Какой же вклад протеинкиназа M-дзета вносит в формирование памяти?
Механизмы формирования памяти — одна из самых загадочных, интересных и важных областей нейрофизиологии, и в последние годы в ней произошел серьезный прорыв. Обнаружилось, что белок под названием протеинкиназа M-дзета (Protein kinase M zeta, PKMζ) критически необходим на одном из этапов возникновения памяти. Об этой многообещающей молекуле, новых работах в данной области и возможных перспективах ее развития рассказал в своей лекции на проходящей при поддержке РВК, Фонда «Династия» и РФФИ Зимней школе Future Biotech директор Института высшей нервной деятельности и нейрофизиологии Павел Милославович Балабан.
Если мы рассматриваем память как физиологическое явление, то особенное внимание нам нужно сосредоточить на синапсах — местах контакта между двумя нервными клетками. Можно представить себе такую модель, в которой запоминаемое событие каким-то образом «записывается» в определенном синапсе, облегчая прохождение через него сигнала. В результате связь между двумя нейронами становится более прочной — что и лежит в основе запоминания.
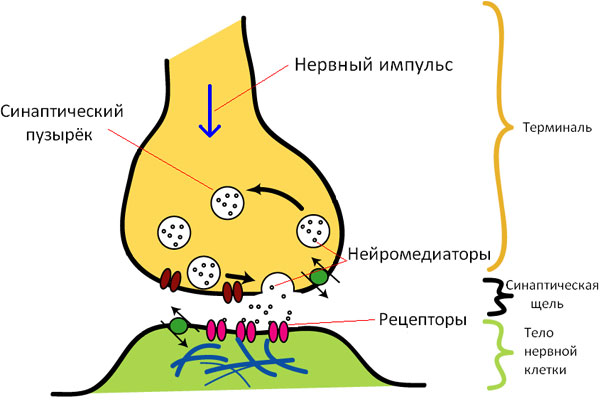
Передача сигнала через химический синапс — это, по сути, просто выброс в синаптическую щель пузырьков с нейромедиатором в ответ на потенциал действия, пришедший по мембране аксона (рис. 1). Медиатор «садится» на соответствующий рецептор постсинаптической мембраны, и в зависимости от того, что это за медиатор и какие рецепторы к нему находятся на мембране, в постсинаптическом нейроне происходят те или иные изменения, в том числе потенциал его мембраны уменьшается (это называется деполяризацией или возбудительным постсинаптическим потенциалом, ВПСП) или увеличивается (это гиперполяризация, или тормозный постсинаптический потенциал, ТПСП). Величина ПСП зависит от площади синапса, количества выброшенного медиатора, количества рецепторов к этому медиатору и так далее. ПСП возникают локально вокруг синапса и распространяются на некоторое расстояние от него, постепенно затухая. Они суммируются между собой в пространстве и времени: например, ВПСП и ТПСП «погасят» друг друга, а два ВПСП, наоборот сольются в ВПСП большей амплитуды.
При этом, если в том участке нейрона, где начинается аксон (этот участок называется аксонным холмиком) мембранный потенциал деполяризуется больше некоей критической величины, то по аксону побежит нервный импульс, то есть уже этот нейрон передаст сигнал какому-то следующему нейрону.
Таким образом, приведет ли активация синапса к генерации импульса постсинаптическим нейроном, зависит от множества вещей: от того, насколько близок синапс к аксонному холмику; какие сигналы пришли на окрестные синапсы; сколько медиатора было выброшено; сколько и каких рецепторов к нему находилось на постсинаптическом нейроне; какая часть медиатора все же добралась до рецепторов, а какая была бесславно метаболизирована или чуть менее бесславно обратно проглочена пресинаптическим нейроном; какая метаболическая ситуация была в тот момент в пре- и постсинаптической клетке и так далее, и тому подобное.
