Замена небольшого участка генома на синонимичный заставляет бактерию иначе решать ту же эволюционную задачу
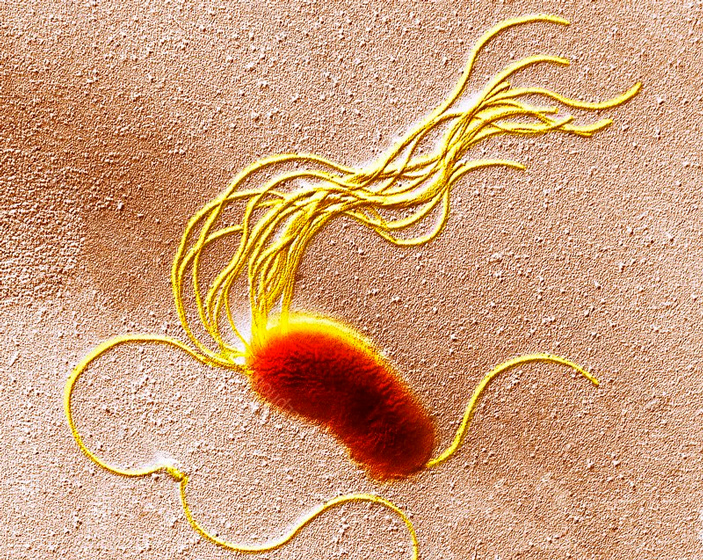
Биологическая эволюция базируется на двух основных процессах: мутационной изменчивости и естественном отборе. Мутационная изменчивость выступает в роли поставщика эволюционного материала, производя случайным образом разные мутации, из которых отбор сохраняет лишь полезные. Таким образом, согласно общепринятому представлению, именно отбор выполняет направляющую роль в эволюции и именно от него зависит, какими окажутся генотипы потомков, после того как в ряду поколений организмы адаптировались к какому-то новому неблагоприятному фактору среды. Однако ученые из Великобритании, работая с двумя близкими, но немного разными штаммами бактерии Pseudomonas fluorescens с искусственно поломанной системой формирования жгутиков, показали, что сам процесс мутагенеза может иметь весьма закономерный характер и оказывается порой не менее значимым в предопределении конечного генофонда эволюционирующей и адаптирующейся популяции, чем фактор отбора. Спектры новообретенных мутаций, позволивших бактериям заново получить рабочий жгутик, оказались для разных штаммов разными, но хорошо воспроизводились в повторных экспериментах. А перестановка небольшого фрагмента ДНК между штаммами заставляла их переключаться с одного эволюционного пути на другой.
В возглавляемой Тиффани Тейлор (Tiffany B. Taylor) лаборатории, относящейся к университету города Бат (графство Сомерсет, Англия), уже давно работают с простым, маленьким, неприхотливым и при этом очень интересным модельным объектом — бактерией Pseudomonas fluorescens.
Pseudomonas fluorescens — палочковидные грамотрицательные бактерии. Эти микроорганизмы обладают несколькими средствами для активного перемещения по поверхностям твердых субстратов. Во-первых, они могут небыстро передвигаться, «расползаясь» по поверхности твердого субстрата (таковым может быть почва в дикой природе, или агар в лабораторной чашке Петри), выделяя биосурфактант вискозин — небольшой биогенный пептид, который не следует путать с одноименным искусственным сурфактантом (см. M. V. Laycock et al., 1991. Viscosin, a potent peptidolipid biosurfactant and phytopathogenic mediator produced by a pectolytic strain of Pseudomonas fluorescens). Во-вторых, эти бактерии могут плавать при помощи жгутиков (при условии, что твердый субстрат покрыт хотя бы очень тонким слоем жидкости). Второй способ передвижения быстрее.
В лаборатории можно получить мутантные линии бактерий P. fluorescens, полностью лишенные возможности активного передвижения по субстрату. Эти мутанты прекрасно себя чувствуют, когда их выращивают в жидком питательном бульоне с активным перемешиванием, но вот на плотной среде с агаром они быстро выедают питательные вещества в зоне роста колонии, а потом начинают голодать и... думаете, вымирают? Как бы не так! Нет, они эволюционируют!
Фокус внимания Тейлор и ее коллег направлен на выяснение того, как именно осуществляется адаптация у бактерий, какова роль мутаций и отбора в этой адаптации, есть ли закономерности в процессе адаптивной эволюции и чем они обусловлены.
