Правильно выбирая друзей, можно спастись от конкурентов
При помощи математического моделирования и анализа реальных симбиотических комплексов растений и насекомых-опылителей удалось показать, что взаимовыгодное сотрудничество между видами (мутуализм) может способствовать снижению конкуренции и росту числа видов в сообществе, но только в том случае, если сеть мутуалистических связей обладает высокой степенью «вложенности». Это означает, что виды-специалисты, имеющие мало «друзей», должны кооперироваться с видами-генералистами (у которых «друзей» много), и наоборот. Что касается отношений «хищник–жертва», то с ними всё наоборот: чем выше степень «вложенности» в сети таких связей, тем острее межвидовая конкуренция и тем меньше видов может ужиться в одном сообществе.
В условиях быстро развивающегося экологического кризиса вопрос о факторах, влияющих на биологическое разнообразие, становится всё более актуальным. Несмотря на значительные успехи, достигнутые экологами на этом поприще, многое еще остается неясным. Одним из наименее разработанных является вопрос о влиянии структуры экологических связей на видовое богатство, то есть на число видов, которые могут жить вместе в одном сообществе. Дело затрудняется тем, что связи между видами в экосистеме могут быть чрезвычайно сложными и разнообразными. Поэтому начинать приходится с заведомо упрощенных моделей.
Испанские экологи попытались разработать теоретическую базу для изучения влияния мутуализма — взаимовыгодного сотрудничества между видами — на свойства экосистемы. Прототипом для модели послужили системы мутуалистических взаимоотношений между растениями и животными — опылителями или распространителями семян. Однако применимость полученной модели может оказаться гораздо шире.

Модельное сообщество состоит из двух групп видов — «растений» и «насекомых-опылителей». Все растения конкурируют друг с другом за ресурсы (например, за место под солнцем). Насекомые тоже конкурируют друг с другом. Между растениями и насекомыми могут возникать взаимовыгодные отношения. Судьба каждого вида в модели зависит от соотношения вреда, приносимого врагами-конкурентами, и пользы, приносимой друзьями-мутуалистами. На выходе оценивается максимальное число видов растений и насекомых, которые могут устойчиво сосуществовать при данном наборе входных параметров. Подробное описание модели дано в дополнительных материалах к статье.
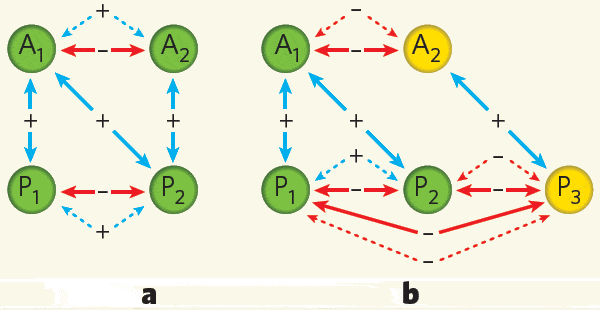
В основе модели лежит вполне разумное предположение о том, что мутуалистические взаимоотношения между растениями и насекомыми могут сглаживать негативный эффект конкуренции внутри каждой из двух групп. Особенно четко этот эффект проявляется в том случае, если каждое растение опыляется всеми видами насекомых. При этом для каждого вида растений остальные растения оказываются не только врагами-конкурентами, но и «косвенными друзьями», потому что они подкармливают своим нектаром насекомых, опыляющих данное растение.
В реальности, конечно, так не бывает: каждое растение опыляется не всеми, а только некоторыми видами насекомых, а каждый вид насекомых опыляет не все, а только некоторые виды растений. Анализ модели показал, что суммарное число видов, которые могут ужиться в сообществе, зависит не только от числа и силы мутуалистических связей, но и от их структуры, то есть от того, кто с кем «дружит». Ключевую роль играет такая характеристика структуры связей, как «вложенность» (nestedness). Высокая степень «вложенности» означает, что виды-специалисты, имеющие малое число «друзей», поддерживают взаимовыгодные отношения в основном с видами-генералистами, у которых «друзей» много, и наоборот. Если степень «вложенности» высока, мутуализм резко ослабляет конкуренцию, что ведет к росту максимально возможного числа видов в сообществе. При низкой «вложенности», то есть в том случае, когда специалисты дружат со специалистами, мутуализм не ведет к снижению конкуренции или даже усиливает ее. Сообщество с такой структурой может вместить гораздо меньше видов.
Для проверки этих выводов авторы проанализировали имеющиеся в литературе данные по 56 реальным сетям мутуалистических связей в сообществах растений и животных — опылителей и распространителей семян. Подставляя реальные цифры в модельные формулы, они показали, что закономерность, выявленная в модели, действительно имеет место в природе. Чем выше степень «вложенности» в сети мутуалистических взаимодействий, тем больше видов, при прочих равных условиях, входит в эту сеть.
