Два независимых исследования подтвердили глобальное ослабление синапсов во время сна
Согласно «гипотезе синаптического гомеостаза», сон необходим животным, потому что усвоение нового опыта во время бодрствования происходит в основном за счет усиления, а не ослабления синаптической проводимости. Общее нарастание проводимости снижает работоспособность нервной системы, которая поэтому должна регулярно переходить в режим «офлайн», отключаясь от внешних сигналов. Это позволяет избирательно ослабить перевозбужденные синапсы, аккуратно отделяя при этом важную информацию от неважной. Двум исследовательским коллективам из США удалось получить новые независимые подтверждения этой гипотезы. Одна работа основана на трехмерной электронной микроскопии и прямом измерении синапсов, другая — на сравнении количества рецепторов и других белков в синапсах до и после сна. Оказалось, что у мышей во сне достоверно уменьшается и размер синапсов, и количество рецепторов, ответственных за прием возбуждающих сигналов.
Вопрос о том, почему животным необходим сон, до сих пор не решен окончательно, несмотря на обилие фактов и гипотез (см. ссылки в конце новости). По-видимому, должна быть какая-то общая важная причина, по которой самые разные животные — от нематод и насекомых до ящериц и людей — должны регулярно впадать в заторможенное, уязвимое состояние, почти полностью отключаясь от всех сигналов, приходящих из внешнего мира.
Красивое объяснение предлагает «гипотеза синаптического гомеостаза», впервые сформулированная около 15 лет назад нейробиологами из Висконсинского университета в Мэдисоне (см.: G. Tononi, C. Cirelli, 2014. Sleep and the Price of Plasticity: From Synaptic and Cellular Homeostasis to Memory Consolidation and Integration). «Элементы» уже рассказывали об этой гипотезе и ее эмпирических подтверждениях, полученных на дрозофиле, в новости Во время сна количество синапсов в мозге уменьшается («Элементы», 06.07.2011).
Суть идеи в том, что во время бодрствования животное постоянно усваивает новый опыт, а это сопряжено с ростом синаптической проводимости (см. Синаптическая пластичность). Как правило, чтобы что-то запомнить хотя бы ненадолго, необходимо усилить проводимость определенных синапсов: этот принцип лежит в основе памяти и обучения (см.: Нейроны соревнуются за право участия в формировании рефлексов, «Элементы», 26.04.2007). В результате общий уровень синаптической проводимости в мозге неуклонно нарастает в течение всего времени, пока животное бодрствует и активно воспринимает внешние сигналы. Ясно, что это не может продолжаться до бесконечности. Разбухшие синапсы и перевозбужденные нейроны будут потреблять всё больше энергии, а способность нервной системы к дальнейшему обучению будет снижаться.
Соответственно, сон необходим для того, чтобы вернуть нервную систему в рабочее состояние, снизив общий уровень синаптической проводимости. Заодно можно отделить в накопленном за день опыте главное от второстепенного, избирательно ослабив одни синапсы и сохранив (или даже усилив) некоторые другие. Этот гипотетический процесс называют «умным забыванием» (smart forgetting) (о консолидации памяти во сне рассказано в новостях Во время фазы медленного сна активно закрепляются новые знания, «Элементы», 21.03.2007; Мозг во сне «проигрывает» дневные события с семикратным ускорением, «Элементы», 19.11.2007).
По сути дела, гипотеза синаптического гомеостаза предполагает, что потребность во сне — это следствие неустранимого конструктивного дефекта нервной системы животных! Нервная система не может долго обрабатывать входящую информацию, не «перегреваясь», потому что усвоение новой информации идет в основном за счет потенциации (усиления) имеющихся синапсов и появления новых, а не их депрессии (ослабления) или уничтожения.
В двух статьях, опубликованных в свежем выпуске журнала Science, приводятся новые весьма убедительные подтверждения гипотезы синаптического гомеостаза.
В первом исследовании (de Vivo et al., 2017), выполненном нейробиологами из Висконсинского университета (в том числе авторами гипотезы), при помощи трехмерной электронной микроскопии (см. Serial block-face scanning electron microscopy) удалось напрямую сравнить размеры синапсов в мозге мышей до и после сна.
