Эволюция белков сдерживается низкой проходимостью ландшафта приспособленности
Рецепторный белок PhoQ передает сигнал из внешней среды внутрь клетки, взаимодействуя с белком PhoP. Ключевую роль в работе PhoQ играют четыре аминокислотные позиции, образующие «белок-белковый интерфейс» и позволяющие безошибочно распознавать PhoP. Американские биологи экспериментально изучили свойства всех возможных аминокислотных комбинаций, занимающих эти ключевые позиции. Неожиданно оказалось, что из 160 000 комбинаций целых 1659 (более 1%) успешно справляются с функцией распознавания PhoP. Это означает, что «белковый код», обеспечивающий узнавание белками друг друга, обладает высокой избыточностью, подобно генетическому коду. При этом эволюционные маршруты от одних «разрешенных» последовательностей к другим, как правило, оказываются длинными и окольными. Это объясняется сильным эпистазом — зависимостью полезности мутации от того, какие мутации успели закрепиться ранее. Возможно, из-за эпистаза эволюция не успевает найти многие потенциально удачные решения, а ее пути становятся до некоторой степени предсказуемыми.
Чтобы понять, как эволюционируют белки, очень важно расшифровать структуру ландшафта приспособленности конкретных белков. В схематичном виде ландшафт приспособленности показан на рис. 1 (подробно о понятиях «пространство последовательностей» и «ландшафт приспособленности» рассказано в новости Расширение белковой вселенной продолжается, «Элементы», 24.05.2010). Конечно, лучше всего было бы провести полное картирование пространства последовательностей, определив для каждой его точки эффективность выполнения белком интересующей нас функции. Однако такая задача невыполнима, поскольку даже для маленькой белковой молекулы длиной в 100 аминокислот число точек в пространстве последовательностей (то есть число всех возможных аминокислотных последовательностей такой длины) на много порядков превышает число атомов во Вселенной (первое равно 20100 ≈ 10130, второе — в 1050 раз меньше).
Поэтому ученым приходится ограничиваться картированием очень маленьких, специально отобранных участков пространства последовательностей. Об одном таком исследовании рассказано в новости Пути эволюции предопределены на молекулярном уровне, «Элементы», 12.04.2006. В этом случае рассматривалось всего пять аминокислотных позиций и по две возможных аминокислоты в каждой из них, то есть всего 25 = 32 точки в пространстве последовательностей.
Сегодня, благодаря стремительному развитию биотехнологий, появилась возможность картировать намного более обширные области. Об этом свидетельствует статья Анны Подгорной (Anna I. Podgornaia) и Майкла Лауба (Michael T. Laub) из Массачусетского технологического института, опубликованная в свежем выпуске журнала Science. Авторы сосредоточили усилия на четырех ключевых аминокислотах, входящих в состав белка PhoQ кишечной палочки Escherichia coli. Белки, гомологичные PhoQ, есть и у других гамма-протеобактерий, таких как сальмонелла и чумная палочка. PhoQ — это рецептор, располагающийся на внутренней мембране бактерии. PhoQ реагирует на изменения концентрации ионов Mg2+ в периплазматическом пространстве и передает сигнал в цитоплазму посредством активации или инактивации белка PhoP (см. PhoP-PhoQ). Белок PhoP, в свою очередь, включает и выключает гены, необходимые для адаптации бактерии к меняющимся условиям среды (рис. 2).

Рис. 2. Регуляторная система PhoQ-PhoP. При низкой концентрации ионов магния (слева) PhoQ фосфорилирует PhoP, переводя его в активное состояние (P — остаток фосфорной кислоты). Активированный PhoP регулирует экспрессию множества генов. При высокой концентрации магния (справа) PhoQ инактивирует PhoP путем дефосфорилирования. У подопытных бактерий в геном был вставлен ген желтого флуоресцирующего белка (yfp), соединенный с промотором PmgrB. Этот промотор обеспечивает экспрессию yfp при наличии в клетке активированного PhoP, что позволяет по силе свечения судить об исправности регуляторной системы PhoQ-PhoP. Четыре звездочки на белке PhoQ символизируют четыре ключевые аминокислоты, необходимые для взаимодействия с PhoP. Рисунок из обсуждаемой статьи в Science
Ключевую роль в работе двухкомпонентной регуляторной системы PhoQ-PhoP играют четыре аминокислоты, входящие в состав PhoQ и образующие так называемый белок-белковый интерфейс. Эти аминокислоты необходимы для того, чтобы PhoQ мог безошибочно опознать своего партнера PhoP и правильно провзаимодействовать с ним.
