Скрытая изменчивость помогает приспосабливаться к новым условиям

Многие мутации не влияют на приспособленность организма и поэтому могут пассивно накапливаться в генофонде, формируя запас «скрытой изменчивости». Предполагается, что скрытая изменчивость помогает популяции выжить при изменении среды, потому что некоторые из накопившихся «бесполезных» мутаций могут оказаться полезными в новых условиях. Швейцарским биологам удалось подтвердить это предположение в экспериментах с молекулами РНК, эволюционирующими в пробирке. Популяции молекул, которым дали возможность накопить «скрытую изменчивость», приспособились к новой среде лучше и быстрее, чем контрольные популяции без скрытой изменчивости.
Живые существа, как правило, обладают высокой «помехоустойчивостью». Это значит, что многие случайные «помехи» (внутренние — мутации, или внешние — колебания условий среды) тем или иным способом компенсируются и не приводят к изменению фенотипа или снижению приспособленности. Естественный отбор поддерживает развитие адаптаций, повышающих устойчивость фенотипа к «помехам», в частности, к потенциально вредным мутациям. Примером такой адаптации могут служить белки-шапероны (см.: Когда вредных мутаций много, они не так вредны, «Элементы», 06.12.2005).
Помехоустойчивость ведет к тому, что в популяции свободно накапливаются мутации, которые до поры до времени не оказывают заметного влияния на приспособленность (остаются «нейтральными») или даже вовсе не проявляются в фенотипе. Но если какой-то из механизмов, обеспечивающих помехоустойчивость, выйдет из строя (из-за сильного внешнего воздействия или мутации в одном из генов, необходимых для работы этого механизма), вся эта накопленная скрытая изменчивость может внезапно «выйти наружу», и тогда в популяции произойдет всплеск видимой (явной) изменчивости (см.: Белок Hsp90 контролирует активность мобильных генетических элементов, «Элементы», 19.01.2010).
Предполагается, что скрытая изменчивость может играть важную роль в эволюции, помогая популяциям приспосабливаться к новым условиям. Некоторые мутации, бывшие нейтральными в прежних условиях, могут оказаться полезными при изменении среды. Чем больше таких мутаций накопилось в генофонде популяции, тем больше шансов, что хоть какие-то из них «пригодятся» в новых условиях.
Идея о положительном влиянии скрытой изменчивости на скорость приспособления к новым условиям выглядит правдоподобной и даже самоочевидной. У нее есть немало косвенных подтдверждений, включая результаты математического моделирования (Masel, 2006), но напрямую в строгом эволюционном эксперименте ее пока не проверяли.
Это удалось сделать швейцарским биохимикам, которые выбрали в качестве подопытного объекта не настоящие живые организмы, а квазиживую систему — популяцию размножающихся рибозимов (молекул РНК с каталитическими свойствами).
Авторы использовали хорошо изученный рибозим Azo, изготовленный из самовырезающегося интрона группы I (Group I catalytic intron) бактерии Azoarcus. Этот интрон-рибозим находится в гене изолейциновой тРНК-бактерии. Его функция в бактериальной клетке состоит в том, чтобы самостоятельно вырезать самого себя из молекулы тРНК, то есть осуществить само-сплайсинг (self-splicing). In vitro, то есть в пробирке, этот рибозим умеет осуществлять «обратный сплайсинг», то есть разрезать в определенном месте «субстрат» — короткую молекулу РНК с определенной последовательностью нуклеотидов, причем «обрезки» остаются прикрепленными к рибозиму. По этим прикрепленным обрезкам можно отличить рибозим, успешно выполнивший свою функцию, от рибозима-неудачника. Это позволяет осуществлять искусственный отбор рибозимов, благополучно разрезавших молекулу субстрата.
Авторы размножили исходный рибозим и создали две подопытные популяции (A и B). Эти популяции «жили» и размножались в течение 10 поколений. В каждом поколении проводился отбор на способность разрезать субстрат (короткую молекулу РНК). Те рибозимы, которым это удалось, отбирались и размножались при помощи мутагенной полимеразной цепной реакции (см. также Mutagenic PCR, PDF, 512 Кб). Скорость мутирования составляла в среднем одну мутацию на «особь» на поколение. Чтобы следить за эволюционными изменениями, авторы секвенировали по 2–3 тысячи «особей» (индивидуальных молекул рибозима) из каждого поколения.