У бактерий имеются гены, которые кодируют структурные белки, непосредственно формирующие нить жгутика; еще ряд генов кодирует белки мотора, который, собственно, приводит жгутик в движение. Экспрессия всех этих генов, собранных в общий оперон, регулируется особым белком FleQ (S. K. Arora et al., 1997. A transcriptional activator, FleQ, regulates mucin adhesion and flagellar gene expression in Pseudomonas aeruginosa in a cascade manner). Этот белок, выполняющий функцию фактора транскрипции, взаимодействует, с одной стороны, с определенными последовательностями ДНК вблизи точки начала транскрипции, а с другой стороны — с одной из субъединиц РНК-полимеразы, которая, собственно, обеспечивает транскрипцию. Клетки, у которых ген fleQ не работает, не могут построить ни жгутик, ни мотор к нему, даже несмотря на то, что гены самих строительных белков для этих структур остаются в целости и сохранности.
В 2014 году при участии Тиффани Тейлор был выведен штамм P. fluorescens, получивший имя SBW25 (A. L. Alsohim et al., 2014. The biosurfactant viscosin produced by Pseudomonas fluorescens SBW25 aids spreading motility and plant growth promotion). Ученые выделили его из дикой природы, а потом модифицировали в лаборатории, внеся поломку (делецию) в ген fleQ. Так они получили бактерий, лишенных жгутиков. Этот штамм все же сохранял способность перемещаться по твердому субстрату посредством расползания. Дополнительная поломка в гене viscB (он кодирует фермент для синтеза вышеупомянутого биосурфактанта) лишила бактерий и этого способа перемещения. Полученный штамм-инвалид, полностью неспособный к активному перемещению, был обозначен AR2.
В 2015 году также при участии Тиффани Тейлор из природного изолята был получен еще один штамм этого же вида бактерий, обозначенный Pf0-1. Этот штамм оказался изначально лишенным способности выделять биосурфактант и «ползать» по поверхности из-за естественной мутации в регуляторном гене gacA. В геном этого штамма тоже внесли делецию в гене fleQ, обозначив новый штамм Pf0-2x. Так же, как и AR2, эти бактерии потеряли способность строить жгутики и распространяться «вплавь» по поверхности агара.
Экспериментируя со штаммами AR2 и Pf0-2x, ученые столкнулись с совершенно неожиданным феноменом: бактерии, лишенные рабочего гена fleQ и высеянные на чашку с агаром, очень скоро полностью восстанавливали жгутики и полноценную способность к быстрым перемещениям по поверхности среды: на это требовалось не более восьми суток (в среднем — около 4-х), причем результат воспроизводился снова и снова во множестве повторных экспериментов. Эти результаты описаны в статье T. B. Taylor et al., 2015. Evolutionary resurrection of flagellar motility via rewiring of the nitrogen regulation system.
В ходе дальнейшей работы исследователям удалось прояснить цепочку событий адаптивной эволюции, посредством которых бактерии «излечиваются» от своей инвалидности. Эта цепочка, состояла всего из двух поочередно приобретаемых мутаций, причем сама последовательность событий, обеспечивающих адаптацию, тоже воспроизводилась в экспериментах снова и снова (рис. 2).

Рис. 2. Эксперимент по индукции адаптивной эволюции и этапы адаптивной эволюции безжгутиковых бактерий в условиях голодания на чашках с агаром. Рисунок из статьи T. B. Taylor et al., 2015. Evolutionary rewiring of bacterial regulatory networks
На первом этапе адаптивной эволюции фиксировалось появление мутации в одном из трех генов (glnA, glnK или ntrB), после которой у бактерий появлялась ограниченная способность к перемещениям на плотной среде. На втором этапе добавлялась мутация в гене ntrC, и бактерии восстанавливали способность к быстрому эффективному перемещению, сопоставимую с исходными природными штаммами. Это плавание обеспечивалось благодаря реверсии полноценных рабочих жгутиков.
Здесь нужно сказать немного о функциях белков, кодируемых каждым из вышеупомянутых генов.
Белок NtrC (так же, как и FleQ) относится к группе активаторов транскрипции. Однако он узнает иные последовательности в ДНК и управляет другими наборами генов. В его подчинении находятся гены, участвующие в поглощении и метаболизме азота. Азот нужен живым клеткам для синтеза аминокислот, но вместе с тем избыток соединений азота во внутренней среде токсичен, так что все эти процессы должны быть хорошо сбалансированы.
Белок NtrB — фермент, который осуществляет фосфорилирование белка NtrC, необходимое для его активации (то есть именно в фосфорилированном состоянии он начинает работать как усилитель транскрипции генов). Активность NtrB в свою очередь зависит от других белков, чувствительных к внутриклеточной концентрации азота и его соединений, в том числе некоторых аминокислот (таких, как, к примеру, глутамин).