В белке PhoQ кишечной палочки эти четыре позиции заняты следующими аминокислотами: аланин в позиции 284, валин в позиции 285, серин в позиции 288, треонин в позиции 289 (сокращенно Ala284, Val285, Ser288, Thr289, или еще короче — AVST).
Но AVST — лишь одна из 160 000 (204) возможных комбинаций четырех аминокислот. Авторы поставили перед собой амбициозную задачу: картировать всё пространство последовательностей для данных четырех аминокислотных позиций. То есть для каждой из 160 000 комбинаций проверить, обеспечивает ли она нормальное взаимодействие PhoQ с PhoP, а значит, и правильную работу всей регуляторной системы.
Для этого были изготовлены плазмиды с геном phoQ, в котором триплеты нуклеотидов, кодирующие аминокислоты белок-белкового интерфейса AVST, были полностью рандомизированы, то есть заменены на случайные тройки нуклеотидов. Авторы использовали метод сайт-направленного мутагенеза с помощью ПЦР (см.: Site-directed mutagenesis). Затем эти плазмиды вставили в клетки E. coli, из которых их собственный ген phoQ был заблаговременно удален. Кроме того, в геном подопытного штамма вставили ген желтого флуоресцирующего белка под управлением промотора, контролируемого белком PhoP (рис. 2). В итоге получилась разношерстная популяция кишечных палочек, у которых в гене phoQ на месте четырех триплетов, кодирующих аминокислоты AVST, находились все возможные триплеты, кодирующие все 160 000 возможных аминокислотных комбинаций (комбинаций на самом деле получилось больше, потому что есть ведь еще и стоп-кодоны, но те бактерии, которым так не повезло, не были предметом дальнейшего изучения).
Теперь нужно было выбрать из этой популяции те клетки, у которых система PhoQ-PhoP по-прежнему исправно работала. Отбор проводился в два этапа. Сначала микробов выращивали при разных концентрациях магния и отбирали микробов, у которых сила желтого свечения соответствовала таковой у контрольных бактерий с последовательностью AVST. Затем отобранных микробов помещали в среду, не содержащую магния. Выдержать такое «магниевое голодание» способны только бактерии с исправно работающей системой PhoQ-PhoP.
Бактерий, выдержавших эти испытания, подвергли секвенированию, чтобы получить полный список аминокислотных последовательностей, обеспечивающих нормальную работу PhoQ-PhoP. Таких последовательностей, позволяющих белку PhoQ нормально взаимодействовать с PhoP, оказалось неожиданно много: целых 1659. Авторы выборочно проверили некоторые из мутантных штаммов на способность конкурировать с контрольными кишечными палочками при разных концентрациях магния. Эти эксперименты показали, что приспособленность мутантов осталась на том же уровне, что и у исходного штамма. Таким образом, по крайней мере некоторые (а скорее всего, большинство) из выявленных 1659 последовательностей не имеют каких-либо серьезных дефектов по сравнению с исходной последовательностью AVST.
Это один из главных результатов обсуждаемой работы. Он показывает, что «плато» ландшафта приспособленности, соответствующее функциональному белок-белковому интерфейсу, весьма обширно: оно занимает более одного процента всего пространства последовательностей (1659 точек из 160 000). Это значит, что, подобно генетическому коду, в котором 20 аминокислот и знак «стоп» кодируются 64 триплетами, «белок-белковый» код, обеспечивающий узнавание белками друг друга, тоже обладает высокой избыточностью («вырожденностью»). Это, по идее, должно облегчать функциональную эволюцию белков. Чем обширнее плато ландшафта приспособленности, соответствующее данной функции, тем больше шансов, что с какой-нибудь его точки удастся перейти (или перепрыгнуть) на другое. Иными словами, повышается вероятность смены функции белка в ходе эволюции.