Хорошо изучено, что в определенных условиях сила синапса (Synaptic strength), то есть то влияние, которое он оказывает на постсинаптическую клетку, может уменьшаться или увеличиваться. Это явление называется синаптической пластичностью. В основе синаптической пластичности лежит несколько различных механизмов, но для нас сейчас важнее всего тот, который имеет непосредственное отношение к памяти и называется долговременной потенциацией.
Долговременная потенциация — это увеличение силы синапса в случае его частого использования. Иными словами, каждое последующее возбуждение синапса вызывает более сильный ответ постсинаптического нейрона, чем предыдущее. То есть, очень упрощая, можно представить себе такую схему, объясняющую формирование памяти с помощью долговременной потенциации. В результате события Х через какой-то синапс прошел сигнал и вызвал ответ постсинаптического нейрона (а именно, изменение его мембранного потенциала). Когда событие повторилось, то прохождение сигнала через тот же синапс вызвало большее изменение потенциала. Значит, нейрон как будто «запомнил» этот синапс и увеличил его значимость, что привело к запоминанию события Х целым организмом.
Долговременная потенциация обеспечивается множеством метаболических каскадов, в которые вовлечено буквально астрономическое количество белков, и выловить в этом океане какую-то конкретную молекулу, которая непосредственно вовлечена в формирование памяти и без которой память «выключится», казалось нерешаемой задачей. Однако постепенно стали появляться доказательства того, что существуют некие белки памяти, которые начинают экспрессироваться в постсинаптической клетке в ответ на возбуждение синапса, подплывают к нему и усиливают его ответ на последующие импульсы, увеличивая количество и активность постсинаптических рецепторов. При этом данные белки должны экспрессироваться постоянно, иначе они быстро развалятся, долговременная потенциация прекратится и никакая память формироваться не будет.
Тот факт, что процессы запоминания и воспоминания связаны с работой белков памяти, подтверждает недавно опубликованная работа (Kaycie K. Tayler et al, 2012. Reactivation of Neural Ensembles during the Retrieval of Recent and Remote Memory), в которой с помощью элегантных методик флуоресцентного мечения показано, что из тех корковых нейронов, которые возбуждаются в ответ на первое предъявление некоего стимула (запоминание), и из тех, которые возбуждаются в ответ на его последующие предъявления (воспоминание), большой процент совпадает. Иными словами, запоминание и воспоминание происходят в одном и том же месте, и, вероятно, с помощью одних и тех же молекул — то есть белков памяти.
Обнаружилось несколько белков, которые можно было отнести к белкам памяти, однако каждый из них оказывался связан не только с памятью, но и с чем-то еще — то есть обеспечивал память не прямо, а опосредованно. Поиски конкретной молекулы памяти стали казаться поисками черной кошки в темной комнате, которую невозможно найти, потому что ее там нет.
И вот в 2007 году произошел прорыв: обнаружилось, что если в постсинаптическом нейроне заблокировать с помощью блокатора под названием ZIP короткий белок протеинкиназу Мζ (PKMζ), то у подопытных животных нарушается только память и (судя по нынешним данным) ничего кроме памяти (Reut Shema, Todd Charlton Sacktor, Yadin Dudai, 2007. Rapid Erasure of Long-Term Memory Associations in the Cortex by an Inhibitor of PKMζ) Иными словами, именно этот белок обеспечивал один из этапов возникновения памяти.

Рис. 2. «Молекула памяти» PKMζ. Картинка из слайдов к лекции П. М. Балабана на Зимней научной школе Future Biotech
«Молекула памяти» PKMζ стала бурно изучаться. Ее высокая консервативность (то есть то, что у разных видов животных аминокислотная последовательность этого белка почти не отличается) намекает на участие в какой-то важной и базовой функции. А постоянная экспрессия PKMζ обеспечивается следующим изящным механизмом: PKMζ «ловит» с помощью некоторых молекулярных каскадов свою собственную матричную РНК и таким образом синтезирует новую молекулу PKMζ, которая повторяет процесс. Таким образом появляются все новые и новые молекулы PKMζ. В результате PKMζ может сохраняться в синаптической области практически вечно (хотя на деле такое может не произойти, потому что в клетке может измениться метаболическая ситуация: например, кончиться белки, необходимые для поддержания уровня PKMζ).
