Обнаружена корреляция между изменениями в составе морских осадков и вымиранием морских животных

Морские животные, населявшие океаны в течение последних 490 млн лет, подразделяются на две большие группы, называемые «эволюционными фаунами», каждая из которых имеет свой характерный темп обновления родового состава. Как выяснилось, скорость вымирания представителей этих фаун коррелирует с распространенностью двух основных типов морских отложений. Это свидетельствует о тесной связи между эволюцией морских животных и характером осадконакопления, которое, в свою очередь, определяется колебаниями уровня моря и другими глобальными геологическими процессами.
Создатель первой глобальной палеонтологической базы данных американский палеонтолог Джек Сепкоски (Jack Sepkoski, 1948–1999) еще в конце 1970-х годов обнаружил, что основные классы животных, населявших мировой океан в течение фанерозойского эона (последние 542 млн лет), подразделяются на три довольно четкие группы, которые он назвал «эволюционными фаунами» (ЭФ). Эти группы различаются, во-первых, характерными темпами обновления таксономического состава (на уровне семейств и родов), во-вторых, временем, когда они достигали максимального разнообразия и доминировали в морских сообществах, в-третьих, некоторыми экологическими особенностями.
Представители всех трех ЭФ живут и поныне, но исторически первой добилась доминирующего положения та ЭФ, которую Сепкоски назвал «кембрийской». Она преобладала в кембрийском периоде. Самые характерные представители кембрийской ЭФ — трилобиты. Две другие ЭФ тогда уже существовали, но занимали подчиненное положение. Для кембрийской ЭФ характерен самый высокий темп обновления таксономического состава (это значит, что роды и семейства в пределах этой ЭФ имеют низкую среднюю продолжительность существования). С ордовикского по пермский период доминировала «палеозойская» эволюционная фауна, для которой характерны более долгоживущие роды и семейства. К палеозойской ЭФ относятся кораллы, брахиоподы, морские звезды, морские лилии и ряд других групп. Большинство представителей этой ЭФ являются неподвижными фильтраторами.
После массового вымирания на рубеже перми и триаса (251 млн лет назад) в морских сообществах стали преобладать представители третьей эволюционной фауны, которую Сепкоски назвал «современной» (двустворчатые и брюхоногие моллюски, морские ежи, ракообразные и др.). Современная ЭФ характеризуется самым медленным темпом обновления таксономического состава. В ней преобладают подвижные животные, которые либо роются в грунте, либо активно передвигаются по его поверхности.
Теория трех эволюционных фаун остается весьма популярной и в наши дни. Она довольно успешно «справляется» с быстрым притоком новых данных и более или менее благополучно выдерживает замысловатые статистические проверки. При этом, однако, многое остается неясным. Например, непонятно, чем обусловлены различия в темпах вымирания и продолжительности жизни родов и семейств в трех ЭФ и почему они сменяли друг друга именно в такой последовательности. Конечно, было предложено множество гипотез, но к единому мнению эксперты пока не пришли.
Шенан Питерс (Shanan E. Peters) из Висконсинского университета в Мэдисоне (University of Wisconsin-Madison) в своей новой статье, опубликованной на сайте журнала Nature, сопоставил эволюционную динамику палеозойской и современной эволюционных фаун с изменениями в составе шельфовых донных отложений. Эти отложения делятся на две основные группы. Терригенные осадки образуются из песка, глины и других материалов, сносимых в океан с материков. В них преобладает кремнезём (SiO2). Карбонатные осадки состоят преимущественно из карбоната кальция (CaCO3). В их образовании принимают активное участие морские организмы, прежде всего планктонные водоросли.
Участки шельфа с преобладанием того или иного типа осадков предоставляют донным животным разные условия жизни. Терригенные осадки обычно мягкие и рыхлые, в них удобно закапываться. Карбонатные осадки более твердые, в них труднее зарываться, зато есть к чему прикрепиться неподвижным животным. Поэтому вполне естественно было предположить, что палеозойская ЭФ, в которой много прикрепленных животных, тяготеет к участкам дна с карбонатными отложениями, а современная ЭФ, в которой много роющих форм, предпочитает терригенные осадки.
Это предположение косвенно подтверждалось тем давно известным обстоятельством, что в палеозое на материковых шельфах в целом преобладало карбонатное осадконакопление, а в мезозое и кайнозое резко возросла роль терригенных осадков. Таким образом, глобальная смена режима осадконакопления примерно совпадает во времени со сменой ЭФ (то и другое произошло примерно на рубеже палеозоя и мезозоя, около 251 млн лет назад).
Однако это могло быть и случайным совпадением. Чтобы доказать реальную связь между развитием ЭФ и режимами осадконакопления, было необходимо сопоставить детальные данные по изменениям характера осадконакопления с динамикой разнообразия представителей палеозойской и современной ЭФ в течение всего периода их существования. Именно это и сделал Питерс.
Прежде всего нужно было выяснить, действительно ли ископаемые представители палеозойской ЭФ чаще попадаются в карбонатных осадочных породах, а представители современной ЭФ — в терригенных. Питерс воспользовался сведениями из международной палеонтологической базы данных (The Paleobiology Database), которая пополняется совместными усилиями ученых из 15 стран и к настоящему времени содержит подробные сведения о 81 144 палеонтологических коллекциях со всего мира. Кстати, вся информация в этой базе является открытой — можно скачать, найти какую-нибудь корреляцию и написать статью в Nature.
Питерс убедился, что представители палеозойской ЭФ действительно чаще встречаются в карбонатных отложениях (59% от общего числа находок), а современной — в терригенных (61%). Само по себе это мало о чем говорит, но, по крайней мере, показывает, что какая-то связь между характером осадконакопления и ЭФ может существовать.
Затем Питерс собрал детальные сведения о 3940 разновозрастных терригенных и карбонатных осадочных толщах, распространенных на территории США. Каждая толща ограничена сверху и снизу так называемыми «перерывами в осадконакоплении». Таким образом, по этим данным можно составить представление о том, когда началось и когда закончилось осадконакопление одного из двух типов в том или ином районе. Кроме того, появилась возможность детально проследить, как менялась со временем распространенность двух типов отложений.
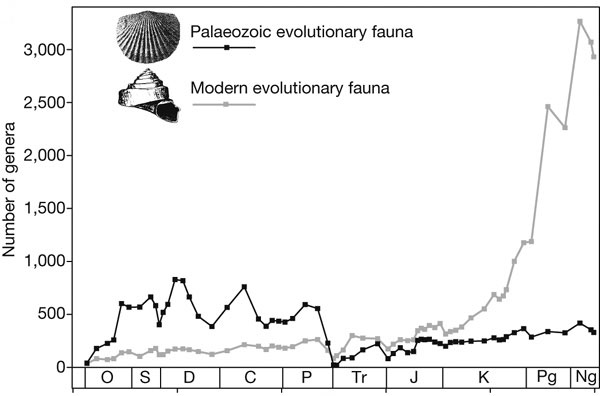
Графики, отражающие динамику двух ЭФ и двух типов осадконакопления, оказались сходными, но только в самых общих чертах (рис. 1–2).

