Что может биоинформатика
М. С. Гельфанд,
доктор биологических наук, кандидат физико-математических наук,
Институт проблем передачи информации РАН
«Химия и жизнь» №9, 2009

Все знают, что биоинформатика — это как-то связано с компьютерами, ДНК и белками и что это передний край науки. Более подробными сведениями может похвастаться далеко не каждый даже среди биологов. О некоторых задачах, которые решает современная биоинформатика, рассказал «Химии и жизни» Михаил Сергеевич Гельфанд (интервью записала Елена Клещенко).
Информация в биологии
В последние десятилетия появилось много новых научных дисциплин с модными названиями: биоинформатика, геномика, протеомика, системная биология и другие. Но по сути, биоинформатика, также как и, скажем, протеомика, — не наука, а несколько удобных технологий и набор конкретных задач, которые решают с их помощью. Можно говорить, что каждый человек, который определяет концентрации белков методом масс-спектрометрии или изучает белок-белковые взаимодействия, работает в области протеомики. Но не исключено, что со временем это деление станет не таким важным: применяемая технология будет менее существенной, чем способ думать, ставить вопросы. И в этом смысле биоинформатика как самая древняя из этих наук — ей целых 25 лет — играет роль цементирующего начала, потому что независимо от того, каким способом получены данные, все равно они потом попадают в компьютер. Иначе быть не может: размер бактериального генома — миллионы нуклеотидов, высшего животного — сотни миллионов или миллиарды. Транскриптомика, изучающая активность генов, получает данные о концентрациях десятков тысяч матричных РНК, протеомика — о сотнях тысяч пептидов и белок-белковых взаимодействиях. С таким количеством информации нельзя работать вручную. Мы еще помним, как печатали на бумаге нуклеотидные последовательности, потом вырезали напечатанные строчки, подставляли друг под друга и таким кустарным способом делали выравнивание — искали сходные участки. Это было возможно, когда речь шла о десятках-сотнях нуклеотидов или аминокислот, но при современном объеме данных нужны специальные инструменты. Набор таких инструментов и предоставляет биоинформатика — в практическом плане это прикладная наука, обслуживающая интересы биологов.
Поскольку моя собственная работа связана в основном с анализом геномных данных, далее речь пойдет главным образом о геномике. Объемы данных еще до появления последнего поколения секвенаторов начали обгонять закон Мура: нуклеотидные последовательности геномов накапливались быстрее, чем росла мощность компьютеров. Не будет большим преувеличением сказать, что за последние годы биология начала превращаться в науку, «богатую данными». Условно говоря, в «классической» молекулярной биологии в одном эксперименте устанавливался один биологический факт: аминокислотная последовательность белка, его функция, то, как регулируется соответствующий ген. А теперь такого рода факты получаются индустриально. Молекулярная биология движется по пути, по которому уже прошли астрофизика и физика высоких энергий. Когда имеется постоянно работающий радиотелескоп или ускоритель, проблема добычи данных решена, и на первый план выступают проблемы их хранения и обработки.
С биологией происходит то же самое, причем очень быстро, и не всегда бывает легко перестроиться. Однако те, кому это удается, оказываются в выигрыше. На нашем семинаре один биолог рассказывал, как они с коллегами изучали некий белок традиционными методами экспериментальной биологии. Это сложная задача: зная, что в клетке выполняется определенная функция, найти белок, который за нее отвечает. Они нашли этот белок, занялись его изучением и убедились, что должен существовать другой белок с подобными свойствами, поскольку наличие первого объясняет не все наблюдаемые факты. Искать второй белок на фоне первого было еще более сложно, но они справились и с этим. А затем был опубликован геном человека — и, получив доступ к его последовательности, они нашли еще дюжину таких белков...