Конечно, PKMζ не отвечает за всю память целиком, она необходима только на определенном этапе и появляется в области синапса примерно через два часа после прохождения через него сигнала. Это означает, что существуют еще другие, наверняка многочисленные и сложные, молекулы памяти. Но это ниточка, потянув за которую, можно распутать весь механизм памяти. Кроме того, PKMζ может оказаться удобной мишенью для лекарств, улучшающих память.
Однако не всё так просто. Некоторые данные прямо противоречат такой жестко определенной роли PKMζ в процессах памяти. В частности, в двух статьях, только что опубликованных в журнале Nature (Lenora J. Volk et al, 2013. PKM-ζ is not required for hippocampal synaptic plasticity, learning and memory; Anna M. Lee et al, 2013. Prkcz null mice show normal learning and memory), утверждается, что ни PKMζ, ни ее «родственница» PKСζ не имеют отношения к памяти, а все ранее исследованные эффекты связаны просто с тем, что использованный в прежних работах блокатор, ZIP, не обладает достаточной избирательностью. Иными словами, ZIP помимо PKMζ блокировал что-то еще, и именно это «что-то» и было связано с памятью. Однако если PKMζ не имеет отношения к памяти, то непонятно, что делать со многими другими данными, например с теми, по которым искусственное увеличение уровня PKMζ приводит к улучшению запоминания у крыс (Reut Shema et el, 2011. Enhancement of Consolidated Long-Term Memory by Overexpression of Protein Kinase Mζ in the Neocortex). То есть вопрос с вовлеченностью PKMζ в формирование памяти остается открытым.
Еще одна молекула, которая, видимо, имеет отношение к процессам памяти (а точнее, забывания) — это нитроксид, NO. Физиологи терпеть не могут эту молекулу, потому что она настолько деятельна и вездесуща, что, блокируя ее, получаешь эффекты всегда — только вот с какой именно деятельностью NO эти эффекты связаны, как правило, остается загадкой. В то же время, ряд данных говорит о том, что нитрозилирование с помощью NO многих белков (и в том числе, видимо, PKMζ) приводит к стиранию памяти. Эксперименты, проведенные на виноградных улитках в лаборатории П. М. Балабана, показали, что если убрать из организма NO, то даже общая блокировка синтеза белков не приведет к исчезновению памяти (Балабан П. М., Коршунова Т. А., 2011. Сетевые, клеточные и молекулярные механизмы пластичности в простых нервных системах). Конечно, все полученные на NO результаты стоит принимать с изрядной долей скепсиса, однако определенную пользу извлечь из них всё же можно.
И наконец, есть другие возможные кандидаты в «молекулы памяти» — прионоподобные белки. Как и прионы, они имеют две конформации — нормальную и патологическую, причем стоит только одной молекуле прионоподобного белка перейти в патологическую конформацию, как все соседние молекулы такого белка сразу же тоже эту конформацию приобретают. Но в отличие от прионов, у прионоподобных белков патологическая конформация не приносит вреда клетке — просто, раз в нее перейдя, прионоподобные белки так навсегда в ней и остаются. Такой конформационный переход выглядит очень соблазнительно для нейрофизиолога, занимающегося молекулярными механизмами памяти: ведь «перещелкивание» прионоподобного белка в новую конформацию может как раз и обеспечивать запоминание, то есть навсегда метить запомнившие что-либо синапсы. Определенные подтверждения того, что прионоподобные белки действительно имеют отношение к памяти, уже получены (Amitabha Majumdar et al., 2012. Critical Role of Amyloid-like Oligomers of Drosophila Orb2 in the Persistence of Memory). При этом интересно, что некоторые молекулярные каскады таких прионоподобных белков, судя по всему, связаны с деятельностью PKMζ — то есть PKMζ, прионоподобные белки и NO могут оказаться звеньями одной цепи, обеспечивающей память.
