Чужеземные растения позже сбрасывают листья, извлекая из этого немалую пользу

Сроки осеннего опадания листьев у кустарников и лиан, образующих подлесок в лесах восточной части США, значительно отличаются в зависимости от того, местные это виды, исконно произрастающие в данной области, или чужеземные, завезенные (по большей части непреднамеренно) из Евразии. Виды местные сбрасывают листья вскоре после того, как опадают листья деревьев верхнего яруса. Виды чужеземные теряют листья в среднем на 28 дней позже. Всё это время они продолжают активно фотосинтезировать, используя свет, который более не задерживается пологом леса. Данное обстоятельство, безусловно, способствует процветанию заносных видов в новых для них районах.
Успех, которого достигают многие растения, попавшие в новые для них места обитания, нередко удивляет исследователей и нуждается в специальном объяснении. В ряде случаев преимущество заносных видов по сравнению с местными связано с тем, что они менее подвержены заболеваниям, вызванным новыми для них вирусами, грибами и бактериями. Соответственно, ресурсы, ранее затрачиваемые на защиту от паразитов, теперь могут быть перенаправлены на рост и размножение (см.: Растения-вселенцы процветают за счет освобождения от паразитов и умелого использования ресурсов, «Элементы», 25.06.2009; Почему некоторые растения на чужбине растут лучше, чем на родине?, «Элементы», 25.05.2009).
Однако, скорее всего, существуют и другие механизмы, способствующие успешному произрастанию растений в новых местах обитания. Так, наблюдения за растениями подлеска в листопадных лесах восточных районов США показывают, что у чужеземных (завезенных из Евразии) видов листья весной распускаются раньше, а осенью сбрасываются позже, чем у видов местных. Благодаря этому заносные растения полнее используют необходимый для фотосинтеза свет в те промежутки времени, когда деревья, образующие основной полог, стоят без листьев. Однако одних только наблюдений, без строгой количественной оценки различий между местными и заносными видами, недостаточно, чтобы уверенно говорить о наличии подобного механизма обеспечения преимущества чужеземцев.
Поэтому столь важны опубликованные недавно результаты исследований Джейсона Фридли (Jason Fridley) из отдела биологии Сиракьюсского университета (см.: Fridley Lab at Syracuse University; Сиракьюс, штат Нью-Йорк, США). В течение трех вегетационных сезонов Д. Фридли проводил эксперименты со специально высаженными растениями (кустарниками и лианами, образующими подлесок в листопадных лесах восточных районов США). Он сравнивал сроки распускания и опадания листьев разных видов, а также оценивал в разные моменты вегетационного сезона количество хлорофилла в листьях и непосредственно интенсивность фотосинтеза.
Для стандартизации условий оценки всех параметров в 2006–2007 годах в окрестностях Сиракьюса (43°03' с. ш., 76°09' з. д.) были заложены небольшие площадки, куда высаживали растения 73 видов (30 завезенных и 43 местных). Всего было три одинаковые площадки (три повторности). Растения (кустарники и лианы) брали преимущественно из местных лесов, но некоторые — из дендрологических питомников. Виды подбирались так, чтобы завезенные и местные по возможности были близкородственными, относящимися к одному роду или, по крайней мере, семейству. Для воспроизведения эффекта затенения верхним пологом леса над растениями натягивали тент, перехватывающий 80% света. Устанавливали затенение только на срок закрытия верхнего полога в окрестных лесах (примерно с 20 мая по 24 октября). Эксперимент длился три вегетационных сезона подряд — с 2008-го по 2010 год.

Сезонные изменения «хлорофилльного индекса» (показателя, по которому можно судить о количестве хлорофилла, а косвенно — и об интенсивности фотосинтеза) для видов растений, являющихся исконными обитателями данной местности (Natives, зеленые линии) или завезенных из Евразии (Non-natives, красные линии). Сплошные линии соответствуют «световым листьям», а пунктирные — «теневым». Диаграмма внизу — распределение дат, в которые была утеряна половина хлорофилла (зеленым цветом показаны местные виды, красным — заносные). Серым цветом выделен период времени, соответствующий наличию листвы в верхних ярусах (в данном случае — затенения, не пропускающего 80% света). Хорошо видно, что и весной, и — особенно — осенью заносные виды гораздо полнее используют свет, не задерживаемый основным пологом леса. Изображение из обсуждаемой статьи в Nature
Весеннее распускание листьев оказалось очень чувствительным к конкретным погодным условиям того или иного года. Так, в особо теплом 2010 году почки у всех растений распустились раньше, чем обычно. Что касается средних сроков появления первых листьев у заносных и местных растений, то они различались незначительно, хотя местные виды всё же отставали от заносных. Однако осенью заносные виды теряли листву существенное позже, чем местные. Если медиана распределения дат потери 50% листьев для всех местных видов в течение 2008–2010 годов приходилась на 16 сентября, то для чужеземных — на 13 октября. Окончание листопада, точнее момент потери 90% листьев, для местных видов в среднем попадал на 27 октября, а для заносных — на 9 ноября. В среднем за три года наблюдений вегетационный сезон заносных видов оказался на 29 суток длиннее, чем у местных.