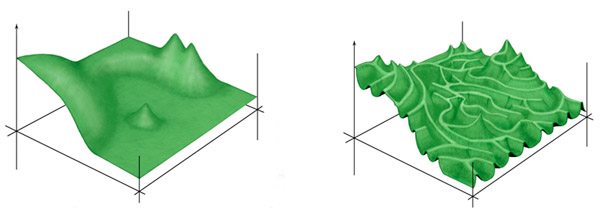
Полученные данные позволили авторам детально изучить топографию плато и оценить его «эволюционную проходимость» (рис. 1). Принципиальный вопрос, от ответа на который зависит общая динамика нейтральных (не полезных и не вредных, сохраняющих функцию белка) эволюционных изменений, то есть динамика перемещения эволюционирующей молекулы по плато ландшафта приспособленности, состоит в том, насколько сильным является влияние четырех аминокислотных позиций друг на друга. Генетики называют такое влияние «эпистазом». Если эпистаз отсутствует, то плато представляет собой ровную плоскую поверхность, по которой эволюционирующий белок может гулять абсолютно свободно. Есть некий набор «разрешенных» аминокислотных замен, и приобретать их можно в любом порядке. При сильном эпистазе плато становится похоже на лабиринт узких тропинок, разделенных пропастями. По такому плато передвигаться трудно, потому что аминокислотные замены, безвредные в одних контекстах, оказываются вредными в других. В результате порядок приобретения замен становится крайне важным: к каждой точке пространства последовательностей можно теперь добраться лишь строго определенными немногочисленными маршрутами. Следовательно, можно ожидать, что многие работоспособные последовательности до сих пор не найдены эволюцией просто потому, что у нее не хватило на это времени (подробнее см. в новости Расширение белковой вселенной продолжается, «Элементы», 24.05.2010).
Анализ полученного списка из 1659 работоспособных последовательностей показал сильнейшее взаимное влияние четырех аминокислотных позиций, то есть очень сильный эпистаз. Это второй главный результат исследования. В наличии эпистаза можно убедиться разными способами. Например, можно сравнить друг с другом последовательности, отличающиеся от AVST одной, двумя, тремя и четырьмя аминокислотами. Авторы обнаружили среди функциональных («разрешенных») последовательностей 16 одиночных мутантов, 100 двойных, 544 тройных и 998 четверных, то есть таких, в которых все четыре аминокислоты отличаются от исходных A, V, S и T. Если бы эпистаза не было, то разрешенные множественные мутации представляли бы собой просто-напросто произвольные комбинации одиночных. Однако ничего подобного не наблюдается. Каждая отдельная аминокислотная замена оказывается разрешенной в одних сочетаниях и запрещенной в других. Из-за этого кратчайший разрешенный эволюционный путь от одной последовательности к другой в большинстве случаев оказывается длиннее, чем кратчайший из потенциально возможных (рис. 3).

Рис. 3. Кратчайшие разрешенные эволюционные пути от AVST к последовательностям MLAI и SIQC. Функциональные (разрешенные) последовательности обведены рамками. Зеленым цветом показаны аминокислоты, отсутствующие в начальной и конечной последовательностях. Два левых рисунка в качестве единичного эволюционного события предполагают замену аминокислоты в белке, на правом рисунке таким событием считается замена одного нуклеотида в ДНК. В последнем случае путь получается длиннее из-за структуры генетического кода: от каждой аминокислоты путем замены одного нуклеотида можно прийти не ко всем, а лишь к некоторым из остальных 19 аминокислот. Рисунок из обсуждаемой статьи в Science
Рассмотрим, например, переход от AVST к разрешенной последовательности MLAI (рис. 3, вверху слева). Чтобы превратить AVST в MLAI, нужно заменить четыре аминокислоты. Соответственно, если считать аминокислотную замену единичным эволюционным событием, то кратчайший потенциально возможный путь от AVST к MLAI будет состоять из четырех шагов. Разных путей такой длины потенциально существует 24, поскольку аминокислоты можно заменять в разном порядке. Однако из этих 24 путей разрешенными являются только два. Все остальные пути требуют пересечения «пропасти» на ландшафте приспособленности, то есть прохождения через нефункциональный промежуточный этап.