Из этого примера вовсе не следует, что практическая молекулярная биология себя исчерпала. Скорее она научилась пользоваться новыми инструментами: интерпретировать не только полоски в геле после электрофореза, концентрации мРНК и белков или, скажем, скорость роста бактерий, но и колоссальные массивы данных, хранящиеся в компьютере. Заметим, что элемент интерпретации неизбежно присутствует и в классической биологии. Когда исследователь утверждает, что белок А запускает транскрипцию гена В, он не наблюдает напрямую, как белок взаимодействует с регуляторной областью гена, а делает такой вывод из расположения полосок на геле и других экспериментальных данных. В биоинформатике, по сути, та же ситуация, только возведенная в абсолют: готовые данные лежат в компьютере, и среди них нужно отыскать пазлы, из которых получится собрать картинку.
К области технической биоинформатики относится первичная обработка данных. Секвенатор не сам «читает» молекулы ДНК, а дает на выходе кривые флуоресценции, пики на которых еще нужно превратить в нуклеотидную последовательность. Эта задача решается каждый раз по-новому для нового устройства секвенирования, и решает ее биоинформатика. Кроме того, как уже говорилось, полученные данные надо где-то хранить, обеспечивать к ним удобный доступ и т. д. Все это чисто технические проблемы, но они очень важны.
Более сложное и интересное занятие биоинформатиков — получать на основе данных о геноме конкретные утверждения: белок А обладает такой-то функцией, ген В включается в таких-то условиях, гены С, D и Е экспрессируются в одно и то же время, а продукты их образуют комплекс. Именно этим занимаемся мы, и в этом состоит практическое применение нашей науки. Для нас пользователи — другие биологи, которым мы сообщаем интересные для них факты.
Расположение и регуляция
Как можно из последовательности нуклеотидов делать выводы о функции белков и генов? Первое соображение кажется банальным: если белок похож на какой-нибудь другой, уже изученный, то с большой вероятностью он делает примерно то же самое. На самом деле оно не так уж банально: первым серьезным успехом в этом направлении биоинформатики было утверждение, что вирусные онкогены — это «испорченные» гены самого организма.
Выполнить подобное сравнение сейчас уже несложно. Существуют банки данных по нуклеотидным и белковым последовательностям (подробнее о них рассказывалось в «Химии и жизни», 2001, №2). Общее представление о том, как это должно быть устроено, появилось в конце 80-х годов, и в этом смысле биоинформатика была готова к потоку геномных данных. Сегодня это стандартный интернет-сервис: вы загружаете свою последовательность в окошко, нажимаете кнопку, и через несколько секунд вам сообщают, на какие последовательности из этой базы она похожа.
Дальше начинаются более тонкие соображения. Известно, например, что у бактерий гены часто бывают организованы в опероны, то есть транскрибируются в виде одной матричной РНК. Есть разные эволюционные теории, которые объясняют, почему так получилось, что функционально связанные гены образуют оперон. Первая теория состоит в том, что это удобно и полезно, потому и поддерживается эволюцией. Если белки имеют общую функцию, например, отвечают за разные этапы переработки одного вещества, логично, чтобы они появлялись в клетке одновременно, по одному и тому же сигналу (естественно, что при общей мРНК и регуляция одна на всех) и в равном количестве. Второе утверждение менее тривиально и более красиво. Генам, продукты которых имеют связанные функции, выгодно находиться рядом из-за горизонтального переноса. Это очень существенный механизм эволюции бактерий: участки генома одной бактерии попадают в другую, которая благодаря этому может приобрести новые полезные признаки. Понятно, что, если в новый геном переместится лишь один ген метаболического пути, то соответствующий белок будет бесполезен: субстрата для катализируемой им реакции нет, а ее продукт, в свою очередь, некому перерабатывать. Дополнительным подтверждением этой теории служит то, что у бактерий бывают геномные локусы, в которых гены из одного метаболического пути лежат на разных цепях ДНК и потому транскрибируются в разных направлениях. Здесь точно играет главную роль повышенная вероятность совместного переноса.
Тот факт, что два гена находятся рядом в каком-то одном геноме, не очень много говорит про их функциональную связь, это может быть и случайность. Однако мы умеем отождествлять гены в разных организмах. Последовательности у них, конечно, не совпадают до нуклеотида, а могут различаться довольно значительно. Но есть некие правила, которые позволяют утверждать, что это один и тот же ген, скажем, у кишечной и у сенной палочки. Итак, если пара генов находится рядом не в одном геноме, а в пятидесяти, причем у представителей разных таксономических групп (то есть это расположение не просто унаследовано от общего предка), — это означает, что они действительно тяготеют друг к другу. Если бы эволюция не поддерживала их близкого расположения, оно не сохранилось бы. И значит, можно предположить, что они функционально связаны.