Исследователи ожидали, что за 10 поколений такой жизни популяции приобретут значительный запас скрытой изменчивости. Рибозимы будут мутировать, вредные мутации отсеются отбором, а нейтральные накопятся. Известно, что исходный рибозим Azo справляется со своей функцией в очень широком спектре условий (например, он выдерживает нагревание до 80°C). Это говорит о высокой «помехоустойчивости» рибозима и позволяет надеяться, что к мутациям он окажется так же толерантен, как и к скачкам температуры.
Надежды оправдались: обе популяции за 10 поколений накопили изрядное количество мутаций. Исходных, немутантных, молекул «дикого типа» почти не осталось. Большинство особей отличались от исходного рибозима тремя-шестью мутациями.
Приспособленность популяций A и B (то есть их усредненная каталитическая активность) за 10 поколений практически не изменилась. Они не стали справляться со своей функцией лучше, чем исходный рибозим Azo. По-видимому, Azo уже был максимально оптимизирован для выполнения этой функции, и поэтому за 10 поколений эволюционирующим популяциям так и не удалось «найти» такую мутацию, которая бы сделала разрезание субстрата еще более эффективным. Ни одна из возникших мутаций не оказалась полезной и не была поддержана движущим отбором. Таким образом, в течение 10 поколений действовал только очищающий отбор, который отбраковывал «вредные» мутации, а нейтральные игнорировал, пассивно позволяя им накапливаться.
Теперь можно было проверить самое главное: поможет ли накопленная скрытая изменчивость приспособиться к новым условиям. «Новые условия» состояли в том, что исследователи заменили субстрат. Вместо стандартного олигонуклеотида они предложили рибозимам его производное, в котором один из остатков фосфорной кислоты (расположенный как раз в том месте, где рибозим должен разрезать цепочку РНК) был заменен на фосфоротиоат. «Дикий» рибозим Azo справляется с таким субстратом, но с очень большим трудом (с низкой эффективностью).
После смены субстрата эволюционный эксперимент продолжался еще 8 поколений, но теперь в нём участвовали уже не две, а три популяции. К прошедшим 10 поколений искусственной эволюции линиям A и B добавилась третья, контрольная линия, состоящая из рибозимов «дикого типа» (WT) и не имеющая запаса скрытой изменчивости.
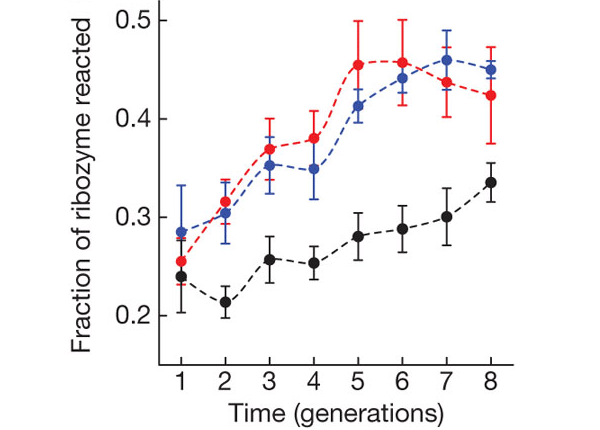
Все три популяции за 8 поколений в той или иной степени «приспособились» к новому субстрату, то есть стали разрезать его эффективнее, чем вначале. Однако популяции A и B приспосабливались быстрее и достигли в итоге большей эффективности, чем линия WT (см. рисунок).
Секвенируя рибозимы последнего поколения, авторы обнаружили, что в популяции A наибольшей численности достиг рибозим-мутант с семью точечными мутациями, получивший название AzoΔ. В популяции B победителем эволюционного соревнования оказался другой мутант — Azo*, у которого обнаружились четыре точечные мутации. Все мутации у двух «чемпионов» оказались разными.
Рибозим Azo*, как показали дальнейшие экспеперименты, действительно разрезает новый субстрат с очень высокой эффективностью. Что же касается рибозима AzoΔ, то в чистом виде он, к удивлению ученых, вообще не смог справиться с новым субстратом. Оказалось, что это своеобразный рибозим-«прихлебатель»: он прекрасно режет новый субстрат только в присутствии других рибозимов (например, исходного или Azo*). Вероятно, другие рибозимы каким-то образом помогают ему свернуться в правильную трехмерную конфигурацию. Подобные ситуации ранее уже были зарегистрированы в экспериментах с рибозимами.