Размер синапса довольно строго коррелирует с его проводимостью. Поэтому гипотеза предсказывает, что во время сна синапсы в целом должны уменьшаться. Уменьшение, однако, должно быть избирательным: важные впечатления должны как-то отделяться от неважных и запоминаться надолго.
Исследователи сравнивали нейроны моторной и соматосенсорной коры у хорошо выспавшихся и у долго не спавших мышей, причем последние делились на две группы: первые не спали по собственной инициативе, потому что время суток было неподходящее (ночное, ведь мыши активны по ночам), а вторым не давали спать днем, развлекая новыми предметами обстановки.
На основе послойных электронных микрофотографий были получены трехмерные модели дендритов со всеми их шипиками и синапсами (рис. 2). В общей сложности было реконструировано 6920 синапсов 12 мышей (по 4 мыши из каждой группы), причем все исследованные образцы мозговой ткани происходили из одних и тех же двух крохотных участков коры. В качестве меры синаптической проводимости использовалась площадь контакта дендритного шипика с аксонной терминалью (ASI, axon-spine interface). Известно, что этот показатель довольно точно отражает силу синапса.

Рис. 2. Объемные реконструкции дендритов и синапсов, полученные при помощи трехмерной электронной микроскопии. Слева вверху показаны участки моторной и соматосенсорной коры, из второго слоя которых брались образцы мозговой ткани. C — реконструкция четырех дендритов с шипиками в изученном образце, D — другие примеры трехмерных реконструкций дендритов, E — пример двумерного снимка, на котором видна часть дендритного шипика (выделена желтым) и часть аксонной терминали (выделена зеленым) с синаптическими пузырьками, в которых содержится нейромедиатор; поверхность контакта выделена красным. Из множества таких послойных фотографий делаются трехмерные модели нейронов. F — реконструкция синапса. dendrite — дендрит, axon — аксон, spine head — головка дендритного шипика, ASI (axon-spine interface) — поверхность контакта шипика с аксонной терминалью; по величине ASI можно судить о силе синапса. Изображение из обсуждаемой статьи de Vivo et al., 2017
Оказалось, что сон сокращает ASI в среднем на 18–19% по сравнению с бодрствованием (как добровольным, так и вынужденным). При этом диаметр дендритов остается прежним: это значит, дело не в общем сжатии нейронов а именно в уменьшении синапсов. Таким образом, подтвердилось первое предсказание гипотезы синаптического гомеостаза: синапсы действительно уменьшаются во сне.
Статистический анализ полученных данных подтвердил и второе предсказание — об избирательности уменьшения синапсов. Пропорциональному уменьшению подвергается большинство синапсов, но не все. При этом вероятность того, что синапс будет ослаблен во сне, связана обратной зависимостью с размером синапса. Наибольшие шансы остаться неуменьшенными имеют 20% самых больших синапсов. Возможно, в этих синапсах закодирована важная информация и самые сильные впечатления, которые не стоит забывать. Кроме того, авторы заметили, что уменьшение синапсов во сне сильнее выражено у дендритов с редко расположенными шипиками по сравнению с дендритами, на которых шипиков больше. Еще одна закономерность состоит в том, что пропорциональному уменьшению чаще подвергаются синапсы, в которых много эндосом.
Расчеты, основанные на другом косвенном показателе силы синапса — объеме головки дендритного шипика, — совпали с результатами, полученными на основе показателя ASI. Головки шипиков тоже уменьшаются во сне.
Второе исследование (G. H. Diering et al., 2017), проведенное нейробиологами и биохимиками из Университета Джонса Хопкинса в Балтиморе, тоже проводилось на выспавшихся и невыспавшихся мышах и тоже в общем и целом подтвердило гипотезу синаптического гомеостаза, хотя основано оно было не на трехмерной электронной микроскопии, а на протеомике.
Исследователи выделили (см.: Cell fractionation) из переднего мозга мышей фракцию постсинаптических уплотнений (Postsynaptic density, PSD) — белковых комплексов, связанных с постсинаптическими мембранами, — и сравнили количество разных белков в PSD до и после сна.
