермутабильности — повышенному темпу мутирования. Такие мутации часто приобретаются в эволюционных экспериментах на бактериях, так как повышают шансы на появление благоприятных мутаций. Однако в данном случае популяции, несущие потенциальные гены-мутаторы, не показали сколько-нибудь значимого превышения числа полученных мутаций над ожидаемым.
Далее ученые оценили сходство мутаций, появившихся в разных популяциях, рассчитав коэффициент сходства Дайса, также известный как коэффициент Сёренсена (S = 2 (X∩Y)/(X + Y), где X и Y — число мутировавших генов в первой и второй сравниваемой группе, а X∩Y — число генов, мутировавших в обеих группах). При этом учитывали только те мутации, которые можно было точно отнести к конкретному гену (например, не учитывались все крупные делеции). Во-первых, коэффициент сходства для популяций внутри каждого варианта опыта оказался существенно больше, чем между разными вариантами: 0,168 против 0,042. При этом статистически незначимой оказалась только разница между линией, культивируемой при 42°C, и культивируемой при переменной температуре (рис. 3А). Во-вторых, удалось выявить специфичные мутации, значимо чаще возникающие (а иногда — и вообще уникальные) для отдельного температурного режима. Всего таковых оказалось четыре — только у переменного температурного режима не оказалось специфичного гена (рис. 3В).
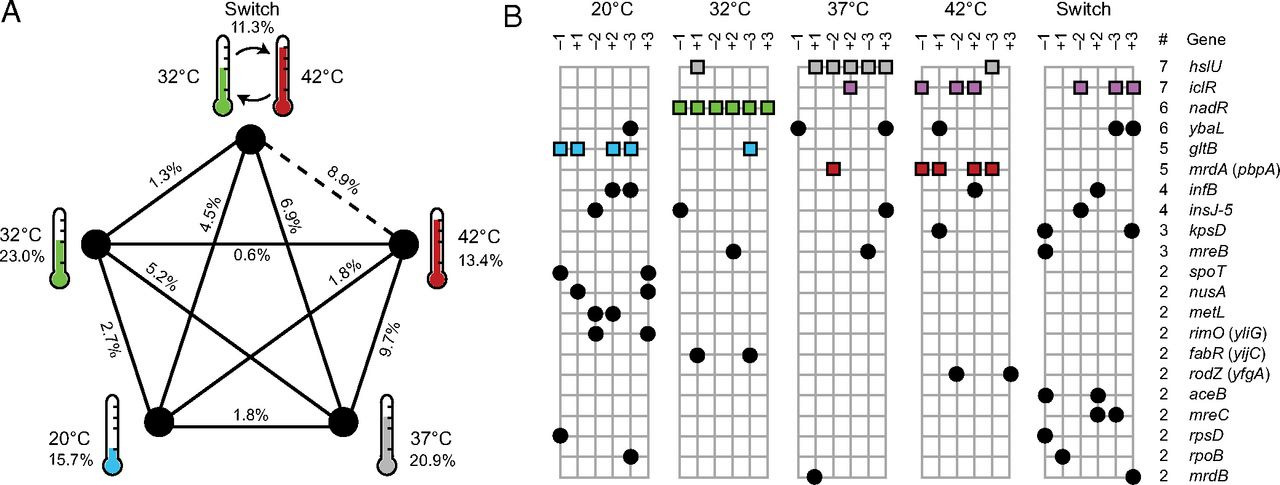
Рис. 3. А — схема сходства состава мутаций, приобретенных бактериями при разных температурных режимах. Проценты возле названия варианта опыта показывают коэффициент Дайса для всех шести участвовавших в нем популяций. Проценты возле соединяющих варианты опыта линий показывают коэффициент Дайса для сравнения двух температурных режимов. Только различия между популяциями, развивавшимися при 42°C и при переменном режиме (чередование температур 32°C и 42°C) оказались незначимыми. В — гены, мутировавшие хотя бы дважды во всех тридцати экспериментальных популяциях. Пять генов, отмеченных цветными квадратиками, значимо чаще встречаются внутри одной или двух линий по сравнению с предположением, что они распределялись между популяциями случайно. Рисунок из обсуждаемой статьи в PNAS
Но, по-видимому, адаптации не сводятся к этим четырем мутациям. Так как часть других мутировавших генов также показывает тенденцию к агрегации в одном из вариантов опыта. Что интересно, все гены, «специфичные» и подозреваемые в этом, относятся к нескольким функциональным группам: они влияют на размер и форму клеток (mrdA, hslU), регулируют метаболические пути, связанные с переработкой питательных веществ (nadR, iclR), и меняют активность метаболического фермента (gltB) (см. рис. 3В). Также среди «холодных» популяций есть тенденция к мутированию генов, связанных с транскрипцией и трансляцией. Таким образом, перед нами типичный пример комплексной адаптации, задействующей разные механизмы — и потому потенциально многовариантной. К счастью, современные молекулярно-генетичские методы позволяют вычленять отдельные гены и признаки, участвующие в приспособлении к тому или иному фактору среды, хотя еще несколько десятков лет назад едва ли не все эволюционисты считали, что распутать этот клубок невозможно и точные механизмы таких сложных адаптаций непознаваемы. Действительно, тогда можно было только сказать что-то вроде: «в ответе на отбор участвует множество генов, в каждой популяции — своих». Сейчас же мы можем сказать, какие именно это гены. И оказывается, что по крайней мере в некоторых случаях этот набор участвующих в адаптации генов оказывается воспроизводимым.