Во многих случаях все кратчайшие пути оказываются запрещенными. Тогда маршрут из одной точки пространства последовательностей в другую (например, путь от AVST к SIQC, показанный на рис. 3 слева внизу) оказывается длиннее, чем кратчайший из потенциально возможных.
Ну а если учесть, что реальные единичные мутации обычно представляют собой замену одного нуклеотида в ДНК (а не триплета на любой другой триплет), то проходимость ландшафта окажется еще ниже. На рис. 3 справа показан кратчайший разрешенный путь от AVST к MLAI с учетом этого обстоятельства. Теперь он состоит не из четырех, а из целых десяти шагов. Это объясняется структурой генетического кода, которая позволяет путем единичных нуклеотидных замен переходить от данной аминокислоты только к некоторым, но не к любым другим аминокислотам. Например, фенилаланин кодируется триплетами UUU и UUC. Изменив в любом из этих триплетов только одну букву, можно заменить в белковой молекуле фенилаланин на лейцин, изолейцин, валин, серин, тирозин или цистеин. Для перехода к любой из оставшихся 13 аминокислот потребуется не одна, а две или три нуклеотидных замены. Таким образом, эпистаз и структура генетического кода совместными усилиями делают ландшафт приспособленности труднопроходимым.
На рис. 4 топография изученного плато показана в виде графа. Как видно, данное плато мало похоже на гладкое, легко проходимое поле. Оно подразделяется на несколько областей, внутри каждой из которых эволюционирующий белок может передвигаться относительно свободно, однако переход в соседнюю область возможен лишь по немногочисленным тропкам. Особенно хорошо это видно на нижнем рисунке, где ребра графа соответствуют нуклеотидным, а не аминокислотным заменам. Некоторые разрешенные последовательности вообще оказались недосягаемыми из «обжитой» области пространства последовательностей (изолированные кружочки на нижнем рисунке, к которым не подходит ни одна линия).

Рис. 4. Исследованная область пространства последовательностей в виде графа. Кружочки — разрешенные последовательности, соединяющие их линии — единичные аминокислотные замены (вверху) или единичные нуклеотидные замены (внизу). Размер кружка отражает число соседей, до которых можно добраться за один шаг. На верхнем рисунке видно, что изученная область распадается на пять подобластей (кластеров), внутри которых эволюционные перемещения могут происходить относительно свободно, тогда как переходы между кластерами затруднены. Буквенные диаграммы показывают частоту встречаемости аминокислот в каждой из четырех позиций для пяти кластеров. Высота буквы соответствует частоте встречаемости соответствующей аминокислоты: например, в кластере 1 первую позицию почти всегда занимает серин (S). На нижнем рисунке черным кружком обозначена последовательность AVST, характерная для E. coli, красными кружками — последовательности белок-белкового интерфейса PhoQ, встреченные у других гамма-протеобактерий. Оттенками синего показана вероятность добраться до данного кружочка из начальной точки (AVST) за 20 мутационных шагов. Рисунок из обсуждаемой статьи в Science
Авторы также проанализировали те варианты последовательностей из числа найденных ими 1659, которые реально встречаются у родственников кишечной палочки — других гамма-протеобактерий. Эти последовательности отмечены красными кружками на рис. 4 (внизу). Характерно, что все красные кружки расположены в той же самой «обжитой» области пространства последовательностей, что и AVST. Возможно, это означает, что нейтральная эволюция белок-белкового интерфейса действительно была настолько затруднена эпистазом и структурой генетического кода, что эволюция просто не успела «разведать» многие потенциально доступные области пространства последовательностей.
Данное исследование кому-то может показаться слишком скучным и специальным. Но только не биологам, привыкшим считать «пространство последовательностей» и «ландшафт приспособленности» абстракциями, полезными для теоретических построений, но практически недосягаемыми для реального изучения. Больше всего впечатляет сам факт, что стало возможным прямое экспериментальное картирование довольно обширных областей пространства последовательностей. Подобные пионерские исследования открывают путь к пониманию фундаментальных законов эволюции белков и к построению более точных эволюционных моделей. Хотя, конечно, речь пока идет лишь о четырех аминокислотных позициях, то есть о переборе 204 вариантов. Полное картирование пространства всех возможных белков навсегда останется неразрешимой задачей.