GlnA — это фермент синтеза глутамина, от его активности зависит количество образуемого в клетке глутамина. GlnK — это регуляторный белок, чувствительный к концентрации азота и регулирующий большую сеть белков регуляции азотистого обмена, в том числе, белки GlnA и NtrB. Повышение активности GlnK ведет к снижению активности NtrB, и наоборот.
Итак, выходит, что восстановление способности к передвижению у безжгутиковых штаммов P. fluorescens так или иначе обеспечивается кооптацией генов системы регуляции азотистого обмена. В конечном итоге синтез и работа жгутиков оказываются восстановленными за счет того, что гены, в норме активируемые белком FleQ, переходят под управление мутирующего в ходе адаптивной эволюции белка NtrC.
Любопытно, что среди множества белков этой бактерии NtrC не самый близкий по структуре белок-активатор к FleQ (идентичность аминокислотного состава этих двух белков около 30%, имеется четыре белка с более высоким уровнем сходства). Тем не менее, у всех исследованных мутантов (полученных в параллельных экспериментах) именно он брал на себя функцию заместителя во включении генов, ответственных за работу жгутика. Как предполагают авторы исследования, это может быть непосредственно связано с воздействием среды в условиях эксперимента. Не имея возможности перемещаться, клетки голодают, возникает недостаток азота, что стимулирует фосфорилирование и активацию белка NtrC. То есть именно этот белок физиологически оказывается в нужном состоянии, чтобы взять на себя роль заместителя недостающего активатора FleQ.
Мутации первого этапа эволюции прямо или опосредованно усиливают активность белка NtrB. Как следствие, благодаря повышению частоты фосфорилирования, усиливается и активность NtrC. Эта избыточная активность одновременно усиливает как синтез белков, отвечающих за поглощение азота, так и, хоть и слабо, но активирует работу генов, ответственных за синтез и работу жгутиков. Мутация в самом NtrC, возникающая на втором этапе адаптивной эволюции бактерий, меняет его специфичность: повышается его сродство к регуляторным последовательностям генов жгутикового аппарата, и вместе с тем, несколько снижается сродство к регуляторным последовательностям азотистого обмена. В итоге оба эффекта оказываются для бактерий положительными: возвращается способность быстро плавать, а клетки перестают чересчур активно поглощать из среды азот (чрезмерное поглощение азота возникает как побочное следствие мутаций первого этапа адаптивной эволюции).
В исследовании, результаты которого были опубликованы в журнале Nature Communications в октябре 2021 года, авторы, в сущности, еще раз воспроизвели эксперименты 2015 года все с теми же двумя штаммами AR2 и Pf0-2x, но теперь сосредоточились на том, насколько сходны пути адаптации, реализуемые в ходе эволюции у каждого из штаммов.
Первый этап адаптивной эволюции у двух штаммов проходил действительно по-разному. У штамма AR2 мутации фиксировались в гене ntrB, причем в 95% (в 23 независимо сформированных клонах из 24) это происходило в одной и той же нуклеотидной позиции: аденин под номером 289 (считая от первого нуклеотида в рамке считывания) заменялся на цитозин (такая замена обозначается A289C). Эта мутация приводит к замене в белке NtrB аминокислоты треонина в позиции 97 на пролин (T97P). В одном случае из 24 в гене ntrB образовалась не замена, а делеция участка длиной в 12 нуклеотидов (в позициях 406–417).
В противоположность AR2, у бактерий штамма Pf0-2x в тех же самых условиях возникали очень разные мутации. Это набор различных замен, инсерций или делеций как в самом гене ntrB, так и в генах glnK или glnA. Причем среди всех изученных мутантов Pf0-2x не оказалось ни одного (!) с мутацией A289C.
Эффекты всех выявленных у обоих штаммов мутаций оказались достаточно схожими по тому, какое влияние (в количественных показателях) они оказывали на подвижность бактерий и на азотистый обмен.
Можно выдвинуть два варианта объяснения такого высокого параллелизма в возникновении одной и той же мутации у штамма AR2: либо эта мутация оказывается по некоторым причинам особенно полезной именно для бактерий этого штамма (но не штамма Pf0-2x), либо в силу некоторых причин это наиболее часто возникающий вариант мутации у этого штамма (но, опять же, не у Pf0-2x). Так или иначе, причина лежит в каких-то особенностях устройства геномов двух штаммов.
