Стереоинформация о запахе помогает кроту находить пищу
Не секрет, что сравнение как зрительных, так и слуховых сигналов, воспринимаемых симметричными органами чувств, помогает с легкостью определить расстояние до объекта или быстро локализовать источник звука. Но роль стереосигналов в нахождении источника запаха оставалась спорным вопросом. Новые исследования поискового поведения у восточно-американского крота подтвердили, насколько важно для этого животного двустороннее восприятие запаха. Особенно критично функционирование обеих ноздрей в пределах 4–5 см от источника запаха. На более далеких расстояниях крот пользуется в основном последовательным тестированием запаха в разных точках пространства.
Механизмы пространственной ориентации по запаху давно обсуждаются учеными. Предлагаются две гипотезы: либо последовательное тестирование запаха в разных точках пространства, либо одновременное сравнение запаха симметричными органами чувств. Последняя гипотеза кажется вполне правдоподобной, так как почти все животные имеют два симметричных сенсорных органа для восприятия запаха. Если животные обладают стереозрением и стереослухом, то логично предположить и наличие «стереозапаха». Удивительно, что в этой области не так много проводилось исследований. Работы, сделанные на крысах и человеке, показали, что блокирование одной ноздри приводит к ухудшению локализации источника запаха. Однако до сих пор почти не было исследований, в которых подробно бы исследовалась роль билатерального (двустороннего) сравнения в локализации источника запаха на основании градиента концентрации (то есть по направлению увеличения концентрации пахучих веществ, с максимумом в районе источника запаха).
Американский биолог Кен Катания (Kenneth C. Catania) из Университета Вандербильта (Vanderbilt University) обратил внимание, насколько быстро находит пищу восточноамериканский крот Scalopus aquaticus, который не может похвастаться ни хорошим зрением, ни развитым слухом. При поисках крот производит характерные нюхательные движения носом. Кен Катания специализируется на исследовании сенсорных систем животных-специалистов, у которых недоразвиты некоторые сенсорные органы, но гипертрофированно развиты другие, таких как голые землекопы или звездорылы. Крот S. aquaticus показался ученому подходящим объектом для исследования механизмов обнаружения источника запаха.
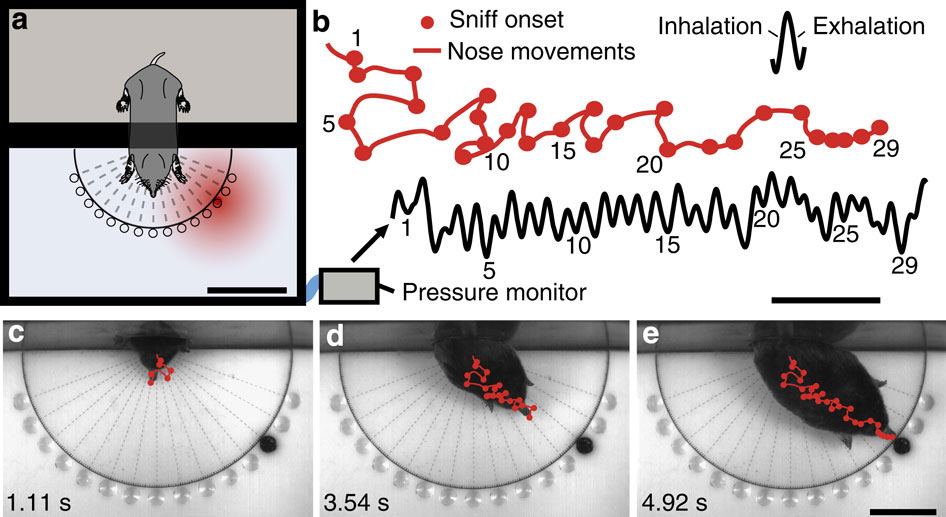
Для исследования нюхательного поведения крота в лаборатории было сконструировано несколько установок. Одна из них, так называемая радиальная арена, показана на рис. 1. Вначале животное помещали в отсек для передерживания, а после того, как оно успокаивалось, открывали отверстие в отсек для тестирования. В одной из 15 кормушек была помещена приманка (фрагменты дождевых червей), которую зверек должен был обнаружить (рис. 2а). В контрольных экспериментах животные быстро и безошибочно находили пищу. Хотя глаза у крота S. aquaticus недоразвиты и вдобавок закрыты мехом, часть экспериментов проводили при инфракрасном свете, к которому крот нечувствителен, чтобы полностью исключить участие зрительной системы в ориентации.