Второе соображение похоже на первое. Не все бактерии имеют одинаковый набор генов: к примеру, если ген кодирует фермент, нужный для переработки какого-то углевода, то его не будет у бактерии, которая этим углеводом не питается. Зато у бактерии, которая питается именно этим углеводом, будет весь необходимый набор: и ферменты, и белок-транспортер, переносящий углевод внутрь клетки. Функционально связанные гены присутствуют в геноме по принципу «все или ничего»: как уже говорилось, бессмысленно иметь лишь фрагмент метаболического пути, а бактерии — существа экономные, то, что не приносит пользы, из их генома быстро исчезает. Поэтому если сделать таблицу, где по строкам расположить различные гены, а по столбцам — разные геномы, и отметить плюсами и минусами гены, присутствующие или отсутствующие в данном геноме, мы увидим группы генов, обслуживающих одну и ту же функцию. И неизвестный ген с тем же набором плюсов и минусов, что у некой группы, скорее всего, можно приписать к ней же.
Третье соображение связано с регуляцией активности генов. Рядом с геном обычно присутствуют участки, с которыми взаимодействуют определенные белки — они могут запускать транскрипцию, блокировать ее, управлять ее интенсивностью, иначе говоря, от них зависит активность гена в каждый момент времени. Некоторые регуляторные участки очень хорошо опознаются по характерным последовательностям «букв», но это бывает редко. Например, участки связывания факторов транскрипции мы распознаем в геномах с невысокой точностью и вместе с правильными сайтами нагребаем кучу «мусора» — похожие коротенькие участки, которые на самом деле не имеют отношения к регуляции генов. Но поскольку совместно регулируются те гены, которые совместно работают, настоящие сайты связывания находятся перед одними и теми же генами в десятке геномов, а случайные — раскиданы там и сям, и никакой закономерности в их расположении не прослеживается. Получается мощный фильтр, позволяющий отсеять «мусор». И если перед геном с неизвестной функцией устойчиво обнаруживается знакомый сайт, будет ясно, что этот ген регулируется в составе функциональной подсистемы, которая регулируется тем же регулятором и обеспечивает ту же функцию.
Мне интереснее всего изучать эволюцию регуляторных систем, но побочным продуктом при этом бывает множество функциональных предсказаний. Исследование развивается как детектив: каждое соображение по отдельности очень мелкое, но если «улик» много и они все попадают в одну точку, то можно делать уверенные утверждения. Был случай, когда мы подробно описали регуляторную систему — фактор транскрипции, сайты его связывания, то, что это будет репрессор, а не активатор, то, что связывание будет требовать кооперативного взаимодействия двух димеров, — просто глядя на буковки генома. Впоследствии все это вплоть до деталей оказалось правильным.
Рибосома как депо цинка
В одной из таких работ центральную роль сыграла Екатерина Панина, на тот момент студентка мехмата МГУ (потом она поступила в аспирантуру Калифорнийского университета Лос-Анджелеса и стала настоящим биологом-экспериментатором). Она пришла к нам на третьем курсе и сказала, что хочет заниматься такой биологией. К окончанию мехмата у нее было опубликовано несколько статьей в серьезных журналах.
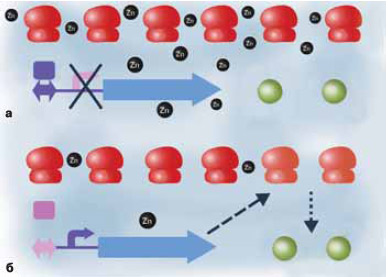
Бактериальной клетке нужны ионы цинка: они, например, входят в состав некоторых ферментов как кофакторы. Соответственно есть и молекулярная машинерия, которая обслуживает все процессы, связанные с цинком. Мы изучали цинковый репрессор (в больших количествах цинк ядовит для клетки, поэтому выключать его транспорт при достаточных концентрациях не менее важно, чем уметь добывать его из окружающей среды), используя идеологию, о которой рассказывалось в предыдущей главке. Если перед геном имеется потенциальный сайт цинкового репрессора, то этот ген, возможно, относится к метаболизму цинка. Именно таким образом мы в свое время «вычислили» цинковый транспортер — трансмембранный белок, который обеспечивает проникновение цинка внутрь клетки.