Более интересные результаты дало применение количественного показателя, который можно условно назвать «скоростью вымирания осадочных толщ». Питерс вычислял этот показатель примерно так же, как вычисляют скорость вымирания родов животных. В случае с животными количество родов, которые закончили свое существование в данном временном интервале, делят на общее количество родов, известных из этого интервала, и на длительность интервала. Питерс проделал то же самое с осадочными толщами (возраст нижней границы толщи он рассматривал как время ее «появления», а возраст верхней границы — как время «вымирания»).
Оказалось, что карбонатные толщи «вымирают» в среднем чаще, а «живут» меньше, чем терригенные. Таким образом, динамика карбонатного осадконакопления похожа на динамику палеозойской ЭФ, а динамика терригенного осадконакопления напоминает динамику современной ЭФ.
Самое интересное, что Питерс обнаружил довольно сильную положительную корреляцию между: (1) темпами вымирания родов палеозойской ЭФ и темпами «вымирания» карбонатных толщ и (2) темпами вымирания родов современной ЭФ и терригенных толщ. При этом никакой корреляции между палеозойской ЭФ и терригенными толщами не обнаружилось. Нет корреляции и между современной ЭФ и карбонатными толщами (рис. 3).

Полученные результаты в целом подтверждают идею о том, что динамика ЭФ была обусловлена изменениями характера осадконакопления — распространение карбонатных осадков способствовало развитию палеозойской ЭФ, а терригенные осадки стимулировали развитие современной ЭФ. Но возможны и другие объяснения. Например, исчезновение толщ определенного типа могло приводить не к реальному вымиранию животных, а всего лишь к их исчезновению из палеонтологической летописи. Обнаруженные корреляции могут быть вызваны и тем, что палеонтологи охотнее относят свои находки к разным родам, если толщи, из которых они происходят, разделены перерывом в осадконакоплении. Выбор наиболее правдоподобного из возможных объяснений представляет собой крайне сложную задачу. Для ее решения требуется привлечение дополнительных данных и всевозможные статистические ухищрения. Некоторое количество того и другого Питерс приводит в своей статье. Его окончательный вывод состоит в том, что доводы в пользу первого объяснения (о реальной связи между характером осадконакопления и эволюционной динамикой) представляются наиболее весомыми.
Если Питерс прав, получается, что основные тенденции в эволюционном развитии морской фауны определялись изменениями характера осадконакопления, которые, в свою очередь, были обусловлены колебаниями уровня моря, подъемом и опусканием материков, изменениями климата и другими абиотическими факторами.
Вопросов, конечно, остается очень много. Например, что делать с самой первой, кембрийской ЭФ?
Забавно, что в развитии наземной биоты, как и в море, тоже можно выделить три этапа, характеризующиеся разными наборами доминирующих групп животных и растений: (1) споровые растения и амфибии, (2) голосеменные и рептилии, (3) покрытосеменные, млекопитающие и птицы. Это уже осадконакоплением не объяснишь.
Источник: Shanan E. Peters. Environmental determinants of extinction selectivity in the fossil record // Nature. Advance online publication 15 June 2008 (doi: 10.1038/nature07032).
См. также:
Количественные закономерности эволюции.
-
А рассматривает ли Питерс также такую проблему, как изменение соотношения мелководных и глубоководных отложений в связи с трансгрессиями и регрессиями, приводящее к "псевдовымираниям"? Пример такого подхода см., напр, здесь: Smith A.B., Gale A.S., Monks N.E.A. Sea-level change and rock-record bias in the Cretaceous: a problem for extinction and biodiversity studies // Paleobiology, 27(2), 2001, pp. 241-253 (pdf: http://homepage.mac.com/nmonks/files/papers/smith_et_al.pdf)
-
Нет, это он обходит. Как и многое другое (сейчас так много развелось публикаций о всяких реальных и воображаемых ошибках в летописи, что все исследователи только тем и озабочены, как бы так извернуться и ускользнуть от всех этих трудноразрешимых проблем, иначе вообще невозможно никакие количественные тренды в летописи анализировать).
-


Последние новости