Одним словом, проблема белков памяти еще таит в себе множество тайн, и может быть, последующие исследования позволят разгадать хотя бы некоторые из них.
Источник: лекция П. М. Балабана «Молекулярные механизмы памяти», прочитанная на Зимней научной школе Современная биология & биотехнологии будущего.
См. также:
1) Greg Villareal, Quan Li, Diancai Cai, Ann E. Fink, Travis Lim, Joanna K. Bougie, Wayne S. Sossin, and David L. Glanzman. Role of Protein Kinase C in the Induction and Maintenance of Serotonin-Dependent Enhancement of the Glutamate Response in Isolated Siphon Motor Neurons of Aplysia californica // The Journal of Neuroscience. 22 April 2009. V. 29. P. 5100–5107.
2) Peter Serrano, Eugenia L. Friedman, Jana Kenney, et al. PKMζ Maintains Spatial, Instrumental, and Classically Conditioned Long-Term Memories // PLoS Biol. V. 6(12): e318. doi:10.1371/journal.pbio.0060318.
3) Hideaki Ogasawara, Tomokazu Doi, Mitsuo Kawato. Systems Biology Perspectives on Cerebellar Long-Term Depression // Neurosignals. 2008. V. 16. P. 300–317.
4) Todd C. Sacktor. Memory maintenance by PKMζ — an evolutionary perspective // Molecular Brain. 2012. V. 5. P. 31.
5) How does PKMζ maintain long-term memory? // Nature Reviews Neuroscience. V. 12. P. 9–15.
Вера Башмакова
-
Кратковременная память явно работает как-то по-другому. Ведь я помню то, что произошло секунду назад, какие "многочисленные и сложные, молекулы памяти" могут успеть синтезироваться за такое время?
-
Да-да, Вы правы, PKMζ отвечает только за достаточно поздний этап формирования памяти и синтезируется только примерно через 2 часа после прохождения сигнала, о чем в тексте и написано. Ранняя память обеспечивается немного другими механизмами, может, как-нибудь и напишу об этом, если там откроют что-нибудь новенькое.
-
Данные по основе кратковременной памяти (ее субъективизированной форме, доступной осознанию) более полувека назад систематизировала О.Виноградова в книге "Гиппокапм и память", а не так давно механизм был описан А.Иваницким, например, в статье "Сознание и мозг". Так что можно не ждать "чего-то новенького" :)
-


-
Вряд ли синапс -- это бит. Поскольку уровень потенциации может меняться в широких пределах, его можно аппроксимировать типом float или double, то есть, это 4 или 8 байт. Потом в мозгу вроде бы не 10, а 100 млрд. нейронов. Наконец, мозг запоминает не в raw формате, а образами, что, видимо, напоминает сжатие MPEG/JPEG или DJVU, то есть, информации запоминается ещё в 10-100 раз больше, что даёт 80 петабайт.
Но это, конечно, нумерология, ведь мы толком не знаем, в каком формате происходит запоминание.-
Не, ссылки на аналоговые вычисления тут не помогут. В лучшем случае аналоговость может превратить бит в байт. И то вряд ли. Клетки это все-таки не прецизионные измерительные приборы. Аналогия с DJVU интересна, но тоже вряд ли может обеспечить прирост более чем на порядок. А сама оценка с меньшей точность.
Меня смущает терабайт потому что кажется что это мало. Хотя кто его знает. Может быть, мы и вправду не шибко много помним. Если же нет, то синапс это не все, должен быть более емкий механизм.-
Ну смотрите: в глазу имеются "оптические" нейроны, которые называются колбочки и палочки. Экспериментально установлено, что true color это 32 бита на тетраду (3 колбочки + 1 палочка). Значит, каждый нейрон по крайней мере способен передавать байт, а не бит.