Оказалось, что во время сна в постсинаптических уплотнениях достоверно снижается содержание ряда ключевых белков, связанных с быстрой передачей нервных импульсов при помощи главного возбуждающего нейромедиатора головного мозга — глутамата. В частности, уменьшается количество белков GRIA1 и GRIA2, которые являются субъединицами (составными частями) ионотропных глутаматных рецепторов. Уменьшение количества этих рецепторов в PSD снижает чувствительность постсинаптической мембраны к глутамату, то есть непосредственно ведет к ослаблению синапса.
К таким же выводам привели прямые наблюдения при помощи двухфотонного микроскопа за нейронами живых трансгенных мышей, у которых белок GRIA1 был помечен флуоресцентной меткой. Наблюдения подтвердили, что во время сна количество GRIA1 в синапсах снижается.
Как и в первом исследовании, ослабление синапсов во сне оказалось избирательным. Авторы отметили более сильное снижение GRIA1 в тех синапсах, где во время бодрствования этого белка было больше. То есть, в отличие от первого исследования, которое показало избирательное ослабление слабых синапсов, второе исследование вроде бы показало избирательное ослабление сильных синапсов. В чем причина этого расхождения, является ли оно следствием ошибки или демонстрирует разные грани сложного процесса перестройки синапсов во сне — покажут дальнейшие исследования.
Дополнительные эксперименты позволили частично расшифровать молекулярный механизм, ответственный за ослабление синапсов во время сна. Выяснилось, что ключевую роль в этом играет белок Homer1a (короткий сплайс-вариант белка Homer1), который во время сна поступает в дендритные шипики. Homer1a разрушает связь между белками mGluR и IP3R, что через ряд промежуточных шагов приводит к уменьшению количества ионотропных глутаматных рецепторов на постсинаптической мембране. Уже было известно, что Homer1a участвует в гомеостатическом снижении чувствительности постсинаптических мембран, на которые приходит слишком много возбуждающих сигналов. Теперь стало ясно, что этот механизм участвует также и в ослаблении перевозбужденных синапсов во время сна.
В заключительной серии экспериментов авторы показали, что поступление белка Homer1a в дендритные шипики контролируется нейромодуляторами норадреналином и аденозином. Во время бодрствования в мозге повышен уровень норадреналина, что препятствует поступлению Homer1a в синапсы, хотя матричная РНК этого белка активно производится в работающих нейронах. Повышение уровня аденозина соотносится с сонливостью и способствует транспорту белка Homer1a, накопившегося в активно работающих нейронах во время бодрствования, в дендритные шипики.
Таким образом, оба исследования убедительно подтвердили гипотезу синаптического гомеостаза. Судя по всему, во время сна действительно происходит глобальное снижение уровня синаптической проводимости в мозге. Оба исследования показали, что ослабление синапсов имеет избирательный характер. Очевидно, что эта избирательность должна быть как-то связана с консолидацией памяти и с отделением важного от второстепенного, то есть с ростом отношения сигнала к шуму (Signal-to-noise ratio). Очевидно также, что имеющихся данных пока недостаточно, чтобы понять, как устроена и чем регулируется эта избирательность.
Источники:
1) Luisa de Vivo, Michele Bellesi, William Marshall, Eric A. Bushong, Mark H. Ellisman, Giulio Tononi, Chiara Cirelli. Ultrastructural evidence for synaptic scaling across the wake/sleep cycle // Science. 2017. V. 355. P. 507–510.
2) Graham H. Diering, Raja S. Nirujogi, Richard H. Roth, Paul F. Worley, Akhilesh Pandey, Richard L. Huganir. Homer1a drives homeostatic scaling-down of excitatory synapses during sleep // Science. 2017. V. 355. P. 511–515.
См. также:
1) Во время сна количество синапсов в мозге уменьшается, «Элементы», 06.07.2011.
2) Мозг во сне «проигрывает» дневные события с семикратным ускорением, «Элементы», 19.11.2007.
3) Во время фазы медленного сна активно закрепляются новые знания, «Элементы», 21.03.2007.
4) Мухи устают от общения, «Элементы», 02.10.2006.