Источник: Anna I. Podgornaia and Michael T. Laub. Pervasive degeneracy and epistasis in a protein-protein interface // Science. 2015. V. 347. P. 673–677.
См. также:
1) Расширение белковой вселенной продолжается, «Элементы», 24.05.2010.
2) Пути эволюции предопределены на молекулярном уровне, «Элементы», 12.04.2006.
-
Мне кажется, что ландшафт приспособленности - слишком грубая абстракция первого приближения, годная только в качестве простой метафоры для понимания принципа. Во втором приближении эволюция ближе к векторному пространству (в одну сторону какому-нибудь признаку эволюционировать, как правило, "проще", чем в другую), а в третьем приближении векторные потоки возможных мутаций канализируются и получается направленный граф (с рёбрами ненулевого сечения ввиду стохастической природы представляемых величин).
Чем больше живой организм зависит от других живых организмов (а не от более статичных факторов неживой природы), тем более пространство его возможных мутаций канализируется и превращается в густую сеть ("губку") возможностей вместо непрерывного объёма. Плюс ко всему эта "губка" кипит, меняется из-за таких же "губок" других организмов, от которых зависит рассматриваемый.-
Ландшафт безусловно слишком грубое приближение, как минимум он сильно меняется при изменении отношений в экосистеме. Но тут есть интересный момент. Дело в том, что эволюция слепа. Каждый отдельный вид меняет себя в соответствии с мгновенным слепком ландшафта и не видит, что например, это ведёт к истощению экосистемы. Изменение отдельных видов идёт, как правило, сильно быстрее изменения экосистемы, так что самому виду рельеф в каждый момент эволюционного выбора кажется статичным.
Так что непосредственно изучая ландшафт при неизменных остальных параметрах мы узнаём достаточно точную информацию о том, как может меняться вид, хотя и не узнаём как от этого поведёт всю экосистему.
-
-
Ландшафт приспособленности - это как раз векторное пространство (в данном случае двумерное). Так что ваше второе приближение тождественно равно первому.
-
Ландшафт приспособленности - это скалярное поле, а не векторное. Конечно, можно в каждой точке двухмерной функции вычислить градиент, получив векторное поле, показывающее направление "выгодности" мутации с точки зрения приспособленности. Но в таком поле не будет возможным появление "циклонов", выражающих, например генетически обусловленные направления возможных мутаций.
-
Векторное поле и векторное пространство это не одно и то же.
---
Вы, видимо, имели в виду векторное поле. Но эволюция по ландшафту как раз моделируется хождением по скалярному полю методом случайного спуска (поиска). Вы ввели векторное поле представив эволюцию детерминированной. Тот самый градиент и образует Ваше векторное поле.
Не Вы один. В комментариях и "исходный код вселенной" и "замысел". "Природа хочет", короче говоря.
P.S. Извините, исправил после Вашего ответа.-
Нет, мы явно говорим о разных вещах, о "замысле" речи не шло, я лишь сказал, что у мутаций больше топологических ограничений, чем может выразить многомерная функция (даже если брать её градиент). В качестве примера, невыразимого градиентом я привёл циклически направленные вектора. Конечно, ландшафт приспособленности на то и приспособленности, что показывает только приспособленность, но тогда он имеет смысл только для конкретной "траектории" мутаций на этом же ландшафте (определённом только в окрестностях этой траектории), и при выборе точки за этой траекторией меняется и весь ландшафт. Т.е., ландшафт - это нечто "постфактумное", его невозможно построить заранее, до наблюдения за эволюцией конкретного вида.
-
-
-


-
Для диплоидных эукариот это штатный путь, но у бактерий две копии одного гена долго в клетке не сохраняются, так как у них обычно идет постоянный отбор на сокращение генома.
-
В принципе, у бактерий появление новых генов в результате дубликации уже существующих тоже довольно распространённое явление. Кроме того, многое зависит от интенсивности экспрессии конкретного гена, например, у большинства бактерий оперон, содержащий субъединицы рРНК, представлен несколькими копиями. В общем и целом, понятное дело, всё определяется соотношением силы давления очищающего отбора, длины пути, который нужно пройти, что бы цепочка мутаций оказалась эволюционно полезной, и степени преимущества, которое даёт сформировавшийся в итоге вариант гена.