Большинство природных сетей мутуалистических взаимодействий, как выяснилось, имеют высокую степень «вложенности». Реальные сообщества словно специально организованы таким образом, чтобы максимально усилить положительный эффект, оказываемый мутуализмом на видовое разнообразие. Это может показаться странным. Виды, входящие в сообщество, как будто знают, с кем им следует «дружить», чтобы в сообщество поместилось побольше видов и суммарное разнообразие достигло максимума.
Авторы предложили этому явлению простое и изящное объяснение. Когда какой-нибудь вид внедряется в уже существующее сообщество, он, разумеется, не думает об общем благе. Однако вероятность того, что новый вид сможет закрепиться в сообществе, стать его постоянным членом, зависит от силы конкурентного давления, которое он будет испытывать. Чем сильнее конкуренты будут давить «новичка», тем меньше шансов, что он сумеет выжить в новом окружении. А сила конкурентного давления, в свою очередь, зависит от выбора «друзей». Новичок, скорее всего, будет вступать в сообщество в роли специалиста. Например, если это растение, маловероятно, что все имеющиеся в сообществе насекомые сразу кинутся его опылять: это было бы слишком большим «везением» для новичка. Скорее всего, найдутся лишь единичные виды опылителей, которые им заинтересуются. И вот тут-то решающее значение будет иметь то, какие именно это будут виды — генералисты или специалисты.
В первом случае растение-«новичок» сразу станет «косвенным другом» многих других растений, и конкурентное давление будет ослаблено. Во втором случае всё будет наоборот, и «новичок», скорее всего, не приживется в этом сообществе. Следовательно, видовой состав сообщества будет пополняться преимущественно за счет видов-специалистов, которым удалось подружиться с уже живущими в сообществе видами-генералистами. Такой механизм пополнения сообщества должен вести к поддержанию высокого уровня «вложенности» структуры связей.
Получается, что мутуализм в природе, как правило, способствует росту видового разнообразия, хотя в принципе он вовсе не обязан этого делать и теоретически может даже оказывать обратный эффект.
Анализ модели дал еще один любопытный результат. Оказалось, что, если некоторые из мутуалистических взаимодействий в системе вдруг усилятся, это может дать соответствующим видам большое преимущество в конкурентной борьбе со всеми остальными видами, у которых взаимовыгодные связи остались слабыми. В результате виды, связанные сильными взаимодействиями, быстро вытеснят всех своих конкурентов. Это означает, что резкое усиление мутуализма у одних организмов теоретически может приводить к вытеснению других и даже к их вымиранию.
Бывает ли такое на самом деле? Точного ответа пока нет. Не исключено, что подобные процессы сыграли какую-то роль в событиях конца мезозоя — начала кайнозоя. У появившихся в середине мелового периода покрытосеменных растений, по-видимому, сразу сложились прочные мутуалистические отношения с насекомыми — опылителями и млекопитающими — распространителями семян, тогда как динозавры, судя по некоторым косвенным данным, не смогли вступить во взаимовыгодные отношения с новыми доминантами растительного мира (см.: Butler et al., 2009). Не стало ли это одной из причин их упадка?
Если в разработанную авторами модель внести небольшие дополнения, она может быть использована для описания более сложных систем. Например, кроме растений и животных-опылителей в нее можно добавить растительноядных животных и хищников, питающихся опылителями. Основные выводы о влиянии структуры мутуалистических связей на видовое разнообразие сообщества при этом не изменяются. Что же касается связей «хищник–жертва», то их структура влияет на конкуренцию и разнообразие противоположным образом. Чем выше степень «вложенности» этих связей, тем острее конкуренция и тем ниже видовое разнообразие сообщества.
Новые виды жертв, вступая в сообщество, не заинтересованы в том, чтобы на них охотился хищник-генералист, потому что это приведет к обострению их конкуренции с другими жертвами. При наличии общего врага у двух видов жертв рост численности одного из видов будет вредить другому не только прямо (через конкуренцию за ресурсы), но и косвенно, через рост численности хищника. Это справедливо и для самих хищников: вступая в сообщество, им выгодно ограничить свою диету жертвами-«специалистами», то есть теми, кто не служит пищей большинству других хищников в этом сообществе. В этом случае другие хищники будут для новичка не столько конкурентами, сколько «косвенными друзьями», потому что они будут выедать конкурентов того вида жертв, которым питается новичок.