Так вот, в 2002 году Катя обратила внимание, что потенциальные сайты цинкового репрессора почему-то часто попадаются перед генами рибосомных белков. Она поделилась этим наблюдением с научным руководителем, и я сказал, что, поскольку в геноме больше сотни генов рибосомных белков, а сайты встречались перед разными генами, это случайность. Но Катя в случайность не поверила и нашла статью Евгения Кунина (о его модели происхождения клетки см. в статье М. А. Шкроб в августовском номере), которая была опубликована незадолго до этого. Там было показано, что некоторые рибосомные белки содержат мотив связывания цинка — так называемую цинковую ленту, три или четыре цистеина на правильном расстоянии друг относительно друга и в правильном контексте. Важное наблюдение Кунина с коллегами состояло в том, что один и тот же белок в некоторых организмах имеет эти цинковые мотивы, в других — не имеет, но, судя по всему, нормально функционирует и без цинка. А у некоторых бактерий один и тот же белок имеется в двух вариантах, с цинковой лентой и без нее.
И вот Катя заметила, что в последнем случае, когда есть два варианта белка в одном геноме, тот, который без цинковой ленты, репрессируется цинковым репрессором. Иначе говоря, в присутствии цинка экспрессируется вариант белка, которому цинк нужен, а в отсутствие цинка — тот, которому он не нужен.
Основа существования любой клетки — тяжелая промышленность, производство средств производства, точно так, как нас учили на лекциях по политэкономии социализма. Около 70% белка клетки — это белки рибосом, то есть органелл, которые нужны, чтобы делать другие белки. С другой стороны, цинк — кофактор ферментов, жизненно важных для клетки, таких, например, как ДНК-полимераза. Если цинка становится мало, его полностью забирают себе рибосомные белки, ферментам ничего не остается, и клетка погибает. Но у клетки есть резервная копия рибосомного белка, которому цинк не нужен. Мы предположили, что клетка включает синтез таких белков в условиях дефицита цинка и они встраиваются в часть рибосом на место цинксодержащих белков. При этом какое-то количество цинка высвобождается. Может быть, рибосомы после этого работают чуть менее эффективно, может быть, и вообще не работают — но ради того, чтобы цинка хватило жизненно важным ферментам, которые представлены существенно меньшим числом копий, стоит пожертвовать небольшой долей рибосом.
Мы написали статью, но в течение года ни один уважаемый журнал не принял к публикации безумную теорию о рибосомах как депо цинка. Однако мне Катина находка казалась очень красивой, и я единственный раз в жизни воспользовался тем, что мой дед, как член Академии наук США, имеет право представлять статьи для публикации в «Proceedings of the National Academy of Sciences of the USA». Он послал статью на рецензию Кунину, который дал положительный отзыв (и, кажется, кому-то еще). Статья вышла в PNAS, и, как вскоре выяснилось, очень вовремя: через полгода появилась статья японских биологов, которые экспериментально показали то же самое. Можно догадаться, что они над этим работали давно, и, вероятно, им было немного обидно, что компьютерное предсказание предвосхитило их результаты.
Заметим, что вся эта история построена на очень мелких частных наблюдениях (есть в белке цистеины — нет цистеинов, есть потенциальный сайт репрессора — нет сайта...). Но в совокупности эти мелочи позволили сделать нетривиальное заключение, оказавшееся абсолютно верным. Вообще, когда мы публикуем статьи, то стараемся как можно более четко сказать, какое из наших предсказаний считаем надежным, а какое может оказаться неправильным. Так вот, среди тех, в которых мы были уверены, неправильных пока не оказалось ни одного (проверены уже десятки), а вот среди слабых проколы действительно были, хотя тоже не часто.