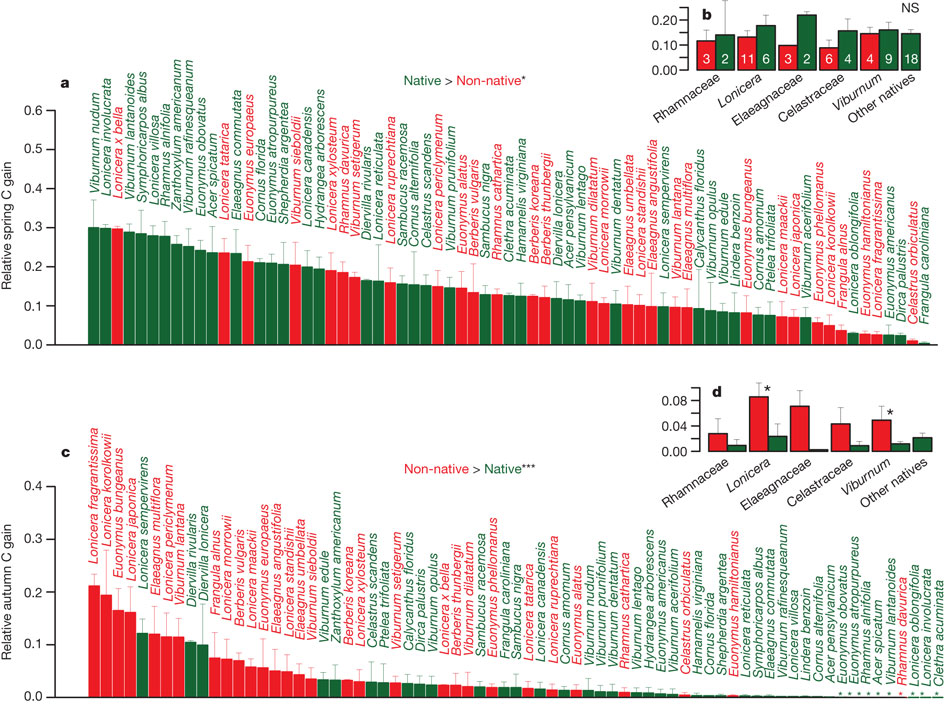
Разные изученные виды подлеска, ранжированные по убыванию той доли связанного за вегетационный сезон углерода, которая приходится на весенний период, до смыкания полога леса (верхний ряд, a), и на осень, после открытия полога (нижний ряд, c). Красным цветом показаны заносные виды (Non-native), зеленым — местные (Native). Если весной, до смыкания полога, местные и заносные виды фиксируют примерно одинаковое количество углерода, то осенью (после открытия основного полога леса) заметный фотосинтез продолжается в основном только у заносных видов. В правой части рисунка на панелях b и d приведены данные, суммированные по семействам растений (красным и зеленым цветом также обозначены заносные и местные виды). Видно, что весной заносные и местные виды из одного семейства значимо не отличаются по относительной величине фотосинтеза, но осенью заметный фотосинтез идет только у заносных видов, а не у местных из того же семейства. Изображение из обсуждаемой статьи в Nature
Само по себе более позднее опадание листьев у заносных растений еще не свидетельствует о том, что осенью они продолжают фотосинтезировать, извлекая из этого для себя заметную пользу. Однако исследования Д. Фридли показали, что у заносных видов, в отличие от местных, продолжается активный фотосинтез. Более того, исследователь рассчитал для каждого конкретного вида растений, какая доля от общего количества углерода, связанного (в виде СО2) за весь вегетационный сезон, приходится на весну (до смыкания верхнего полога леса), а какая — на осень (после опадения листьев верхнего полога). Обнаружилось, что если весной местные и заносные виды примерно в равной степени используют имеющий в их распоряжении свет, то осенью заметную выгоду получают почти исключительно заносные, но не местные виды.
Источник: Jason D. Fridley. Extended leaf phenology and the autumn niche in deciduous forest invasions // Nature. 2012. V. 485. P. 359–362.
Алексей Гиляров
-
Вот интересно, если сохранение листьев так выгодно, почему среди местных растений отбор не выработал эту способность? Или это выгодно только в краткосрочной перспективе, а на более длинных промежутках времени это пагубно -- допустим, раз в несколько поколений особо ранние морозы прореживают слишком долго зеленеющие растения?
-
Это очень даже резонный вопрос. Остается только предполагать! Возможно как раз по причине, указанной Вами.