Авторы придумали эксперимент, который бы позволил либо подтвердить, либо опровергнуть вторую гипотезу. Они предположили, что частому возникновению мутации A289C может содействовать структура цепочки ДНК в окрестностях 289 нуклеотида. В его непосредственной окрестности (не далее 14 нуклеотидов в обе стороны) имеется шесть нуклеотидов, различающихся между штаммами AR2 и Pf0-2 (C276G, C279T, C285G, C291G, T294G и G300C; здесь до номера позиции указан нуклеотид из последовательности AR2, а после номера — из Pf0-2x). Все они синонимичны, то есть кодируют одни и те же аминокислоты в белке. И вместе с тем, моделирование потенциальных вторичных структур, которые могут формироваться в участке ДНК с двумя разными вариантами последовательности, дает неодинаковый результат. Если дело действительно в этом, то можно проверить, как поменяется спектр мутаций, если устроить подмену последовательностей в геноме каждого штамма.
Итак, заменив шесть нуклеотидов в геноме штамма AR2 на шесть соответствующих по положению нуклеотидов из генома штамма Pf0-2x, авторы получили новый экспериментальный штамм AR2-sm. Аналогичная процедура по модификации генома Pf0-2x дала штамм Pf0-2x-sm6.
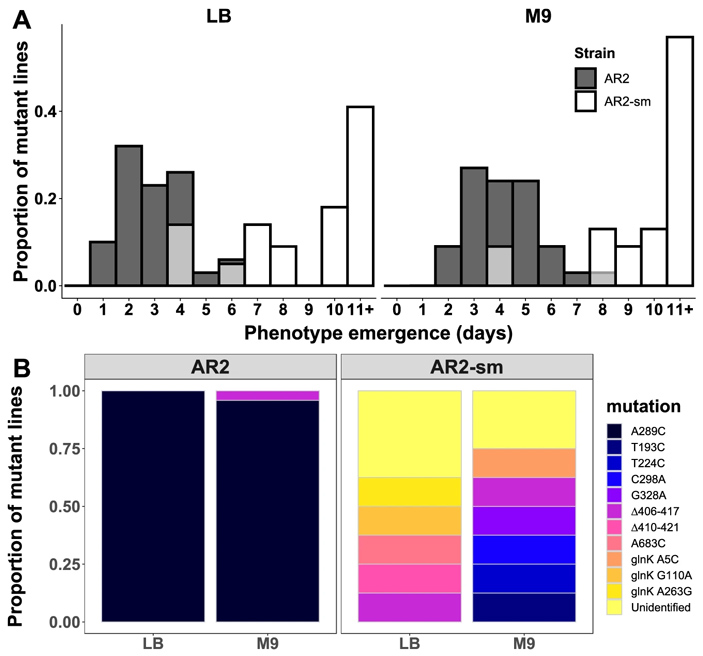
Рис. 3. Адаптация штаммов AR2 и AR2-sm к росту на агаризованной среде (эксперимент проводился с использованием двух вариантов питательной среды: богатой питательными веществами LB, и «диетической» M9). А — скорость адаптации, то есть число дней до появления подвижных клонов. По горизонтальной оси — количество дней до появления мутантных клонов с подвижным фенотипом. По вертикальной оси — их доля среди всех проанализированных клонов. Видно, что два штамма (strain) адаптируются с разной скоростью. B — спектры мутаций, обнаруженных у клонов с восстановленной подвижностью для штаммов AR2 и AR2-sm. Рисунок из обсуждаемой статьи в Nature Communications
Эти штаммы поместили в те же селективные условия, что и родительские штаммы. Посмотрев на рисунки 3 и 4, вы легко заметите, что ситуация резко поменялась на прямо противоположную. Теперь уже штамм, производный от Pf0-2x, давал преимущественно мутантов A289C по гену ntrB (она была у восьми клонов из десяти проанализированных), а у штамма AR2-sm такая мутация не появилась ни разу (на в одном из восьми проанализированных клонов), адаптация же формировалась за счет самых разных мутаций в трех разных генах.
