Фолдинг белка
Белки — это биологические молекулы, выполняющие тысячи специфических функций внутри каждой клетки живого организма. Белки синтезируются в рибосомах в виде длинной полипептидной нити, но затем быстро сворачиваются в свою естественную («нативную») пространственную структуру. Этот процесс называется фолдинг белка. Может показаться удивительным, но этот фундаментальный процесс до сих пор плохо понят на молекулярном уровне. В результате предсказать нативную структуру белка по его аминокислотной последовательности пока не удается. Для того чтобы почувствовать хотя бы некоторые нетривиальные аспекты этой задачи, попытаемся решить ее для следующей исключительно простой модели белковой молекулы.
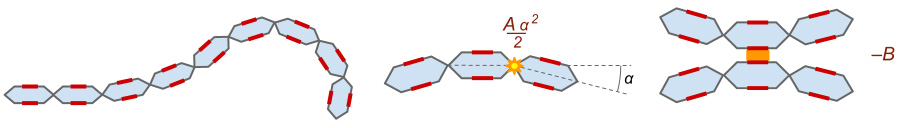
Пусть белок состоит из совершенно одинаковых звеньев, последовательно соединенных друг с другом (рис. 1). Эта цепочка может изгибаться, и для простоты будем считать, что она изгибается не в пространстве, а только в плоскости. Цепочка имеет определенную упругость на изгиб: если направления двух соседних звеньев образуют угол α (измеряемый в радианах), то такое соединение повышает энергию молекулы на Aα2/2, где A — некоторая константа размерности энергии. Пусть также у каждого звена по бокам имеется два «контактных участка», которыми звенья могут склеиваться. Каждая такая склейка обладает энергией –B (то есть она понижает энергию цепочки на величину B). Наконец, будем предполагать, что B меньше A (то есть цепочка достаточно упруга).
Задача
Какая конфигурация молекулы из N звеньев будет наиболее энергетически выгодной? Исследуйте, как меняется эта конфигурация с ростом N.
Подсказка
Наиболее энергетически выгодной является конфигурация с минимальной энергией. Поэтому надо придумать, как устроить большое число «склеек» звеньев (каждая их которых понижает энергию), но при этом не слишком резко изгибать цепочку, чтобы чересчур сильно не увеличивать ее упругую энергию.
В этой задаче не требуется искать абсолютно точную форму цепочки для каждого конкретного числа звеньев. Надо лишь описать характерные «узоры», которые будут возникать при оптимальном фолдинге этой «белковой молекулы», и найти, при каком примерном N молекуле выгодней перестроиться из одной конфигурации в другую.
Решение

Энергия абсолютно прямой цепочки равна нулю. Для того чтобы понизить ее, некоторые звенья должны слипнуться. Но для этого цепочка должна организовать петлю, и наличие петли повышает энергию. Если петля слишком длинная, то большое количество звеньев, которые могли бы связаться друг с другом, остаются без связи. Эти звенья можно соединить, словно на застежке-молнии, укоротив тем самым петлю, но от этого увеличится ее энергия упругости. Поэтому надо найти такую оптимальную длину петли, при которой силы упругости, расширяющие петлю, и силы связи, ее «застегивающие», сбалансированы.
Энергия петли
Пусть имеется петля из m несклеенных звеньев (рис. 2). Характерный угол между соседними звеньями в ней — примерно 2π/m. (На самом деле, этот угол меняется от звена к звену, поскольку наиболее выгодная форма петли вовсе не круговая, но для приближенного исследования наша оценка вполне пойдет.) Таких соединений имеется m штук, поэтому петля обладает энергией 2π2A/m. Застегнем ее еще на одно звено. Тогда петля станет короче на два звена, а энергия всей цепочки изменится на величину
Если же, наоборот, разорвать одну связь, то энергия цепочки изменится на
Петля из m звеньев является оптимальной, когда оба эти изменения энергии положительны, то есть с энергетической точки зрения петлю невыгодно ни удлинять, ни укорачивать. Поскольку B много меньше A, ясно, что величина m получится значительно больше единицы. Поэтому для примерной оценки оптимального m эти два неравенства можно заменить одним равенством:
![]()
Таким образом, оптимальная длина петли примерно равна
![]()
Во всех последующих формулах под буквой m будет подразумеваться именно оптимальная длина петли. Наконец, полезно найти энергию упругости такой оптимизированной петли; она получается равной
![]()
Это выражение (энергия петли в m/2 раз больше величины B) очень удобно для дальнейших вычислений.
Когда появляется петля?
Теперь легко выяснить, при цепочке какой длины будет выгоднее не оставаться прямой, а свернуться в петлю с «двойным хвостиком» длины n. Для этого нужно, чтобы полная энергия такой конфигурации была отрицательна:
![]()
Таким образом, если длина цепочки N > m + 2(m/2) = 2m, то ей выгоднее образовать петлю.
Когда появляется вторая петля?