Затем животному временно блокировали левую или правую ноздрю пластиковой затычкой (эта процедура проводилась под анестезией), и эксперименты повторялись. Что же получилось? Животные стали ошибаться на 8–21°, причем эта ошибка всегда была в сторону открытой ноздри (рис. 2, b–f). Надо отметить, что животное после затыкания ноздри в конце концов находило приманку, но после более длительных поисков, чем интактное животное (на рисунках это не показано).

Рис. 2. Поведение крота Scalopus aquaticus на радиальной арене. (а) Примеры поведения двух интактных животных, разными цветами обозначены траектории нюхательных движений носа при движении к соответствующим кормушкам. (b) Примеры поведения двух животных с одной закрытой ноздрей (у одного левой, у другого правой). (c–f) Усредненные результаты по каждому из четырех животных (каждому давалось 10 попыток) показывают, что блокирование одной ноздри (красная стрелка) у одного животного приводило к ошибке в 21,6° (с), у другого — 11,6° (d), у третьего — 10,9° (e), у четвертого — 8,4° (f). (g, h) У двух дополнительных животных (им тоже давалось 10 попыток), которым вместо затычки вставили в ноздрю пустую трубочку (зеленая стрелка), точность поиска приманки была сравнима с таковой у интактных животных. Рисунок из обсуждаемой статьи в Nature Communications.
Для контроля автор заблокировал ноздрю крота не затычкой, а кусочком пустой пластиковой трубки (который свободно пропускал воздух и позволял чувствовать запахи), чтобы исключить просто влияние постороннего элемента в ноздре (это же может быть достаточно неприятно!). Но в этих контрольных экспериментах животные опять быстро находили приманку (рис. 2, g, h).
Следующая установка больше напоминала естественные условия, в которых обитают кроты. В туннеле кормушки были расположены рядами, и в зависимости от того, какая из них была заполнена кормом, расстояние от входа в туннель до источника корма варьировало (рис. 3а). Животные, у которых была блокирована одна ноздря, опять-таки ошибались в сторону открытой ноздри (рис. 3b). При этом они всё-таки находили корм, но после существенно более продолжительных поисков (рис. 3, с–f).

Рис. 3. Поведение крота Scalopus aquaticus в туннеле и Т-образном лабиринте. (а) Схема установки, каждый кружок обозначает кормушку. (b) Примеры траекторий нюхательных движений носа интактного животного (зеленый цвет), животного с закрытой левой ноздрей (красный) и животного с закрытой правой ноздрей (синий). (c–f) Усредненные результаты экспериментов, проведенных на четырех животных. Гистограммы показывают, что в случае блокирования левой ноздри животное подавляющее время тратило на поиски в правом секторе от кормушки, в случае блокирования правой ноздри — в левом секторе от кормушки. (g) Примеры траекторий нюхательных движений носа в Т-образном лабиринте животного с закрытой левой ноздрей. Рисунок из обсуждаемой статьи в Nature Communications
Автор предположил, что крот руководствуется не только билатеральным сравнением сигналов, но и сравнением сигналов в ходе последовательного тестирования запаха в разных точках пространства. Чтобы подтвердить эту гипотезу, животным предложили найти пищу в Т-образном лабиринте — условиях, максимально приближающихся к естественным (рис. 3g); в нём, чтобы найти пищу, кроту приходилось поворачивать налево или направо. На рисунке продемонстрированы траектории животного с заблокированной левой ноздрей, и видно, что животное с задачей неплохо справлялось. В целом, из четырех протестированных в Т-образном лабиринте животных одно животное находило приманку в 100% попыток, два — в 90% и одно — в 85%.