Отвертка со съемным жалом
Не менее красивыми были работы с белками-транспортерами (я в них участвовал только на ранних стадиях, поэтому имею полное право их хвалить, не становясь хвастуном). Транспортеры — золотое дно для биоинформатиков, поскольку опознать транспортер, в особенности бактериальный, достаточно легко. У них есть несколько гидрофобных спиралей, проходящих через мембрану: между ними находится канал, сквозь который ион или молекула, нужные для жизни клетки, проникают внутрь. Трансмембранные сегменты можно найти в белковой последовательности с помощью специальных программ. И если в неизвестном бактериальном белке пять или шесть таких сегментов, это почти наверняка транспортер (потому что другие трансмембранные белки, например участники дыхательной цепи или родопсин, хорошо известны). Остается установить, что за вещество он переносит.
Изучать специфичность транспортеров в эксперименте — удовольствие ниже среднего. С ферментами гораздо проще, это практически рутинная задача, которую можно доверить роботу. Вы гиперэкспрессируете фермент (то есть заставляете синтезироваться в больших количествах), а потом предлагаете ему пять сотен разных субстратов и смотрите, с каким из них пойдет реакция.
Транспортер, конечно, тоже можно гиперэкспрессировать. Но чтобы он заработал, он должен сразу встроиться в мембрану, иначе гидрофобные сегменты «налипнут» друг на друга, белок образует нефункциональные агрегаты. Поэтому приходится делать множество мембранных пузырьков-везикул, встраивать в них белки в правильной ориентации, а потом смотреть, попадает ли искомое вещество внутрь везикул. Вдобавок транспортеры бывают разные. Одни закачивают в клетку полезные вещества против градиента концентрации и затрачивают на это энергию молекулы АТФ, которую расщепляет специальный белок — АТФаза. Другие осуществляют вторичный транспорт — впуская «нужную» молекулу, одновременно выпускают по градиенту концентрации ион водорода, калия или натрия. Если транспортер АТФ-зависимый, то, чтобы он заработал, необходимо собирать конструкцию из нескольких белков, в том числе АТФазы. А если это вторичный транспорт, то нужно еще угадать, концентрацию какого иона надо увеличить внутри шарика. Отсюда ясно, что биохимия транспортеров — наука для сильных духом и экспериментальных данных по ним мало.
С другой стороны, определять специфичность транспортеров биоинформационными методами существенно проще. Достаточно прибегнуть к уже знакомой логике: например, если синтез этого белка регулируется цинковым репрессором, это, скорее всего, и будет цинковый транспортер, а если его ген находится в одном локусе с генами катаболизма рибозы, он, очевидно, переносит внутрь клетки рибозу... Именно таким образом мы в свое время нашли рибофлавиновый транспортер: имеется белок с неизвестной функцией, у него шесть потенциальных трансмембранных сегментов, регулируется совместно с генами рибофлавинового пути — значит, это транспортер либо рибофлавина, либо его предшественника. Но поскольку в некоторых геномах присутствовали и этот транспортер, и рибофлавин-зависимые белки, но не было пути синтеза рибофлавина из предшественников, значит, это мог быть транспортер только рибофлавина.
Проверять экспериментально конкретное предсказание существенно проще, чем начинать с нуля. Я всегда объясняю студентам, что биоинформатик — существо совершенно беззащитное, наподобие того персонажа приключенческого романа, который знает, где лежит клад. Пока он молчит, все его берегут и за ним ухаживают, но, когда он проговорится, он уже не нужен. Как только биоинформатик сказал «этот белок обладает такой-то функцией» — исключительно от порядочности экспериментаторов зависит, возьмут ли они его в соавторы после того, как проверят это утверждение. А утверждения, как читатель уже убедился, предельно простые и конкретные, достаточно один раз произнести их вслух.