Что касается линии WT, то в ней за 8 поколений так и не появились высокоэффективные мутанты, сравнимые с Azo* или AzoΔ.
Авторы также выяснили, каким образом накопленная скрытая изменчивость помогла популяции B произвести удачливого мутанта Azo*. Оказалось, что после 10 поколений очищающего отбора в популяции уже были рибозимы с одной, двумя и даже тремя мутациями из четырех, составляющих генотип Azo*. Эти мутации по отдельности не дают никакого преимущества ни в старых, ни в новых условиях. Для эффективного разрезания нового субстрата нужны все четыре мутации вместе. Понятно, что наличие у некоторых рибозимов в популяции B отдельных мутаций из этого комплекса резко повысило вероятность возникновения удачного генотипа Azo*.
Исследование наглядно продемонстрировало потенциальную полезность скрытой изменчивости для быстрой адаптации к новым условиям. Чтобы скрытая изменчивость могла накопиться, эволюционирующие организмы должны быть помехоустойчивыми. Если бы исходный рибозим не обладал этим свойством, подавляющее большинство мутаций были бы для него в нормальных условиях вредными. Очищающий отбор безжалостно бы их отсеивал, и при смене условий в популяции не оказалось бы особей, «преадаптированных» к новому субстрату.
Источник: Eric J. Hayden, Evandro Ferrada, Andreas Wagner. Cryptic genetic variation promotes rapid evolutionary adaptation in an RNA enzyme // Nature. 2011. V. 474. P. 92–95.
См. также:
1) Эволюция под управлением компьютера, «Элементы», 12.04.2008.
2) Рибозимы могут размножать друг друга, «Элементы», 13.04.2011.
3) Теплолюбивая бактерия помогает понять происхождение интронов, «Элементы», 28.06.2010.
-
Является ли "мусорная" часть днк запасом скрытой изменчивости? Ведь при нынешних переменах условий это очень актуально
-
Часть мусорной ДНК является посланием инопланетной цивилизации, посеявшей жизнь на Земле путём направленной панспермии. Когда прочтём - сможем вступить с ними в контакт :))
-
С такой скоростью мутаций за несколько поколений послание превратится из "Мой дядя самых честных правил..." в "Мой дядя самых строгих правил..."...
-
(строго) Дык этож ИНОПЛАНЕТЯНЕ! Неужели вы думаете, они неспособны защитить какой-то отрезок ДНК от мутаций, а? :)
Кстати интересно, есть-ли какие-либо ультраконсервативные отрезки ДНК (естественно функциональные) общие для всех живых существ?-
Есть! Гены симметричности и направления развития тела, отвечающие за то где голова, а где зад,где внутренние органы, а где наружние, еще более общие гены это гены белков костей, зубов, ногтей, у любых эукариотических организмов подобные же гены отвечают за образование оболочки клетки, а у бактерий за создание мембраны. В общем рекомендую, почитайте книгу Нила Шубина "Внутренняя рыба"
-
Возможно в самих этих "неприкасаемых" генах закодирован запрет на их мутации - и это ещё одно(и самое главное) доказательство неслучайности мутаций! Но тогда возникает закономерный вопрос - каким же образом в первой "мертвой" молекуле рнк образовался такой совершенный защитный механизм? Можно сформулировать ещё точнее: как появился инстинкт самосохранения у молекулы?
-
1)Возможно? Аккуратнее с предположениями, которые не подтверждены фактами, в информатике есть такое понятие "мусор" на входе, "мусор" на выходе, которое говорит о том что если в рассуждение ввести изначально неверное предположение, правды не получить, а чистой формальной логикой можно доказать все что угодно.
2) По поводу механизма он один и прост как пять копеек, естественный отбор, а точнее происхождение сопровождаемое модификацией, просто гены которые отвечают за план строения тел и белки клеточных стенок и мембран настолько фундаментально важны для выживания, что практически любое изменение в них сразу приводило к смерти индивидума содержавшего этот мутантный ген.