Но глобально я согласен, обычно бактериям выгоднее получить новый ген в результате горизонтального переноса, чем пытаться "вывести" ген с аналогичными функциями своими собственными силами.-
Бактерии заимствуют гены извне только при сильном стрессе (антибиотики и т.п.), потому что такое редко кончается хорошо.
У людей может происходить подобное за счёт ДНК-полимеразы-тета, встраивающей в разрывы ДНК гены на основе РНК-матриц (своих, вирусов и т.д.) https://elementy.ru/novosti_nauki/433831/Odna_iz_chelovecheskikh_polimeraz_sposobna_provodit_obratnuyu_transkriptsiyu
-
-
-
Вы не так поняли: это не "спортивное ориентирование", а бегство от хищника по пересечённой местности (вариант пожара не предполагает бега кругами, часто применяемого паразитами и вирусами).
Другой вариант ответа https://elementy.ru/novosti_nauki/432406/Evolyutsiya_belkov_sderzhivaetsya_nizkoy_prokhodimostyu_landshafta_prisposoblen nosti?ofm=1#fm5307851
Надо отметить что из прошлого исследования 2006-ого года следует, что из найденных путей большая часть будет непроходима по причине того, что некоторые промежуточные шаги снижают приспособленность. Пройти по такому пути будет можно, но никто постарается туда не ходить.
В курсах или одиночный изогнутый овраг как иллюстрация сложности или две ямы в одномерном случае как иллюстрация нелокальных минимумов. В наше медийное время такие рисунки необычайно ценны.
Однако, для данной задачи такое приближение вызывает сомнение. Во-первых, пространство комбинаций аминокислот дискретно. Во-вторых, не факт, что поверхность гладкая. Скорее, она имеет вид столбчатой гистограммы с довольно хаотично меняющимися величинами. Да и параметр приспособленности очень условный.
Непонятно каким образом определяется соседство точек.
Хорошо бы проанализировать эту абстракцию на более простом примере. Скажем, органическая молекула (а не белок) с обозримым числом комбинаций. И параметр функции более однозначный. Например, суммарная энергия связи или что-то в этом роде. Вот тут можно построить математическую модель, которая бы прояснила что здесь лишь полёт фантазии, а что позитивная наука. Промежуточное исследование - пептиды.
-
Белки сложнее, потому что есть фолдинг, который зависит от других белков, локального pH и прочих условий. А ещё есть аффинность с теми же пептидами – "Неизвестные пептиды: «теневая» система биорегуляции" Чугунов 11.11.2010 https://biomolecula.ru/articles/neizvestnye-peptidy-tenevaia
-sistema-bioreguliatsii и "Возможности пептидов безграничны" интервью Наталии Лесковой с Вадимом Ивановым «ЗНАНИЕ — СИЛА» №1, 2020 https://elementy.ru/nauchno-populyarnaya_biblioteka/436186/V ozmozhnosti_peptidov_bezgranichny
Это объяснение почему так. В моменте действуют вариативность и отбор – статьи из "См. также" и https://elementy.ru/novosti_nauki/432406/Evolyutsiya_belkov_sderzhivaetsya_nizkoy_prokhodimostyu_landshafta_prisposoblen nosti?ofm=1#fm5307851
Последние новости











Рис. 1. Схематическое изображение «ландшафта приспособленности» в случае слабого (слева) и сильного эпистаза, то есть влияния одних мутаций на полезность или вредность других. Два горизонтальных измерения символизируют «пространство последовательностей», то есть все возможные варианты аминокислотной последовательности белка. Вертикальное измерение — эффективность выполнения белком своей функции. Если эффекты мутаций слабо зависят друг от друга, ландшафт оказывается легкопроходимым, что позволяет эволюции быстро освоить все потенциально доступные области. В случае сильного эпистаза ландшафт превращается в лабиринт тропинок, разделенных пропастями. В такой ситуации многие области пространства последовательностей могут остаться неразведанными эволюцией просто потому что до них трудно добраться. Рисунок Андрея Томиленко