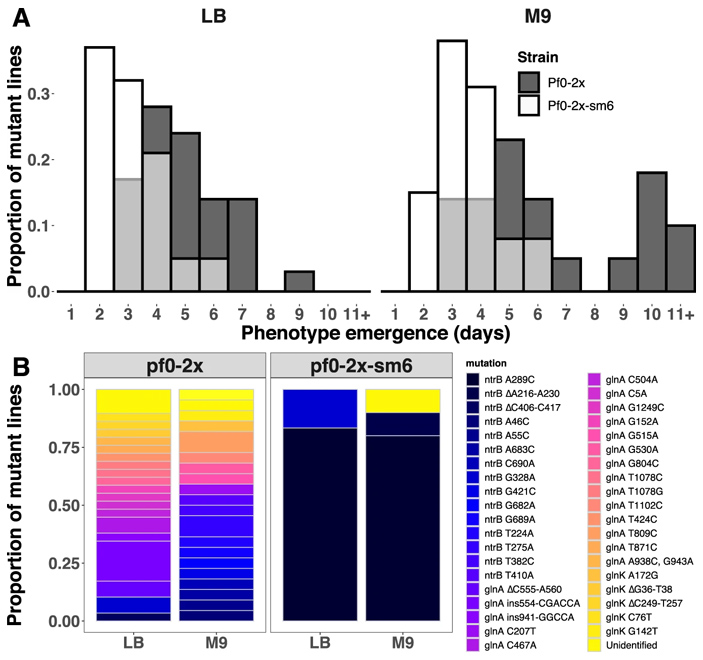
Рис. 4. Адаптация штаммов Pf0-2x и Pf0-2x-sm6 к росту на агаризованной среде. А — скорость адаптации (число дней до появления подвижных клонов). B — спектры мутаций, обнаруженных у клонов с восстановленной подвижностью для штаммов Pf0-2x и Pf0-2x-sm6. Рисунок из обсуждаемой статьи в Nature Communications
Была замечена, к тому же, разница в скорости приобретения адаптивных изменений: штамм AR2-sm немного медленнее восстанавливал подвижность, чем родительский штамм AR2. И наоборот, штамм Pf0-2x-sm6 восстанавливал подвижность немного быстрее по сравнению с родительским штаммом Pf0-2x. То есть структурные особенности небольшого фрагмента ДНК, позволяющие с высокой вероятностью произвести полезную мутацию A289C в гене ntrB, дают любому из двух штаммов преимущество в способности быстро ответить на давление среды адаптивными изменениями. Кстати, аналогичное влияние структуры ДНК на частоту параллелизмов в адаптивной эволюции было также продемонстрировано ранее у колюшек (Ломкие сайты хромосом предопределяют параллельную эволюцию у трехиглой колюшки, «Элементы», 23.01.2019).
В английском языке для описания способности организма быстрее или медленнее формировать новые адаптации используют специальный термин evolvability. В русском языке для этого понятия, к сожалению, пока нет удобного эквивалента. Изучение факторов, благодаря которым у разных видов живых существ разная evolvability, — одно из горячих направлений в современной эволюционной биологии, в котором пока много открытых очень интересных вопросов. В частности, было бы здорово понять, могут ли эволюция и необходимость адаптации к часто меняющимся факторам среды содействовать формированию таких структурных особенностей молекул ДНК, которые повышают эту самую способность быстро производить новые адаптации (evolution of evolvability).
Источник: James S. Horton, Louise M. Flanagan, Robert W. Jackson, Nicholas K. Priest & Tiffany B. Taylor. A mutational hotspot that determines highly repeatable evolution can be built and broken by silent genetic changes // Nature Communications. 2021. DOI: 10.1038/s41467-021-26286-9.
Татьяна Романовская
-
Я в своих эволюционных модельках способ упаковки генома, влияющий на характер возможных мутаций этого генома, тоже подвергал давлению отбора (т.е., в генокоде особи дополнительные гены отвёл для хранения параметров мутаций, применимых для различных частей генома, в том числе для самих этих генов). Называл это регуляцией «эволюционной подвижности», почему-то мне казалось, что это общепринятый термин (щас не смог нагуглить). Возможно, я как раз изобрёл русскую версию термина evolvability :).
О, вот нагуглил, есть общепринятое «эволюционная пластичность», это не оно? Ещё прямо на Элементах нашлось устойчивое словосочетание "направленность мутагенеза" на схожую тематику :).
И, кстати, ответ на последний вопрос статьи - да, системы с эволюционной динамикой способны «помнить» об «истории» своей эволюции и адаптивно «канализировать» мутагенез. Вообще, кажется, подобные вопросы лежат в области логики (математики), а не биологического эксперимента. Ценность адаптации канализирования мутаций на компьютерных моделях можно показать со степенью абстрактности, достаточной для проецирования на любые биологические системы. Реальные биологические механизмы этой «кармической памяти генома», конечно, у различных видов могут быть оч разными и их полезно найти (и найти характер изменения факторов отбора, делающих этот механизм полезным), но самая базовая механика дана абсолютно всем живым существам с неточечной геометрией генома в мире с ненулевой длительностью любой химической реакции (и различающимися длительностями различных реакций).
Чисто математически адаптивная направленность мутагенеза эквивалентна появлению у каждого поколения дополнительного раунда отбора, не требующего реальной генерации поколения потомков (геном как бы заранее предсказывает его результаты, виртуально убивая всех мутантов, не попадающих в «канал мутагенеза»).