С таких же простых умозаключений начиналась история более сложная, но и более интересная. Мы изучали регуляцию пути биосинтеза биотина (биотин — витамин Н, или В7, кофактор многих важных ферментов). Биотиновый транспортер был в это время не известен. У нас по ходу работы обнаружился транспортный белок, который регулируется, а иногда и локализуется вместе с генами биотинового пути. Дальше все как с рибофлавином: нашлись организмы, где биотинового пути нет, но есть белки, которые зависят него как от кофактора, и есть тот самый потенциальный транспортер — следовательно, это транспортер биотина.
Как уже было сказано, транспортеры бывают АТФ-зависимые и осуществляющие вторичный транспорт. Биотиновый транспортер был одиноким, никакого гена АТФазы поблизости не просматривалось, а значит, это был вторичный транспортер. Но затем мы увидели, что в некоторых геномах рядом с биотиновым транспортером попадаются какие-то АТФазы. Что это означает, на том этапе было непонятно, и потому мы просто упомянули про это в статье одной фразой.
Примерно тогда же мы изучали регуляцию кобаламинового пути. Кобаламин, или витамин В12, —также кофактор важных ферментов, очень крупная молекула с метаболическим путем соответственной сложности. Для этой истории существенно, что в центре молекулы кобаламина есть ион кобальта, который приносят в клетку опять же транспортеры. Таких транспортеров мы нашли немало, опубликовали о них статью — и в скором времени получили письмо от Томаса Эйтингера из Института микробиологии Гумбольдтовского университета (Берлин). Он призывал нас обратить внимание на то, что любой кобальтовый транспортер также может транспортировать никель, и наоборот, потому что специфичность у них слабая. Мы ответили, что рассматриваем транспортеры с точки зрения их функциональной роли в клетке, и если ген белка находится в одном опероне с большим набором генов кобаламинового синтеза — безусловно, белок нужен клетке как транспортер кобальта, хотя in vitro его и можно заставить переносить никель. А если мы видим ген транспортера в одном опероне с никель-зависимой уреазой, то это, безусловно, никелевый транспортер.
Намечались перспективы совместной работы, и Дмитрий Родионов, который делал эту работу, подал вместе с немецкими коллегами заявку на небольшой совместный грант и поехал на три месяца в Берлин. (Дмитрий закончил МИФИ, после чего занимался у нас геномикой; потом работал в США, а сейчас выиграл грант академической программы «Молекулярная и клеточная биология» на создание новой группы и возвращается в Москву.)
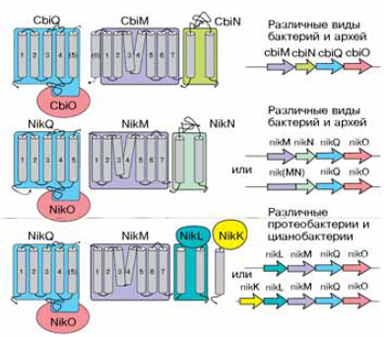
К этому времени мы с ними начали делать (по электронной почте) большой проект по сравнительной геномике транспортеров никеля и кобальта, где классифицировали их, во-первых, по регуляции, а во-вторых, по локализации, совместной с кобальтовыми или никелевыми функциональными белками. Так вот, в одном из этих никелево-кобальтовых семейств наблюдались некоторые странности. С одной стороны, АТФазы и трансмембранные белки, образующие канал для иона, как положено, располагались рядом и регулировались совместно. С другой стороны, в том же опероне мог находиться еще один трансмембранный белок. Причем эти «посторонние» белки в кобальтовых и никелевых транспортных системах отличались довольно сильно, не были гомологичными в отличие от АТФаз и трансмембранников. И вдобавок АТФаза и трансмембранный белок оказались гомологичными тем самым «лишним» биотиновым белкам, которые то попадались, то нет в предыдущем исследовании.
До сих пор не знаю, каким способом Дима уговорил немецких коллег на следующий безумный эксперимент. «Классическим» биохимикам, которые всю жизнь изучают транспорт кобальта и никеля у бактерий, он предложил: давайте у транспортера отключим АТФазу и трансмембранный белок, гомологичные биотиновым, оставим один только уникальный компонент. Ведь биотиновому транспортеру АТФаза и «основной» трансмембранник не очень нужны, они то есть, то их нет, — может быть, они и никелевому транспортеру не нужны, одинокий негомологичный трансмембранник и сам справится? Неизвестно, почему добропорядочные немецкие биохимики решились на это странное деяние: лишить вроде бы обычный АТФ-зависимый транспортер АТФазы и посмотреть, что будет. Так или иначе, Дима оказался прав. Одинокий трансмембранник работал как кобальтовый транспортер — менее эффективно, но работал. Это был первый пример двойной системы, которая, если есть АТФаза, работает как АТФ-зависимая, а если ее нет, работает как ион-зависимая.