3) "Мертвая" молекула меня вообще позабавила, а молекулы РНК в данном исследовании тоже мертвые? Вы способны дать адекватное определение живому организму? Субстанция которая способна самовоспроизводить себе подобных с модификацией? Согласно книге Ричарда Доккинза Слепой часовщик под это определение подпадают даже мелкие частички глины)))) Почитайте там много ответов.
И ради того чтобы вы понимали как же гены могут "думать", "защищаться" влиять на мир и при чем не только на тот организм который носит эти гены, но и на многие окружающие почитайте книгу хорошую книгу Ричарда Докинза "Эгоистичный ген" и если хватит интереса довольно сложную для неспециалиста книгу "Расширенный фенотип: Длинная рука гена"-
Чтож, по поводу первого пункта согласен, буду осторожнее. Со вторым согласен также. Насчет третьего - слово "мертвая" я поставил в ковычки именно потому что грань провести невозможно. Но вы так и не ответили на второй вопрос. Разве это не просто как пять копеек? Зачем ссылаться на Докинза, когда можно ответить самому своими словами, ведь меня интересует только один вопрос, которому в книге может быть посвящена лишь одна страница. Ради этого читать всю книгу не вижу смысла. Если вам не составляет труда писать длинные ответы, то я прошу вас ответить. Это ведь просто просьба.
Вот есть куча молекул, которые как-то образовали рибозимы(ну тут пока чистая химия), затем они образовали системный динамичный симбиоз с аминокислотами, и вот как эта система стала самовоспроизводится? Может это уже открыто учеными, но я пока не нашел этой информации, в моем понимании здесь пробел. Если у Докинза есть объяснение то может укажете страницу. Был бы признателен-
не составляет труда писать, но сейчас не успею, позже теперь, вопрос попытки объяснить происхождение жизни довольно сложный, одной фразой не отделаешься. Но даже то объяснение которое я могу предложить, заведомо лучше всемогущего творца, так как не оставляет идиотских и при этом очень сложных вопросов типа откуда взялся сам творец.
-
Хорошо, подожду, не тороплюсь никуда.
А насчет творца. Если под творцом понимать некий изначально присущий энергии принцип самоорганизации, то тут все довольно ясно(но вопрос "откуда?" остается). Если же представлять какую-то сверхличность то это не идиотизм, а обычная игра ума - проецировать некий образ вовне. Ум каждого человека играет в такие игры. Многие ученые например неосознанно проецируют свою логику(образ мышления) на устройство вселенной, поэтому слепо верят в создание единой теории(бог ученых)-
Нашел ответ на свой вопрос
http://www.evolbiol.ru/paleobac.htm#1
Но все равно как-то подогнано все, нет четкого представления-
Там в как раз описано практически столько сколько нам известно и в основном то что я бы вкратце Вам описывал. К сожалению палеонтологические раскопки нам дают только общее представление о условиях на Земле 4 миллиарда лет назад. Но Скеп-тик ниже очень верно отозвался и я с ним даже спорить не буду, все в тему и к месту, про глину он не правильно меня понял, но это мелочи, развивать не буду.
В современном океане если б и могла зародиться жизнь, то совсем не в спокойных его частях))) А глубоко под водой там где вулканическая активность, метан, аммиак, взвесь различных металлов, высокая температура(50-80 градусов) и полное отсутствие кислорода. Первый океан был бы для многих современных жизненных форм смертельным местом, там было все то что я описал выше + нестабильная атмосфера: молнии, грозы, ураганы. И где-то читал статью что современные химики в лаборатории попробовали повторить подобные условия и в результате получили бульон различных аминокислот, от которых как вы понимате до РНК - первой самокопирующейся молекулы в общем не очень далеко.
И вот общий смысл что Докинз в своей книге как раз и пытается аппелировать вероятностями сравнивая вероятность того что статуя махнет рукой и тем чтобы такой сложный организм как человек(Или БОГ)))) образовался сразу из сгустка молекул, за один шаг и то и то как вы понимаете практически невероятное событие какой бы вы отрезок времени для проверки ни брали, миллиард лет или 10 со 100 нулями лет не важно, вероятность очень ОЧЕНЬ низка. А вот вероятность того что я скажу разрази меня гром и меня в тот же момент ударит молния В понятиях нашей короткой жизни событие казалось бы такое же маловероятное, а если б я мог прожить миллион лет, насколько я б сильнее опасался такого совпадения? Думаю что для меня это было бы сродни тому что я в метро своего школьного друга встретил с которым лет 10 не виделся, т.е. в общем вероятно, но очень удивительно.