И, кстати, половой процесс (и последующее возникновение полового отбора и разделения полов) - это тоже канализирование эволюции, уже «следующего» «комбинаторного» уровня (сохранение в геноме иерархических зависимостей колебаний более комплексных факторов отбора и реакции генома на них), который невозможен (в рамках эволюционно-стабильных стратегий) без предыдущего «геометрического» (сохранения в геноме «инерции» прошлых мутаций, неявно закодированных положением гена на хромосоме/ДНК и неравномерностью профиля химической устойчивости гена к различным видам мутаций).
В своей модельке когда-нибудь расширю «гены параметров возможных мутаций» до каких-нибудь «генов полнотьюринговых мутационно-рекомбинирующих субмашин», чтобы покрыть сразу «все» потенциальные уровни эволюции evolvability, и выращу половой диморфизм в системе, изначально разделения по полам не содержащей :). -
Мутационная изменчивость -это сила, продавливающая жизнь сквозь сопротивление среды. Сила активна, она постоянно напирает, среда же равнодушна, но тоже меняется под воздействием иных сил. Как бы, среда - это лабиринт с движущимися коридорами, а жизнь должна продавиться к выходу. Кто застрянет, тот сам виноват.
И тут вдруг "помогите, хулиганы зрения лишают", то бишь, жгутиков. "Всё пропало, шеф?". Отнюдь, внутри проекта псевдомонады зашит план Б. Очень горячий план отращивания жгутиков другим способом, наводящий на жгучую мысль о наличии тайной сверхадаптационной структуры внутри проекта. Однако, идея высшей структуры тянет за собой идею высших сил.
Жгутики повторно появились просто потому, что их проще всего создать внутри клетки. Вот если бы вместо жгутиков покалеченная монада отрастила весла с парусом, тогда бы ой...-
Вместо высших сил можно заподозрить существование аттрактора. Но да, слабо верится, что какое-то геометрическое положение конкретного гена в геноме или его химическая активность могут стать переключателем эволюции между столь радикально различающихся приспособлений (жгутики и паруса), для этого на ландшафте приспособленности бактерии должна бы была идти очень-очень-очень узкая дорожка приспособленности к абсолютно специфичным условиям :).
-
"Образно говоря, аттрактор выполняет функции автопилота, удерживая сложный комплекс авиационных приборов и средств управления в определенных режимах и параметрах, не позволяющих сбиться с курса". Цитата отсюда https://terme.ru/slovari/istorija-i-filosofija-nauki.html
Получается, что автопилот есть в каждом геноме каждого живого существа прямо от Луки. Ну, как тут не перекреститься? Опять же, никакие аттракторы не помогли динозаврам и всем прочим древним тварям...-
Не, если пользоваться этой метафорой, то эти "квазиаттракторы" evolvability есть у каждого генома, но ведут эти аттракторы по ландшафту приспособленности куда попало. Можно представить evolvability в виде инерции этого движения. И успешные курсы, ведущие геном по ландшафту к процветанию, и ошибочные курсы, ведущие к вымиранию, имеют некоторую инерцию. Сам ландшафт приспособленности виртуален ("невычислим" без физической генерации потомков) и напрямую недоступен для анализа, но у генома есть "историческая память" о своих прошлых мутациях (какое-то состояние генома, делающее одни мутации более вероятными, а другие - менее). Эти отпечатки истории в геноме (в какой бы форме они ни сохранялись) по сути являются информацией об уже "прощупанном" в прошлом ландшафте приспособленности.
Какими механизмами можно хранить в геноме прошлое? Например, хранение в ДНК нерабочих обломков генов даёт материал для мутаций - какой-то ген, успешный 1000 поколений назад, но сейчас сломанный, в результате единичной мутации может включиться обратно, а без этого обломка геному пришлось бы добираться до нужного гена через 1000 мутаций (т.е., в этом случае геном "не бывал" в той области приспособленности, "не помнит" её). Или, например, хромосома своей геометрией делает какие-то участки генома более хрупкими и мутирующими (подверженными радиации или сложными для репарации), и тогда туда "сползают" гены, кодирующие признаки под давлением самых актуальных изменений среды. Т.е., скорость мутаций синхронизируется со скоростью изменения среды, фенотипические признаки вида под увеличивающимся давлением отбора становятся более эволюционно подвижными.