Позднее берлинские коллеги то же самое сделали с биотином: взяли бактерию, у которой биотиновый транспортер имеет АТФазу и трансмембранник, отключили их гены — и показали, что этот белок в одиночестве тоже работает как биотиновый транспортер, хотя и с меньшей мощностью, чем в присутствии АТФазы.
Дмитрий Родионов в это время уже работал постдоком в лаборатории Андрея Остермана в Институте медицинских исследований Бэрнема в Ла-Хойе. Остерман — замечательный человек, биохимик, который понял эффективность биоинформатических методов, научился ими пользоваться и нашел с их помощью множество новых ферментов. И вот, когда Дмитрий попал в круг биохимиков и начал с ними общаться, оказалось, что подобных транспортеров, переносящих разные субстраты — кофакторы, аминокислоты, ионы, — существует несколько десятков. (Кстати, таким же оказался и рибофлавиновый транспортер.) Разные исследовательские группы независимо друг от друга изучали эти транспортеры, не имея представления о том, что они принадлежат к одному семейству.
Стало понятным и то, как возможна подобная организация. Кобальтовые и никелевые транспортеры отдельно от своей АТФазы не встречаются (если ее не убрать экспериментально). Но есть и другой класс бактериальных транспортеров, которые используют одну и ту же АТФазу — как отвертку со съемным жалом. Универсальные АТФаза и трансмембранный белок в этом случае могут кодироваться вместе с рибосомными белками, то есть экспрессируются постоянно и в больших количествах. А те белки, которые обеспечивают транспортерам специфичность, раскиданы там и сям в соответствующих оперонах. А в отсутствие АТФазы такой белок худо-бедно работает как вторичный транспортер, и поэтому в геномах некоторых организмов мы видим только его.
Биоинформатика и теория эволюции
Однако эти «прикладные» открытия — дело очень важное и полезное, но для нас, биоинформатиков, не главное. А главное, что принесла нам индустриальная революция в биологии, — появилась возможность на другом уровне обсуждать эволюцию. Даже банальные утверждения, скажем, о процентном сходстве геномов человека и шимпанзе нетактривиальны, как могут показаться. Молекулярная эволюция поучительна тем, что на ней замечательно выполняются дарвинистские представления о природе вещей.
Данные, полученные молекулярными биологами, теперь оказывают серьезное влияние на таксономию — классификацию растений и животных. Поначалу ботаники и зоологи скептически относились к молекулярным генеалогическим деревьям, показывающим степень родства между видами на основе сравнения нуклеотидных последовательностей, но надо признать, что и первые молекулярные деревья были не слишком удачными. Сейчас прямо на глазах происходит конвергенция — классическая и молекулярная таксономии движутся навстречу друг другу. Уже понятно, что молекулярные деревья, если они построены с соблюдением определенных правил, достаточно близки к реальности и вполне могут стать поводом для пересмотра ортодоксальных таксономических представлений, основанных на морфологии — сравнении внешних черт организмов. И, как ни странно, оказывается, что у видов, которых будто бы насильно помещают вместе исходя из сходства их генов, действительно отыскиваются общие признаки. Получается, что хорошее молекулярное дерево не противоречит морфологической конструкции, просто другие признаки оказываются ведущими.
Что касается бактерий, то в эпоху классической биологии их классифицировали по форме клеток и по метаболическим свойствам: какие сахара они могут утилизировать, какие аминокислоты и кофакторы могут синтезировать сами, а в каких нуждаются как в составной части внешней среды и т. п. Эта таксономия была очень слабой, поскольку у бактерий по сравнению с высшими организмами очень мало морфологических и функциональных признаков. Сегодня таксономия бактерий, по-видимому, полностью основывается на молекулярных данных. В массовом порядке пересматриваются видовые названия. Но самым впечатляющим достижением в этой области была, конечно, работа Карла Вёзе, который в 1977 году на основании молекулярной таксономии постулировал существование архебактерий (сейчас их называют археями) — третьего домена жизни, отличного от эукариот и «настоящих» бактерий.