В любом случае, информация в мозгу сильно заархирована, как и в геноме. В геноме уж точно содержится всего 10 мегабайт -- а это кодирует всё человеческое тело!-
Архивация это другой вопрос. Емкость это то, сколько битов может быть сохранено. Например, купили вы гигабайтный диск, у вас гигабайт. Независимо от того архивирована ваша информация или нет, и вообще, записана ли она или диск пустой.
Вопрос в том, сколько есть физических элементов для сохранения информации.
-
-
-
-
Кстати, есть же организмы с количеством нервных клеток, которые чуть ли не по пальцам можно пересчитать. Как у них обстоят дела с памятью?
-
В экспериментах на ИЗОЛИРОВАННОЙ нервной клетке ещё в 1968-1970 годы было продемонстрировано, что нейрон НЕ ИМЕЮЩИЙ НИКАИХ СИНАПСОВ обладает способностью а) вырабатывать оптимальную линию поведения при взаимодействии с искусственной внешней средой 2)обучаясь, запоминать тактику поведения, причёим после нескольких сеансов скорость нахождения оптимальной тактики поведения снижалась с 30 минут до несколь ких секунд. Подробности см.Б.Г.Режабек О поведении механорецепторного нейрона в условиях замыкания его цепью искусственной обратной связи. ДАН СССР, 1971, т. 196, вып.4. С. 981-984
1) связь одного нейрона с другим (две ссылки объёмом, в Вашем примере, 10 000 000 000 вариантов, что укладывается в 34 бита минимум)
2) как минимум один коэффициент передачи сигнала (положим, ещё пара-другая байт числа, плюс один бит на знак, потому что есть и синапсы, тормозящие активность нейрона, а не только возбуждающие его). А то и не один коэффициент - ибо импульсная передача сама по себе обладает рядом характеристик, обусловленным текущим химическим составом синапса - порог и задержка срабатывания, скорость возрастания и убывания импульса, его форма....
3) Кроме того, каждый синапс имеет своё состояние развития (то есть, можно сказать, хранит информацию о нём) - чтоб меняться со временем и обстоятельствами. Тут мне тоже не очень понятно, сколько бит потребуется, чтоб все значимые характеристики описать...
5) вокруг синапсов и нейронов находится питательная среда межклеточного пространства, множество клеток нейроглии, капиляры... Они все обеспечивают условия функционирования синапса, влияют не его рост, функционирование, восстановление после каждого пробоя импульсом. И всё это, как и он сам, динамически меняется и взаимозависит... Возможно, их влияние может быть описано в терминах внутреннего состояния синапса (из предыдущего пункта), но возможно придётся добавить ещё что-то.
4) кроме того, есть текущее напряжение на синапсе, которое постоянно меняется - нервная активность живой нервной сети - это тоже надо учитывать как обрабатываемую синапсом информацию, и она постоянно меняется - это непрерывный аналоговый поток, типа аудиоканала. С какой частотой дискретизации его записывать, и с какой глубиной? 16 бит? Double? Наверно, как звук пишут, так и здесь - в конце концов, звук пишут чтобы мы его слышали, и значит 44 килогерц должно хватать и нервам... В любом случае, немалая часть информации хранится не в синапсах, а в циркулирующих по разным маршрутам мозга цепочках таких импульсов. И это, несколько я понимаю, не только кратковременная память.
И это только о синапсах - а ведь порог срабатывания и внутреннее состояние есть и у самих нейронов, к тому же у них есть характеристики потерь и скорости сигнала на аксоне и дендритах, зависящие от их длины, защищённости миелином и маршрута.
В общем, тут явно терабайтом не обойтись. Ни одним, ни десятком.
Последние новости







Рис. 1. Структура синапса. Изображение с сайта ru.wikipedia.org