-
Меня больше интересует вопрос как листопадным растениям в лесу удается сопротивляться листопаду? Листопад вызывается гормоном отмирания/созревания - этиленом. И именно из-за массового его выделения в лесах столь внезапны и мгновенны листопады. Еще вчера дерево стояло зеленом назавтро желтое к концу недели полуоблетевшее. Хотя может быть еще и тепло и светло и влажно. Отдельно стоящим деревьям еще удается сопротивляться этилену но в лесу все просиходит дружно. Вот и вопрос как заносные виды не только уходят в темный и холодный период с листвой но и переживают массовую этиленовую атаку?
-
Могу предположить что для разных видов требуется разный уровень этилена. Ведь рядом растущие клен и береза все равно опадут каждый в свое время.
-
да нет кленово-березовые рощи опадают как раз дружно. Думаю на самом деле вопрос в том что это растения полога, которым приходится расти всего 2 раза за год весной и осенью, поэтому умение сопротивляться этилену у них возможно есть. Не уверен просто гипотеза.
-
Вы, что же всерьёз думаете, что концентрация этилена в лесу сравнима с коцентрацией в камерах дозревания для томатов и бананов? В растениях это следовые концентрации. А для запуска механизма листопада важен индивидуальный баланс синтеза ауксинов. Что же касается работы, то создаётся впечатление, что местные виды успешно завершают фенофазу, а слегка придавленным "пришельцам" остаются крайне непрогнозируемые "крохи с барского стола". Можно ли это считать "процветанием" покажет будущее.
-
Я не думаю а пишу на основании прочитанных работ. А в камерах концентрация если вы знаете какая тоже весьма не большая, да и гормоны тем и отличаются что действуют именно в следовых количествах. Знаете что в промышленных холодильниках обязательны системы контроля содержания этилена, чтобы фрукты овощи не гнили? А это не камера дозревания а просто холодильники. А что с синтезом ауксинов? Они управляют ростом корневой и совместно с кинетинами обуславливают рост растения в целом. Отмиранием или состоянием покоя заведуют этилен, гидроксикоричные кислоты и эпибрассолиды. Чтобы ауксин вызывал опадение листьев слышу впервые. Вы ничего не перепутали? Напротив есть даже методика пробуждения спящих растений с опавшими листьями обработкой гетероауксином, что вызывает рост корней, ответный синтез кинетина и пробуждение спящих почек.
Про процветание вопрос согласен спорный.-
Нет, не очень перепутал:) Ауксины во многих случаях определяют скорость синтеза этилена. У быстро растущих растений могут возникать проблемы с удалением избытка этилена, особенно, если листья подтоплены или очень сильно затенены. Все искусственные аналоги ауксинов в высокой концентрации - дефолианты или гербициды (вспомните, хотя бы, Agent Orange). Ауксины (вернее их баланс)определяют формирование и свойства слоя клеток, отделяющих черешок листа от побега. Так что листопад - сложный мультифакторный внутренний процесс, а не просто газовая атака. При несомненной важности этилена в процессах созревания и отмирания, сомневаюсь, чтобы слаборастворимый в воде газ при его естественной эмиссии в ПРИРОДЕ действовал бы на соседние растения. Да и зачем, с эволюционной точки зрения, такая репетиция апокалипсиса?
-
Про дефолианты читал, там эффект совсем другой. Снижение уровня синтеза ауксинов приводит к формированию отделительного слоя. А что ее вызывает? Воздействие этилена и отмирание листа. Так что ауксины как минимум вторичны в этом процессе. Хотя и участвуют в собственно опадании листа. И лишь потом когда пектиназу и целлулазы отделительный слой разрушат лист опадет. Но не листопадные растения этилен так же действует.
Еще раз повторяю проведите классический опыт описанный во всех учебниках ботаники - поместите ветку листопадного дерева в стакане с водой под колпак с яблоком, за неделю все листья опадут.
С эволюционной точки зрения такой процесс несомненно выгоден позволяет всему сообществу разом уйти в состояние покоя, никто не рискует замерзнуть никто не получает преимуществ от продления активной фазы, листогрызы и листовые болезни разом уничтожаются а не накидываются на оставшихся. А растворимость этилена в воде нормальная, не хуже других. Например в 4 раза лучше чем кислорода и всего в 10 раз хуже чем у СО2 который считается хорошо растворимым. Знаете форму применения этилена в теплицах? Это именно этиленовая вода - вода, через которую пропустили этилен. Ей просто поливают растения если нужен эффект, например бромелиевые сразу зацветают.
И не надо пренебрегать действием этилена через воздух известно что акации в африке синтезирует токсины именно под действием этилена передающемуся по ветру. Травоядные обьедают 1 дерево, оно выделяет этилен в ответ на повреждение, и начинает синтезировать токсин ядовитый для травоядных, под действием этилена все соседние акации начинают синтезировать токсины тоже. Вот вам и пример этиленовой атаки.
Вроде ответил на все ваши вопросы?
-
-
-
-
-
-
-
-


Последние новости






Листопадный лес осенью. Парк Гарримана (Harriman State Park), штат Нью-Йорк. Фото Duncan N. L. Menge с сайта nceas.ucsb.edu