А у Земли был практически МИЛЛИАРД лет прошедших с ее зарождения до появления первых одноклеточных организмов(согласно палеонтологии это определено очень точно) Т.е. даже если считать вероятность того, что рядом окажутся определенные аминокислоты в нужных энергетических состояниях, такой же низкой как и удар молнии в меня после моих слов разрази меня гром, то за миллиард лет это просто должно было случиться и неоднократно.
Но лично я склонен думать что у материи есть такое свойство как стремление к стабильности и усложнению, т.е. существует как бы естественный отбор веществ, стабильные вещества и субстации(галактики, звезды, планеты, молекулы, атомы...) существуют дольше чем нестабильные и в результате идет авторегулирующийся процесс организации материи по пути все увеличивающейся сложности. А потому вероятность зарождения жизни из аминокислот очень высока.
А с вашим творцом я не согласен, давайте называть вещи своими именами во избежание недопонимания. Верим в бога, говорим Творец, не верим, говорим Великие силы природы, а то уж к чему склонны люди это понятно, эволюция у нас в мышелении оставила много ненужных рудиментов))) Но и подарила нам самый мощный разум(на Земле) так давайте им пользоваться, а не слушать чего нам шепчут "устаревшие" участки мозга))). А то таким образом и Энштейна в верующие записали, а он как раз силами природы восхищался))) У Докинза в "Бог как Иллюзия" или по другому "Заблудение богом" об этом написано.
зы. Докинз у меня в последнее время любимый автор так что не судите строго за постоянные апелляции к нему.-
Да, согласен, не трудно представить себе как зародилась жизнь. К тому же в некоторых участках земли молния может ударять по несколько раз в секунду. Читал недавно в новостях что в какой-то стране(забыл блин название) за день от молнии погибло 60 человек. Наверняка несколько человек успели сказать "разрази меня гром" перед смертью))) Природа создала такое разнообразие условий что в них обязательно должна была зародится жизнь и не раз. Но мне бы хотелось увидеть этот процесс если это возможно.
А насчет бога. Я имел в виду не силы природы(во множественном числе), а силу природы, просто изначальную силу или энергию, как угодно. Ведь какую теорию возникновения вселенной не возьми во всех сначала было что-то одно, а затем из него появилось всё многообразие и появляется до сих пор. Вот это изначальное Одно некоторые ученые называли и называют богом. Верующие называют богом тоже самое, но подсознательно придают Ему черты личности, доброй или мстительной - зависит от личности конкретного верующего. Ученые также подсознательно придают Ему качество своей логики, поэтому часто приходится пересматривать свою логику(теорию) когда открывают какую-то новую закономерность этой Силы-
Я не спорю с Вашим определением начала начал, Энергия, Великая Сила природы, Творец, и т.д.
Я лишь призываю, если Вы не верите в некую всемогущую и всезнающую личность не называть Энергию или начальную силу природы - Творцом, потому как понятие Творца-сверхличности подразумевает абсолютное всемогущество!-
Чтож, справедливо. Хотя не совсем ясно почему изначальная Энергия не может быть абсолютно всемогущей и абсолютно разумной? Почему это должна быть обязательно сверхличность? Может просто хочется чтоб было второе, а не первое?
Ладно, это уже разговор не по теме. Если хотите можем продолжить его в другом месте или закончить на этом
-
-
-
-
-
-
-
А я давно сформулировал, что такое жизнь: Самовоспроизводящаяся структура, использующая энергетическую нелинейность среды. Глина под это определение не попадает, но кристаллы в термалных источниках, турбулентные потоки газов в атмосфере, магнитно-плазменые процессы меж термоядерной и конвективной зонами звезд - пожалуста. Только разнообразие этих систем невелико, и "эволюция" далеко не заходит. А "химическая" биология из пятидесяти элементов, плюс потрясающая эквилибристика соединений углерода с шалосями гидридных связей, да плюс низкая энергетика процессов - десятки эВ - живи и радуйся (эволюционируй). По этой причине я не верю ("вера"- мое нелюбимое состояние), что жизнь возникла в океане - слишком стабильные условия. А вот в почвенной влаге, в луже при вулкане - ультрафиолет, оксиды, газообмен - любая ЦеОдва замечтает стать фенолом. Ну а дальше - эволюция...
