Академик В. Н. Пармон
«Химия и жизнь» №5, 2005
Вообще говоря, проблема зарождения жизни некорректна с точки зрения нормальной науки. Действительно, речь здесь может идти лишь о создании более или менее правдоподобной и аргументированной гипотезы либо теории. Проверить же эту гипотезу экспериментально, как того требует серьезная наука, по понятным причинам никто и никогда не сможет... Тем не менее такие гипотезы и теории существуют, о них спорят и пишут. В последние годы — даже российские химики, которым еще недавно было совсем не до того. В частности, об этой проблеме задумались химики из новосибирского Академгородка.
Все теории зарождения жизни можно разделить на две группы: одни постулируют зарождение жизни на Земле, другие — вне Земли. Причем последние появились именно потому, что ни одна из теорий земного происхождения жизни так и не объяснила, как появились первые молекулы РНК и ДНК, без которых современные формы жизни просто немыслимы. Самая известная и хорошо аргументированная гипотеза А. И. Опарина и Дж. Холдейна утверждает, что жизнь возникла в океанах, где перед этим сформировался насыщенный пребиотический бульон из различных органических соединений (рис. 1). Потом из этих соединений под действием вулканов, молний, УФ-излучения Солнца и прочих факторов начали образовываться первые биополимеры, затем фрагменты протоклеток и так далее. В 50-е годы прошлого века С. Миллер пытался подтвердить опытами, что из смеси простых газов в разряде молний могут синтезироваться сложные органические молекулы. Тем не менее ни эти, ни другие теории не объясняют, как в пребиотическом бульоне образовались первые молекулы нуклеотидов, то есть мономеры РНК и ДНК. Случайно они получиться не могли — для этого не хватило бы всех пяти миллиардов лет существования Земли, нуклеотиды слишком сложные молекулы. Теория внеземного происхождения жизни, конечно, временно позволяет расслабиться — ну занесли из космоса готовые молекулы РНК на метеоритах, и дело с концом. Но все-таки опять возникает недоговоренность — а в космосе-то первые молекулы РНК и ДНК откуда взялись?

По геологическим масштабам, жизнь на Земле возникла почти сразу после того, как появилась сама Земля. Ученые считают, что на это потребовалось не более 500–700 млн лет из 4,6 млрд лет существования Земли. В Институте катализа им. Г. К. Борескова СО РАН, расположенном в новосибирском Академгородке, начали проверять теорию, согласно которой образование первых молекул нуклеотидов, а следовательно, и происхождение жизни — явления далеко не случайные. И сейчас идет эксперимент, который уже дал очень интересные результаты (о них позднее). Причем почти одновременно физики из этого же института выдвинули и начали просчитывать новую гипотезу возникновения планет, опирающуюся отчасти на те же физико-химические принципы. И все потихоньку состыковалось и выстроилось в правдоподобную и стройную систему. Конечно, пока еще со множеством нерешенных вопросов.
В космосе
Итак, образование планет Солнечной системы и возникновение жизни на Земле произошли практически в один момент во временных масштабах Вселенной. Поэтому, если бы удалось решить проблему планетообразования, это могло бы стать исходной точкой и для решения проблемы зарождения жизни. Ведь научные вопросы часто решаются комплексно. Но как получить данные о происшедшем событии, если известен лишь конечный результат? Этим занимаются сейчас физики Института катализа и нескольких других институтов СО РАН. Основной инструмент их работы — методы, разработанные для моделирования в лабораторных условиях ядерного взрыва. То есть физики вместе с математиками собирают достаточно надежные данные о каждом процессе, влияющем на картину взрыва, а потом моделируют его на компьютере. Чем мощнее компьютер, чем больше он учитывает процессов и параметров, тем более реальной получается картина. Проверить работу компьютера можно, сопоставив расчеты с наблюдением природных явлений — например, вспышек на Солнце. Естественно, на каждом этапе моделирования надо строго соблюдать все без исключения фундаментальные физические законы, в частности законы сохранения энергии, импульса и т. д.
Сначала сибирские ученые проанализировали наиболее популярные существующие теории образования планет — и остались неудовлетворены. Появилось новое предположение: формирование планет происходило в тех же условиях, в которых каталитические химические реакции органического синтеза. И следующий шаг: именно каталитические реакции привели к возникновению жизни на Земле, а затем — к естественному отбору.