Таким образом, высокое видовое разнообразие в сообществе становится возможным в том случае, когда степень «вложенности» мутуалистических связей высокая, а связей «хищник–жертва» — низкая. Чтобы ослабить конкуренцию, всем конкурентам следует иметь как можно больше общих друзей и как можно меньше общих врагов (или жертв). Максимальное видовое разнообразие может быть достигнуто в том случае, если мутуалистические связи предельно генерализованы (каждый дружит со всеми, с кем только можно), а связи «хищник–жертва», наоборот, предельно специализированы (каждый вид хищников охотится только на один вид жертв).
Данная модель, по-видимому, в какой-то мере приложима не только к биологическим, но и к социальным системам. Например, есть основания полагать, что высокая степень «вложенности» партнерских (мутуалистических) отношений между компаниями (например, между производителями и заказчиками), препятствует образованию монополий и способствует мирному сосуществованию большого числа компаний в данной отрасли экономики.
Источники:
1) Ugo Bastolla, Miguel A. Fortuna, Alberto Pascual-García, Antonio Ferrera, Bartolo Luque, Jordi Bascompte. The architecture of mutualistic networks minimizes competition and increases biodiversity // Nature. 2009. V. 458. P. 1018–1020.
2) George Sugihara, Hao Ye. Complex systems: Cooperative network dynamics // Nature. 2009. V. 458. P. 979–980.
-
вывод: тот, кто дружит со всеми, имеет больше шансов, чем тот, кто со всеми воюет. и в чем здесь новизна?
кроме того, в статье сказано:
"При наличии общего врага у двух видов жертв рост численности одного из видов будет вредить другому не только прямо (через конкуренцию за ресурсы), но и косвенно, через рост численности хищника."
однако в этом случае, жертва с более высокой численностью становится более доступна для хищника (просто по закону вероятности), а значит вредит этим не только другому, но и своему виду.
так что тут еще бабушка надвое сказала.-
Не все так просто. Вложенность (nestedness) оказывается действительно очень важным параметром как трофических, так и мутуалистических сетей. А увеличение хищника и влияние его на исход конкуренции между жертвами - это пример с двумя видами зайцев на Ньфаундленде. Когда туда был завезен европейский беляк, то в ответ очень возросла численность рыси, а услиление пресса рыси на аборигенный вид зайца (кажется,это был Lepus artcicus) привело к его уничтожеению... Ранее этот пример рассматривался как случай непосредственного конкурентного вытеснения.
-
численность хищника (Х), допустим, рыси, зависит от численности их жертв (Y), в данном случае, зайцев. допустим также, что на 100 зайцев приходится 1 рысь, т.е. Y/Х=100. это соотношение в природе колеблется, но математики любят средние числа, их и попридержимся. итак, 1/100.
популяция жертв (зайцев) состоит из 2х видов, аборигенов (А) и иммигрантов (В): a+b=Y ;
путем простых арифметических вычислений выходит, что при любой численности зайцев (a, b или Y), их соотношение с численностью хищника (Х) остается постоянной, 1/100, а значит пресс хищника не меняется независимо от удельной численности одной из популяции зайцев.
эта моделька, как и все добропорядочные матмодели, не учитывает все параметры, поэтому нам следует задаться вопросом: с какого перепугу численность зайцев начала расти (за счет пришельцев)?
и тут на поверхность вылезает толпа факторов, из которых не все имеют численное выражение, как то: лучшая приспособленность к кормовой базе, лучшая сопротивляемость условиям среды или болезням и т.д. что говорит о том, что пресс хищников (сам по себе) вряд ли может являться объяснением.
а в свете последних событий в мире, я нисколько не удивлюсь, если окажется, что зайцы-аборигены вымерли от инфекционных болезней, скажем, заячьего гриппа, занесенного иммигрантами. в этом случае и конкуренция, и пресс хищников вообще могут и оказываются не у дел - одна популяция может вымереть даже при полном отсутствии хищников и изобилии еды просто за счет инфекции.-
Не буду сейчас приводить всю аргументацию, но м.б. для "хроника" будет небезынтересен тот факт, что на природных популяциях гуппи показано, как хищник отдает предпочение массовой форме окраски жертв и избегает более редкую, и речь идет вовсе не о пропорциональном сокращении выбора: Robert Olendorf, F. et al. 2006. Frequency-dependent survival in natural guppy populations // Nature. 2006. V. 441. P. 633-636
Речь идет об отрицательном частотно-зависимом отборе.-
факт был небезынтересен, спасибо. так же интересно было бы узнать какое отношение гуппи имеют к полярным зайцам.
кстати, если вы не заметили, то исходные данные про зайцев и рысь содержали совершенно обратное утверждение. но это нюансы.
осталось только ввести "предпочтения" в виде цифирек в модельки вместо полуабстрактного nestedness.
-
-
-
-


Последние новости