«Двойной хвостик» — это не максимально удобная конфигурация, поскольку в каждом звене «работает» только один из контактных участков, а хотелось бы, чтоб работали оба, хотя бы у некоторых звеньев. Это можно устроить, образовав вторую петлю (рис. 3).
Для того, чтобы сосчитать, когда переход к двум петлям энергетически выгоден, возьмем цепочку длины N и сосчитаем ее энергию для одной петли и для двух петель:
Условие для перехода к двумя петлям, E1 > E2, тогда даст N > 8m.
Очень длинная цепочка
Когда цепочка становится очень длинной, ее удобно сворачивать так, чтобы как можно большее количество звеньев было склеено обоими своими контактными участками. Таким образом мы получаем конфигурацию, напоминающую обрамленное петельками полотно. Если закрыть глаза на то, что соседние петли мешают друг другу, можно провести аналогичное вычисление и найти наиболее выгодное количество петель для заданного N (оно растет пропорционально квадратному корню из N). Если же учесть, что петли мешают друг другу, то вычисления резко усложнятся. Однако общая структура останется той же: наиболее выгодным будет плоское полотно некоторой формы, обрамленное по краям петельками. Желающие могут попробовать найти оптимальную форму полотна с помощью компьютерного моделирования, а также поразмышлять над аналогичной задачей в трехмерном пространстве.
Послесловие
Эта простая задача, конечно же, не может отразить ни закономерностей фолдинга настоящих белковых молекул, ни тех методов современной теоретической физики, которые применяются при описании белков и полимеров (эта область деятельности, кстати, является вполне серьезным разделом физики конденсированных сред). Цель этой задачи состояла лишь в демонстрации того, как «количество переходит в качество», то есть как при изменении лишь одного численного (а не качественного) параметра задачи может принципиально меняться ее решение.
Задачу можно было бы сделать чуть более «живой» и интересной, если ввести ненулевую температуру. В этом случае оптимальная конфигурация определялась бы не только энергией, но и энтропией, она бы тогда отвечала минимуму так называемой свободной энергии молекулы. При изменении температуры тогда происходил бы настоящий фазовый переход, при котором молекула сама бы распрямлялась, сворачивалась или перестраивалась из одной формы в другую. К сожалению, такая задача потребует методов, которые выходят за рамки школьной программы.
Любопытно также заметить, что теоретическое изучение фолдинга белков вовсе не сводится к одному лишь численному моделированию. В этой, казалось бы, «прямолинейной» задаче вскрываются довольно нетривиальные математические тонкости. Более того, имеются даже работы, в которых для описания этого процесса привлекаются методы квантовой теории поля и теории калибровочных взаимодействий.
Потренироваться на практике в поиске оптимальной конфигурации белка можно на сайте Fold.it.
-
-
Витки спирали разные, поэтому там регулярно звенья будут не попадать друг на друга. Так что какой-то ненулевой (и довольно) большой процент связей будет не задействован. А в конфигурации типа полотна процент незадействованных связей стремится к нулю с ростом числа звеньев.
В трехмерном пространстве, кстати, спираль была бы удобна.-
Между прочим, неозвученная, но подразумеваемая вами формулировка, в которой в спирали "регулярно звенья будут не попадать друг на друга", нефизична. Она предполагает, что энергия контакта есть только для звеньев с чётко совпадающими гранями. Такое требование настолько сложно вообразить выполняющимся в практической ситуации, что его не стоит принимать даже как нулевое приближение.
-
Конечно задачу можно усложнять, можно ввести например частичный контакт с энергией пропорциональной длине слипания, или можно сделать цепочку слегка растягиваемой. Но в какой-то момент надо остановиться: задача не для студентов, а для широкой публики и для школьников, и больше, чем одну идею в нее вставлять нежелательно.
-
-


> описания этого процесса привлекаются методы
> квантовой теории поля и теории калибровочных взаимодействий.
Какой снобизм. Вообще-то задачами не то что фолдинга белка, а хотя бы агрегации (коллапса, глобулообразования) гомополимерной цепи сразу занимались люди, пришедшие из весьма "навороченных" областей теорфизики: И. М. Лифшиц, С. Эдвардс, де Жен, де Клуазо, Е. Гельфанд и прочие. Это было полвека назад, и сразу применялись формализм квантов и КТП (например, континуальный интеграл, ренорм-группы, метод случайных фаз), топологии (полиномы Александера, Гаусс-Бонне) и куча всего прочего (метод реплик, скажем). Статистическая физика полимеров вообще основывалась на работах научных школ (московская, парижская, кембриджская, страсбургская, массачусетская), которые к тому моменту заслужили признания в "высоких" разделах теорфизики (физика магнетизма, в первую очередь). На память я могу назвать лишь одну заметную школу, пришедшую в статфизику полимеров не из теорфизики: ленинградскую (Волькенштейн), а также одного полимерщика, недостаток классического физического образования которого компенсировался его гениальностью - П. Флори.