5) Нематоды во сне перестраивают свою нервную систему, «Элементы», 16.01.2008.
6) Елена Наймарк. Наука во власти сна, «Что нового в науке и технике» №7–8, 2005.
7) Анна Хоружая. Почему человек спит?, «Популярная механика» №8, 2015.
-
Года два назад встроил периоды регуляризации в свои экспериментальные нейросети, и оно хорошо работало. По моему мнению, основанному только на экспериментах с искусственными нейросетями, это не неустранимый дефект, а фича такая, открывающая новые возможности для консолидации полезного опыта. На стадии обучения увеличить количество новых синапсов, а потом в процессе избирательной регуляризации вычистить их до исходного количества на много на много эффективнее, чем сразу запустить оба процесса обучения и и регуляризации имея дело с постоянным количеством синапсов.
Собственно приятно, что исследования на мышах подтвердили смутные гипотезы.-
-
Я как-то даже хотел попробовать организовать встречу с вами на тему "почему эволюция идёт с ускорением с точки зрения математики", с этой точки зрения это близкородственные задачи, но потом подумал, что без экспирементального подтверждения к вам такие умники десятками ломятся. :)
-
Какие-то условные параметры вроде "количество изобретений в год" явно выросло, если сравнить 2 периода:
1) 100-700 гг. нашей эры;
2) 1982-2002 гг. нашей эры.
А почему так произошло - есть несколько причин.-
Речь не об открытиях и сингулярности. То что на меметику распостраняются похожие законы логично, но очень трудно доказуемо формально, а кроме того эволюция живого от меметической революции очень сильно отличается метапараметрами в большинстве случаев.
Я имею в виду строго математическую трактовку того почему с ускорением идёт эволюция геномов живых существ. И как раз из-за того, что эту задачу можно строго формализовать и аргументы не сводятся к убедительности рассуждений эта задача становится гораздо более трудной.
-
-
-
-
-
Я бы немного подправил Ваши наблюдения с учетом своего опыта работы с искусственными нейронными сетями. В случае компьютерной программы нейронная сеть дизайнируется не методом самосборки, а «творцом» (всякую жесть типа генетических алгоритмов для перебора коммутаций в нейонных сетей в расчет не берем). Так вот, многие моменты организации природных НН (включая и сон/регуляризацию) будут оптимальны *только* с учетом сложности генных сетей необходимых для кодирования их самосборки.
-
Согласен полностью. Поэтому мои сети со значительной самосборкой, они сами добавляют и удаляют синапсы и нейроны и и то и другое делают неравномерно и как раз с перебором коммутаций, не случайным, конечно, а с очень сильной эвристикой. Этого оказывается достаточно чтобы эффект проявился. А сейчас я к ним планирую прикрутить разделение неронов на семейства и генетический алгоритм настраивающий метапараметры семейств, надеюсь ещё больше увеличить выигрыш.
Но не сложность организации сети первична. Первична сложность задачи с которой плохо справляются сети без всех этих наворотов. Если мою сеть натравить на ирисы или вообще любую задачу с более или мене гладким адаптивным рельефом выигрыша никакого, я прверял. Неравномерная регуляризация выстреливает если в исходных данных есть паттерны, обобщение которых значительно повышает результат - именно это она помогает осуществлять.-
У Вас, по-ходу, серьезные задачи, коллега. Очень интересно. Я вот до вашего поста был абсолютно уверен, что современный хард еще не способен подымать такое «многомерное обучение»… А на чем считаете, если не секрет? И сколько миллионов нейрнов/синапсов/сэмплов приходиться на Вашу среднюю НН? И правильно ли я понимаю, что “1D” обучение и оценка дизайна НН у Вас все же на backpropagation/deep-belief а не на вариации рандом чего-то там?
З.Ы. У меня, всякие молекулярные связывания со сложностью по формуле выше 1-2/1К/10 и это все весьма туго идет на 128-cpus (тредов не камней) Xeon E7-8867 от Dell.