Подозреваю, что биологическая эволюция зашла дальше и реализовала и более сложные и многоуровневые механики управления геномом, подмешивая "неслучайность" (например, какие-нибудь циклические стратегии в духе "этот ген выключить на 100...150 поколений" или условия в духе "этот ген комбинировать только с таким-то геном по такому-то шаблону") в случайные генетические процессы. Да, кривые примеры "геномных программ", навскидку не могу ничего более толкового придумать, и на компе ещё не моделировал подобное. Когда буду заниматься, то, наверное, постараюсь сделать какую-то максимально абстрактную и универсальную машину "любых геномных операций", чтобы не зависеть от ещё не сделанных открытий в биологии (по сути, мои кибер-существа будут обладать технологией прямого редактирования своего генома изначально, им не надо будет его изобретать, надо будет только наращивать стабильные нелетальные алгоритмы этого редактирования :))-
Представим себе эволюционную нишу в форме полого диска: вход на 6 часов, выход на 12, в середине лабиринт. Жизнь течет, и вдруг выход смещается на 3 часа. Понятно, что память не поможет найти новую дорогу.
А что помогло нащупать старую дорогу? У нас есть поток, значит, можно применить гидравлические понятия. Тогда получается, что правильная струя течет быстрее прочих. Следовательно, давление внутри неё меньше, чем у остальных. Ну, и... Манометр - наше всё!
Что внутри клетки подойдет на роль "напряжёметра"? Три нобелевки, и дело в шляпе...)))-
Ваша метафора, действительно, об предсказании принципиально неизвестного будущего ландшафта. А моя метафора - об самомодификации генома на основе данных о прошлой эволюции вида (что для внешнего наблюдателя в масштабе множества поколений популяции будет выглядеть проявлением «знания будущего», но будет являться, по сути, «инерцией знания прошлого» :))
-
Э-э... Жизнь течет по пути наименьшего сопротивления. В какую сторону ручеек течет быстрее, там и жизнь живее. Философия эволюции, понимаете ли...)))
-
Для популяции эволюционная динамика получится сложнее, чем просто «плыть по течению». Это философия многофакторной оптимизации в условиях динамического изменения факторов :).
Допустим, у нас фиксированный и заранее известный (нам, исследователям) ландшафт приспособленности вида - некий лабиринт из гор и холмов с долинами, каньонами и ядовитыми озёрами (на нулевом уровне приспособленности). На этом ландшафте находится рой точек, представляющий популяцию, каждая точка - это индивид со своим генотипом (в том числе и со своим направлением эволюционной инерции). Точки мутируют (с учётом и этих разнонаправленных инерций), суетятся вокруг некоего центра масс, но остаются цельной популяцией, т.к. слишком далеко отлетевшие одинокие точки вымирают из-за генетической несовместимости с основным контингентом вида.
Таким образом рой кучкой медленно двигается по ландшафту под давлением отбора (по наклону ландшафта) или дрейфует в случайном направлении. И вот он доехал до ядовитого озера, крайние точки роя «прикасаются» к нему и начинают хуже выживать, оставляя меньше потомства, чем центровые, и таким образом меняя в популяции соотношение «прикоснувшихся к смерти» версий генотипа и генотипа «центровых». Таким образом рой накапливает информацию и «использует» её - геном начинает двигаться в сторону от ядовитого озера (точек рядом с озером рождается меньше, а с противоположной стороны роя - больше).
К примеру, по курсу обнаружилась новая опасность, но теперь рою нужно не просто от неё отбежать, но ещё и не вернуться к предыдущей опасности, и рой выбирает какое-то третье направление, используя свою «память» о прошлом столкновении.
К примеру, таких толканий и ощупываний озёр уже накопилось много в истории генофонда, и тогда рой может «предположить» даже некую периодичность этих озёр (если они и правда периодичные, и структура генома достаточно развита, чтобы «почувствовать» эту периодичность), и объехать третье озеро уже даже не прикасаясь к нему.
Не знаю, насколько сложным может быть поведение генома по итогу (с какой скоростью в нём наращиваются обратные связи, или, другими словами, насколько сложной может развиться та самая «машина геномных операций»).-
Подставьте в вашу модель множество роев, двигающихся во всех направлениях. Те рои, что наткнутся на эволюционно ядовитые участки, там и умрут, либо мутируют и остановятся. То есть, уткнутся тупик лабиринта. Какие-то рои будут медленно продвигаться по холмам. Ну, а одна эволюционно удачная популяция случайно(!) выберется на тучное место и будет усиленно питаться/размножаться. Здесь образуется новое множество популяций. И если ландшафт вдруг наклонить, именно эта новое множество снова начнет двигаться во всех направлениях. И так без конца, вплоть до той популяции, которая сядет за компы и сама начнет наклонять природу...)))