В ходе последующих экспериментов животным вставляли искривленные пластиковые трубочки в ноздри таким образом, что трубочка из левой ноздри была направлена вправо и, наоборот, трубочка из правой ноздри была направлена влево (эту операцию также проводили под анестезией). После такого «надругательства» бедные животные оказались абсолютно дезориентированы: они искали по обе стороны от источника пищи, причем поиски их были безуспешны и пищи они так и не находили. Результаты были сходны как на радиальной арене, так и в туннеле.
В процессе этих экспериментов автору удалось оценить критичное расстояние до источника запаха, когда начинает работать билатеральный механизм распознавания. Оно составило 4–5 см и менее. Когда животное с заблокированной ноздрей приближалось к кормушке на это расстояние, его начинало заносить вправо или влево от траектории, типичной для интактного животного, то есть его траектория больше отличалась от контрольной, чем на более далеких расстояниях.
Итак, проведенные эксперименты убедительно показывают, что двустороннее восприятие запаха играет большую роль в пространственной ориентации крота при поиске источника пищи. Возникает резонный вопрос, каким же образом сравниваются химические сигналы, проходящие через две ноздри. К сожалению, это практически не обсуждается в статье, возможно потому, что эта проблема пока мало исследована. Автор лишь сравнивает свои результаты с данными, полученными на слуховой системе сов. У последних блокирование одного уха приводило к систематической ошибке в ориентации в сторону открытого уха. Но механизмы локализации звука понятны: поскольку звук достигает одного уха раньше, чем другого, источник звука определяется по разнице задержки прихода сигналов и разнице интенсивностей звука. Если животному блокировать одно ухо, то такая ситуация может имитировать ситуацию, когда источник звука помещен со стороны интактного уха, и поэтому понятна ошибка в сторону открытого уха.
Но в случае локализации запаха возникает множество вопросов. Расстояние между ноздрями настолько мало, что трудно себе представить, каким образом оценивается разница между временем попадания летучего вещества на обонятельный эпителий правой и левой носовых полостей или количеством попавших в ту или иную ноздрю молекул. Непонятно, в каких единицах измерять интенсивность запаха и от чего она зависит. Несмотря на то, что многие свойства системы восприятия запахов можно объяснить на молекулярном уровне (см., например, В. Майоров «Восприятие запахов», «Наука и Жизнь» №2, 2007), остается неизвестным, где может происходить сравнение сигналов, попавших на эпителий правой и левой носовых полостей. В итоге можно заключить, что открывается огромное поле деятельности в области исследований молекулярных механизмов билатерального восприятия химических сигналов.
Источник: Kenneth C. Catania. Stereo and serial sniffing guide navigation to an odour source in a mammal // Nature Communications. V. 4. Статья №1441. Doi:10.1038/ncomms2444. (Статья в свободном доступе.)
Варвара Веденина


Рис. 1. Экспериментальная установка для исследования ориентации крота Scalopus aquaticus по запаху. (а) Камера из плексигласа разделена на два отсека, один для передерживания животного (верхняя часть на рисунке), а другой — для тестирования. Кружки обозначают потенциальные источники пищи. Длина масштабной линейки — 8 см. (b) Пример одновременной записи дыхания (черная траектория) и нюхательных движений носа (красная траектория) крота. Длина масштабной линейки — 1 с. (c–e) Примеры траекторий, черной точкой обозначен источник пищи. Длина масштабной линейки — 4 см. Рисунок из обсуждаемой статьи в Nature Communications