З.Ы.Ы. Не буду спорить - природные НН действительно решают самую сложную из всех возможных задач - выжить. :)-
Задачи у меня синтетические, сложные для сети, но малой размерности. Так что миллионами я не оперирую. Всё на проце, пока изучаю возможности алгоритмов, а не машинное зрение на них. Ну и алгоритм специфический - много нужно писать руками, на проце проще.
Но тем не менее, такие алгоритмы железо современное вполне потянет, нужно только знать что именно делать. Неравномерная регуляризация тяжёлая, но весит как O(n) и запускается раз в эпоху. Так что накладных расходов на неё совсем немного. Эвристика от перекоммутации запускается часто, но весит при этом только O(log(n)) так что не всё так страшно. Ну и допрасходы на то что сама сеть оказывается сложнее на два обращения в массив по индексу, чем нормальная, потому что требуется поддерживать сеть с почти произвольным коннектомом, вот тут самое большое неудобство. А оценка дизайна - вообще копейки.
Алгоритм получается заметно сложнее того что обычно гоняют на кластерах титанов, но при миллионах всё равно должно получаться приемлемо. Когда на видюху рассчёты переведу померяю точно.
-
-
-


Предположим, что 20% музыкантов симфонического оркестра вносят 80% вклада в звучание симфонии. Т.е. если убрать 80% тех, то еще можно получить удовольствие, а вот если замолчат 20% этих, то публика разойдется. Допустим, этим 20% платят больше, и отдыхают они тоже больше (потому что они ценнее и устают больше, поскольку выкладываются сильнее). Концерт=бодрствование. Сравнения - до концерта (=после сна) и по завершении (=перед сном).
Первая группа исследователей обнаружила, что 80% музыкантов после концерта получают вознаграждение на 20% меньше, чем "ценные" 20% оркестрантов (которым заплатят одинаково - как за концерт, так и за отдых). Вторая группа показала, что эти 20% музыкантов между концертами отдыхают больше. Нет противоречия.
Как-то так примерно... Или нет? )))
-
Вот это Вы завернули! Хотя мне тема симфонического оркестра и весьма близка, это Ваше сравнение (вероятно, имеющее целью упростить понимание описанной в статье картины?) мне куда менее понятно, чем сама статья.
Даже вызвало сомнение - а понял ли я вообще что-либо в статье? Настолько мало написанное Вами коррелирует с тем, что я себе понапредставлял по прочтении её)
Последние новости










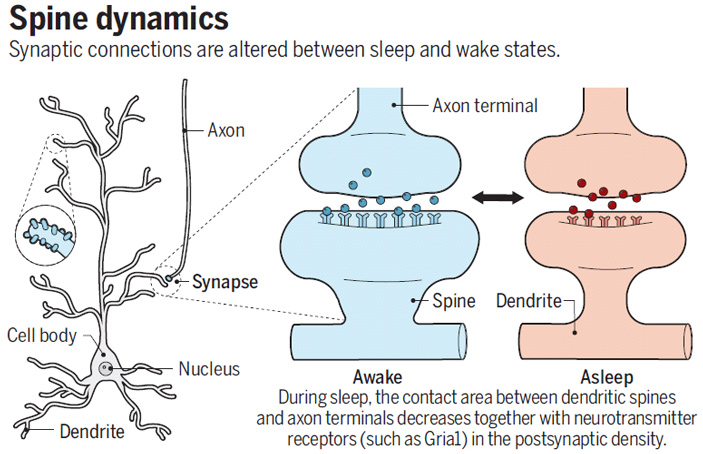
Рис. 1. Рисунок, иллюстрирующий главный вывод обеих обсуждаемых работ: во время сна происходит ослабление синапсов. Слева — схематическое изображение нейрона (Dendrite — дендрит, Nucleus — ядро, Cell body — тело нейрона, Synapse — синапс, Axon — аксон). Справа — синапс после бодрствования (Awake) и после сна (Asleep). Во время сна сокращается площадь контакта аксонной терминали (Axon terminal) с дендритным шипиком (Spine), а также количество глутаматных рецепторов на постсинаптической мембране. Рисунок из синопсиса к обсуждаемым статьям в Science (L. Acsády, K. D. Harris. Synaptic scaling in sleep).