Интересные результаты принесло сравнение описываемого эксперимента с продолжающимся долговременным эволюционным экспериментом. Оказалось, что бактерии из ДЭЭ со временем накапливают мутации в тех же генах, что и адаптирующиеся к разным температурам! С одной стороны, им нужно адаптироваться не только к температуре — например, в среде обоих экспериментов не хватает глюкозы. С другой — где же тут специфичность адаптаций? Да, линия, культивируемая при 37°С, показывает наибольшее сходство с ДЭЭ, протекающем при той же температуре, — но ведь и остальные приобретают мутации, которые впоследствии оказываются полезными и при других температурах (рис. 4).

Рис. 4. Гены, служившие мишенью для отбора во время эксперимента по температурной адаптации, часто накапливали мутации и во время долговременного эволюционного эксперимента (ДЭЭ) при 37°C. На графике показано, как с течением долговременного эксперимента (секвенировали клоны на 5 000, 10 000 и 20 000 поколений; бактерии для температурного опыта были взяты из 2 000-го поколения) менялся индекс рекапитуляции, показывающий, какая доля генов, мутировавших при данном температурном режиме, «повторно» мутировала и во время ДЭЭ. Как и следовало ожидать, поначалу опыт с 37°C показывал наибольшее сходство с ДЭЭ, однако к 20 000-му поколению разница пропадала, что свидетельствует в пользу того, что мутации, возникшие при разных температурных режимах, полезны и при оптимальной температуре (напомню, что, кроме температуры, среда во всех вариантах была одинаковой). Рисунок из обсуждаемой статьи в PNAS
Авторы объясняют этот парадокс тем, что разные температурные режимы меняют относительную ценность разных мутаций, но редко делают полезную мутацию вредной — то есть, например, мутация, закрепившаяся в «холодных» линиях, в условиях низких температур оказывается более благоприятной, чем в оптимальных условиях, что повышает ее шанс на более раннее закрепление. В пользу этого говорит и прямая оценка адаптивной ценности отдельных мутаций. Исследователи вводили их в геном предковых бактерий и смотрели, как эти модифицированные организмы чувствуют себя при разных температурах. Ни для одной из мутаций не обнаружилось значимых различий по положительному эффекту на скорость роста при разных температурных режимах, хотя попарные сравнения выявили слабую тенденцию к тому, чтобы мутация была более благоприятной в том варианте опыта, где она возникла. Это подтверждает интерпретацию авторов: со сменой температуры все закрепившиеся мутации остаются благоприятными, но их относительная полезность меняется и естественный отбор работает именно с ними: оказавшиеся более благоприятными в данных условиях мутации закрепляются раньше.
Это исследование позволяет еще раз подчеркнуть, что эволюция может идти за счет вроде бы слабых, малозаметных различий. Не наличие эффективных и неэффективных путей, но небольшие различия в эффективности тех или иных приспособлений при разных температурах меняют вероятности их закрепления естественным отбором, что приводит к разным путям эволюции.
В конце статьи авторы подчеркивают, что, учитывая широкий диапазон возможностей для адаптации, их результаты показывают неожиданно высокую предсказуемость эволюции. И предполагают, что наличие таких специфических следов адаптации к конкретным условиям среды может помочь, например, в расшифровке эволюционной истории патогенных микробов, отслеживая историю перехода между разными видами, или даже индивидами хозяев (что поможет отследить динамику заболеваний), а также в судебной микробиологии, помогая выяснять обстоятельства гибели или историю перемещений отдельных предметов (см. R. E. Lenski, P. Keim, 2005. Population genetics of bacteria in a forensic context).
Источник: Daniel E. Deatherage, Jamie L. Kepner, Albert F. Bennett, Richard E. Lenski & Jeffrey E. Barrick. Specificity of genome evolution in experimental populations of Escherichia coli evolved at different temperatures // Proceedings of the National Academy of Sciences. 2017. V. 114. №10. P. 1904–1912. DOI: 10.1073/pnas.1616132114.
Сергей Лысенков
-
Сегодня просмотр «Элементов» начался с неприятного сюрприза. Все-таки переключили на новый дизайн. Дискуссии читать стало очень сложно, поскольку длинные комментарии с большой вложенностью вытягиваются в узкую и очень длинную полосу. Большие по объему статьи также сильно вытягиваются.