Нельзя сказать, что все проблемы систематики бактерий отныне решены. В значительной мере оказалось разрушенным представление о том, что такое бактериальный вид. Обнаружилось, например, что у двух штаммов кишечной палочки — представителей одного вида — до трети генов могут быть уникальными, то есть присутствовать в одном штамме и отсутствовать в другом. Много неожиданного и интересного уже известно о бактериальной эволюции. В частности, оказалось, что горизонтальный перенос — обмен генетическим материалом — может происходить между таксономически далекими существами. Например, Metanosarcina — типичная архея, но треть ее генов имеют бактериальное происхождение, и эти гены обслуживают практически весь ее метаболизм, в то время как механизмы транскрипции, трансляции, репликация, устройство мембраны у метаносарцины характерны для архей. По этому примеру можно судить о том, насколько увлекательно сейчас заниматься эволюцией бактерий.
На мой взгляд, самое интересное — это эволюция регуляторных систем. Мы достаточно много знаем про эти системы у бактерий и можем представить, как меняются регуляторные системы, как локальный регулятор вдруг начинает управлять десятками генов или меняет специфичность, как перестраиваются регуляторные каскады. И это может быть очень важно с фундаментальной точки зрения, потому что здесь можно пойти гораздо дальше. Отличие человека от шимпанзе или даже от мыши едва ли обусловлено набором генов: они у млекопитающих практически одни и те же, если сравнивать по набору функций. Причина скорее в регуляции: какие гены, когда и в каких тканях активны.
Скорее всего, «скачки» эволюции, любые резкие изменения морфологических признаков обеспечиваются как раз на уровне регуляции. Мы уже знаем такие примеры у бактерий, дрожжей и других относительно простых организмов. У большинства бактерий имеется один железный репрессор, который реагирует на присутствие ионов железа и регулирует множество генов: белки, обеспечивающие запасание и транспорт железа, железозависимые ферменты. А у других бактерий есть три разных репрессора, которые эти функциональные группы поделили: одни регулируют запасание железа, другие транспорт и синтез, третьи — ферменты. Это на самом деле радикальное изменение, был один ответ на железо, а получилось три разных.
Есть замечательные экспериментальные работы, выполненные на многоклеточных. Почему морской еж единственный среди иглокожих имеет твердый скелет? Ответ предложил Эрик Дэвидсон из Калифорнийского технологического института. Он изучил регуляторный каскад, который отвечает за развитие этого скелета, а потом нашел этот каскад у морской звезды, только у нее он включается существенно позднее, поэтому развиваются лишь основания иголочек, не соединенные между собой. У ежа тот же каскад включается на какое-то количество клеточных делений раньше, соответственно захватывает большее число клеток, и развивается сплошной скелет. Таким образом, чисто регуляторное изменение дает абсолютно новый признак.
У меня есть надежда, что сравнительный анализ регуляции даст ответы на вопрос, который беспокоит палеонтологов и морфологов на нынешнем этапе развития синтетической теории эволюции: каким образом накопление мелких изменений дает радикально новые признаки? Похоже, что это можно объяснить перенастройкой регуляции. Мы уже умеем это делать на простых организмах, но рано или поздно очередь дойдет и до более сложных. И когда это случится, произойдет третий большой прорыв в этом направлении, если первым считать дарвиновский естественный отбор, а вторым — соединение эволюционной биологии с генетикой.
-
Михаил Гельфанд? Это же тот придурок, который не читав принял статьи в научный журнал, сгенерированные компьютером цепями маркова по шаблону. Он еще крупно опозорился тогда, и его погнали из института
-
Вы всё немножко перепутали, см. https://ru.wikipedia.org/wiki/Корчеватель_(статья).
-