-
-
-
-
-
-
-
-


Я не специалист и если кому-то не влом - объясните пожалуйста. Вопрос такой:
"Мусорная" же считается часть ДНК которая не экспрессируется?
И если это так, то как же накопленные в ней изменения проявили фенотипический эффект, она вдруг при изменении окружающих условий стала экспрессироваться? Такое действительно происходит что не экспрессирующаяся часть ДНК начинает вдруг считываться?
Прошу прощения если вопрос тупой)
-
В статье говорится не о "мусорной части ДНК". В эксперименте эволюционировала самая что ни на есть экспрессирующаяся, не мусорная нуклеотидная последовательность, кодирующая функциональную молекулу - рибозим.
-
Благодарю за пояснение, значит всё верно сначала понял, комментаторы запутали.
Но вот уж раз натолкнули они на такой вопрос, стало интересно: всё-таки имеют место случаи когда исходно не экспрессирующийся участок ДНК таки начинал экспрессироваться?-
Это возможно, но дело даже не в этом. Многие участки не экспрессируются, но выполняют регуляторные функции, т.е. это не "мусор". Ситуация такая. В геноме (млекопитающих, например) огромное количество "эгоистических" мобильных генетических элементов (напр., ретротранспозонов), которые плодят свои копии в геноме. Они это делают без всякой мысли о нашей пользе. Но, однако, очередная копия такого элемента с ненулевой вероятностью может встать так, что повлияет на экспрессию близлежащего гена, и это может оказаться полезным (или вредным, но тогда отбор отсеет это изменение). Или во встроившейся ранее копии произойдет случайная мутация, которая окажется полезной, и отбор ее поддержит. Поэтому часть "мусора" со временем закономерно и неизбежно превращается в полезные участки генома. Тем более, МГЭ к этому неплохо преадаптированы, ведь у них есть собственные промоторы. См. также: http://elementy.ru/news/430636
-
P.S. Кстати, на мой взгляд разговор о мусорной части ДНК зашел не зря. Ведь представляется достаточно естественным предположить что молекула имеет ограниченный объем багажа нейтральных мутаций. Его превышение становится безусловно вредным, что приводит к тому, часть нейтральных мутаций начинают проявлять движущий характер, в результате чего или исчезают из популяции, или переходят в разряд нужных.
Так вот, исходя из такого предположения. что заставляет думать, что у дикой линии не было багажа нейтральных мутаций, а в двух лабораторных линиях они вдруг накопились?
Скорее всего из-за того, что проверялось всего 3 линии, случайно оказалось что лабораторные приспособились быстрее и лучше. Но все могло быть и наоборот. Для корректного ответа нужны сотни лабораторных линий, и сотни диких.
----------------------------------------------------------
Далее в диаграмме, показывающей степень приспособленности видно что «дикий тип» WT изначально не намного уступает А (0,24 и 0,265 где-то) и В (0,24 и 0,29). В чем же новые условия тогда?
-
Я впал в заблуждение, на самом деле в статье подразумевалось так: раз уже дикий рибозим справляестя с таким новым субстратом с трудом, а
-------------------------------------
«Приспособленность популяций A и B (то есть их усредненная каталитическая активность) за 10 поколений практически не изменилась. Они не стали справляться со своей функцией лучше, чем исходный рибозим Azo.»
--------------------------------------------------
То и А и В должны справляться с субстратом с трудом.
На графике видно, что стартуют А и В уже с форой, то есть A и B все-таки накопили положительные мутации. Причем фора между В и WT равна изначально уже 40% различия приспособленности в итоге. Еще стоит отметить, что практически все итоговое различие между A и WT и значительная доля отличия между B и WT сложилось из-за падения только лишь второго поколения WT и это падение второго поколения из 8-ми поколений всех трех рибозимов было самое значительное и самое выбивающиеся, внесшее значительный вклад в итоговые различия совместно с различными стартовыми условиями.
И почему только 8 поколений, а если больше ? Видно что WT c 5 по 8 поколение превосходит A и В по росту приспособленности.
Последние новости