Есть возможность переключить на старый дизайн http://old.elementy.ru . Старая версия также немного испортилась. При просмотре в «Опера» вроде все нормально. При просмотре в «GооgleChrome» не работают «резиновые» строки. Так-что, пока есть такая возможность, буду использовать старую версию сайта. Дальше будет очень грустно.
Извините, что не по теме. -
"В конце статьи авторы подчеркивают, что, учитывая широкий диапазон возможностей для адаптации, их результаты показывают неожиданно высокую предсказуемость эволюции." - Пардон, никаких "широких возможностей" для адаптации в данном случае нет. "Широкие возможности" есть только в головах людей, свято исповедующих СТЭ. С фактором среды "изменение температуры" E. coli сталкиваются столько раз, сколько их носители, еще раз пардон, ходят "до ветру". Все уже давно преадаптировано - и метаболические пути, и активность фермента, и гены транскрипции и трансляции. Таким образом, перед нами нет и не может быть никакой "потенциально многовариантной комплексной адаптации", как нет ее, например, при адаптации людей к недостатку пищи, каковые вполне предсказуемы - уменьшение роста и веса и усиление эффективности метаболических путей, и активности ферментов. Никого не удивило сходство с бактериями? А жаль - должны были удивиться вместе с авторами статьи. "Потенциальная многовариантность" была бы, если бы где-нибудь в джунглях Амазонки, где голодовка - обычное периодическое явление, находили бы племена индейцев, питающихся опилками. А что? У всех людей есть бродильные бактерии в прямой кишке.
-
Есть 2 вопроса.
Вопрос первый, про делеции. Если мои познания в генетике не слишком примитивные, то я знаю о таких кусках генома, как "команда на старт транскрипции" и "команда на конец транскрипции" (или как там это называется). А может при мутации пропасть одна из этих 2 команд? Или любой код реального организма защищен от столь явных сбоев простым дублированием?
Второй, про "чередование умеренных и экстремальных условий". Оно происходило буквально каждое поколение или речь про десятки или сотни?

Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки -
 14.10.2018Успех адаптации мух Drosophila melanogaster к избытку соли в корме зависит от дрожжей определенного видаСергей Ивницкий • Журнал общей биологии • №5, 2018
14.10.2018Успех адаптации мух Drosophila melanogaster к избытку соли в корме зависит от дрожжей определенного видаСергей Ивницкий • Журнал общей биологии • №5, 2018
-
 06.03.2018Мухи, приспособленные к неоднородной среде, получают эволюционное преимуществоЕ. Яковлева, А. Горшкова, Е. Фетисова • Журнал общей биологии • №1, 2018
06.03.2018Мухи, приспособленные к неоднородной среде, получают эволюционное преимуществоЕ. Яковлева, А. Горшкова, Е. Фетисова • Журнал общей биологии • №1, 2018
-
 23.10.2017В долгосрочном эксперименте Ричарда Ленски из одинаковых бактерий сформировались полиморфные сообществаАлександр Марков • Новости науки
23.10.2017В долгосрочном эксперименте Ричарда Ленски из одинаковых бактерий сформировались полиморфные сообществаАлександр Марков • Новости науки
-
 08.05.2017Эксперимент на улитках подтвердил классическую идею о «двойной цене самцов»Александр Марков • Новости науки
08.05.2017Эксперимент на улитках подтвердил классическую идею о «двойной цене самцов»Александр Марков • Новости науки
-
 16.03.2017Способы адаптации бактерий к разным температурам оказались предсказуемымиСергей Лысенков • Новости науки
16.03.2017Способы адаптации бактерий к разным температурам оказались предсказуемымиСергей Лысенков • Новости науки
-
 03.11.2016В меню плодовой мушки может появиться... соленый огурец!Сергей Ивницкий • Журнал общей биологии • №4, 2016
03.11.2016В меню плодовой мушки может появиться... соленый огурец!Сергей Ивницкий • Журнал общей биологии • №4, 2016
-
 13.09.2016Эволюционный эксперимент показал, где и как появляются наиболее приспособленные особиЕлена Наймарк • Новости науки
13.09.2016Эволюционный эксперимент показал, где и как появляются наиболее приспособленные особиЕлена Наймарк • Новости науки
-
 09.12.2015Неадаптивная фенотипическая пластичность затрудняет видообразованиеАлександр Марков • Новости науки
09.12.2015Неадаптивная фенотипическая пластичность затрудняет видообразованиеАлександр Марков • Новости науки
-
 07.09.2015Неадаптивная пластичность ускоряет адаптивную эволюциюАлександр Марков • Новости науки
07.09.2015Неадаптивная пластичность ускоряет адаптивную эволюциюАлександр Марков • Новости науки
Последние новости







Рис. 1. Колонии Escherichia coli в чашке Петри на среде из агара, содержащего арабинозу. Розовые — способные перерабатывать арабинозу (Ara+), красные — не способные (Ara–). Фото с сайта telliamedrevisited.wordpress.com