-
В том и дело, что рой благодаря эволюционной динамике, показанной выше, сможет выбираться из тупиков. На первых порах случайно, но с накоплением опыта будет делать это всё эффективнее (применяя успешный опыт решения старых проблем к новым, по сути). Или в чём состоит ваша критика? (Или ваш комментарий - не критика?) Да, какие-то виды (рои) будут блуждать по лабиринту, какие-то осядут на полянках с благоприятными условиями, каких-то утопят «озёра смерти», и лишь немногие доберутся до условного финиша лабиринта :).
Рой генотипов какого-то животного на ландшафте приспособленности сам ведёт себя как животное, очень медленное. Жизнь отдельной особи животного в реальном мире - это как жизнь одной клеточки такого метаживотного в мире виртуального ландшафта приспособленности. Возможно, при обретении животным мозга или «материально-культурного» способа ускоренной обработки информации появляется следующий метауровень метаживотного со своим металандшафтом метаприспособленности, а при разделении генотипа на несколько полов, например, у метатуловища появляются какие-нибудь метажгутики или метамембраны (поведение точек роя дифференцируется на отдельные классы).
Ох, ладно, меня и так уже унесло в очень умозрительные метаконцепции, лучше пока приторможу в своих выводах, до реализации модельки с хоть немного похожей на вышеописанную эводинамикой. Основная сложность всего этого компьютерного моделирования - в дизайне необходимого градиента экологических проблем, ведущих к описанному усложнению структуры генома, нужно построить самоусложняющийся мир (или мир связанных экологических ниш с различными сложностями). К примеру, если мы хотим научить геном играть в шахматы, нам нужно будет начинать с какой-то экологической ниши для тренировки хождения пешками на поле 3*3, например, последовательно её усложняя до полных шахматных правил :).
-
-
-
-
-
-
-
-
-


Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
-
 24.08.2023Бактерия с синтетическим минимальным геномом сохранила способность к адаптивной эволюцииАлександр Марков • Новости науки
24.08.2023Бактерия с синтетическим минимальным геномом сохранила способность к адаптивной эволюцииАлександр Марков • Новости науки
-
 12.06.2023В эволюционном эксперименте дрожжи превратились в макроскопические многоклеточные организмыАлександр Марков • Новости науки
12.06.2023В эволюционном эксперименте дрожжи превратились в макроскопические многоклеточные организмыАлександр Марков • Новости науки
-
 15.08.2022Эволюция генетического разнообразия бактерий направляется внутривидовой конкуренцией и гонкой вооружений с фагамиАлександр Марков • Новости науки
15.08.2022Эволюция генетического разнообразия бактерий направляется внутривидовой конкуренцией и гонкой вооружений с фагамиАлександр Марков • Новости науки
-
 14.01.2022Замена небольшого участка генома на синонимичный заставляет бактерию иначе решать ту же эволюционную задачуТатьяна Романовская • Новости науки
14.01.2022Замена небольшого участка генома на синонимичный заставляет бактерию иначе решать ту же эволюционную задачуТатьяна Романовская • Новости науки
-
 14.01.2021В ходе эволюционного эксперимента удалось получить генералистов при симпатрическом видообразованииЕлена Наймарк • Новости науки
14.01.2021В ходе эволюционного эксперимента удалось получить генералистов при симпатрическом видообразованииЕлена Наймарк • Новости науки
-
 18.11.2019Новый метод генетического штрихкодирования позволяет детально следить за эволюцией дрожжейАлександр Марков • Новости науки
18.11.2019Новый метод генетического штрихкодирования позволяет детально следить за эволюцией дрожжейАлександр Марков • Новости науки
-
 11.11.2019Симбиотические дрожжи помогают дрозофилам адаптироваться к соленому кормуАлександр Марков • Новости науки
11.11.2019Симбиотические дрожжи помогают дрозофилам адаптироваться к соленому кормуАлександр Марков • Новости науки
-
 15.08.2019В ходе эволюционного эксперимента у дрожжей появился новый генТатьяна Романовская • Новости науки
15.08.2019В ходе эволюционного эксперимента у дрожжей появился новый генТатьяна Романовская • Новости науки
-
 06.11.2018В ходе эволюционного эксперимента патогенный гриб превратился в полезного симбионтаАлександр Марков • Новости науки
06.11.2018В ходе эволюционного эксперимента патогенный гриб превратился в полезного симбионтаАлександр Марков • Новости науки
Последние новости





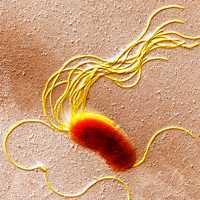
Рис. 1. Бактерия Pseudomonas fluorescens (трансмиссионная электронная микрофотография). Хорошо видны ее многочисленные жгутики (желтые). Ширина поля зрения — ~6,5 мкм. Фото с сайта sciencephoto.com