Мы уже давно свыклись с представлением о том, что планеты Солнечной системы зародились из первичного газопылевого облака, окружавшего Протосолнце около 5 млрд лет тому назад. При этом многие ученые предполагали, что планеты Солнечной системы сформировались при столкновении твердых тел. Тем не менее, согласно В. С. Сафронову, если тело величиной порядка тысячи километров столкнется с телом меньшего размера, осколки притянутся к крупному, а вот средние тела (несколько сот километров в диаметре) не могут ни укрупниться, ни поглотиться. Они просто разрушаются при ударе. Что же заставило систему самоорганизоваться и привело к образованию планет земной группы?
Таким механизмом должно быть развитие коллективной неустойчивости, одновременное объединение многих малых тел. Не будем рассматривать в этой статье физическую и математическую часть модели (см. В. Н. Снытников, В. Н. Пармон «Жизнь создает планеты?» — «Наука из первых рук», январь 2004). Скажем только, что модель предполагает формирование солитонов (одиночных волн плотности материи) в околозвездном диске. А мы займемся чистой химией.
Огромное газопылевое облако, вращающееся вокруг Протосолнца, было не просто облаком, а огромным каталитическим реактором, в котором вовсю шли химические реакции и синтезировались разнообразные органические соединения. Почему каталитическим? А потому, что в этом реакторе находились мириады частиц допланетной пыли, содержавшей железо, никель и кремний, — именно они составляли, как полагают, основу первичного газопылевого облака. Там же в избытке находились и газообразные реагенты (основные компоненты межзвездного и межпланетного газа) — водород, угарный газ и т. д. В газопылевом облаке такого состава просто не могли не образовываться органические соединения! Более того, как выяснилось при численном моделировании образования планет, о котором шла речь выше, реакции образования таких соединений были не просто важными, они могли играть решающую роль! Действительно, для слипания твердых частиц «чистой» космической пыли при интенсивном ударе нужен «клей», а образующиеся органические молекулы, закрепляясь на поверхности твердых частиц, образовали необходимый липкий поверхностный слой. Именно он способствовал быстрому соединению пылинок, из которых формировались более крупные частицы, а затем и планетозимали (промежуточные образования) и далее протопланеты. Так, по-видимому, образовались и Земля, и все остальные планеты, астероиды и кометы, которые вращаются сейчас вокруг Солнца.
Надо отметить, что описанная система хорошо знакома специалистам — по классификации химиков-каталитиков глобальный космический реактор относится к разряду реакторов «с псевдоожиженным слоем катализатора». Давление и температура газообразных реагентов в зоне формирования планет были типичными для хорошо известных химических процессов. Другие важные характеристики глобального космического реактора, также близки душе технолога — состав катализаторов, реакционной среды и т. п. Из водорода и угарного газа на железо- и никельсодержащих пылинках были обязаны синтезироваться углеводороды и кислородсодержащая органика — например, простые спирты и альдегиды. Все как в промышленности.
Понятно, что каталитическая активность космического материала — ключевой момент в уточнении теории образования планет. Недавно в Институте катализа мы экспериментально проверили, что вещество реальных метеоритов в самом деле катализирует эти реакции. Каменные и железо-каменные метеориты, которые мы использовали в эксперименте, по своему составу приближаются к веществам, распространенным в космосе. За 4,6 млрд лет своего существования они спеклись и имеют обычную непористую поверхность, но когда-то, в молекулярных облаках и околозвездном диске, межзвездная пыль состояла из частиц нанометрового размера. Поэтому мы испаряли метеориты при помощи лазера и получали порошки с размером частиц 3–4 нм (это примерно соответствует размеру активного компонента в промышленных катализаторах). Затем смотрели, получается ли что-нибудь из смеси угарного газа, водорода и гелия в присутствии метеоритной пыли при атмосферном давлении и температуре до 500°С. В наших экспериментах с достаточно высокими выходами синтезировались этилен и другие углеводороды.
Все это означает, что наши планеты сформировались там, где были условия для каталитического синтеза органических соединений, и что первичные органические соединения, из которых потом могла появиться жизнь, образовались уже на допланетной стадии эволюции Солнечной системы (рис. 2). Интересно, что значительная часть первичного «допланетного» органического вещества могла захватываться при формировании планет (хотя основная масса такого вещества, конечно же, разрушается и развеивается в космосе) и стать впоследствии одним из компонентов ископаемого органического сырья, которым мы сейчас пользуемся. Так это или не так — покажут дальнейшие исследования.

Рис. 2. Схема действия космического каталитического реактора в момент образования Земли
Еще один очень важный вопрос: насколько жестко предопределено образование планет? Оказалось, что самоорганизация вещества по нашей модели идет в одном направлении и поэтому предопределена. Но место появления уединенной волны повышенной плотности вещества, а следом и формирование планеты на данном расстоянии от Солнца — для нашей модели величины случайные. Правда, из общих соображений следует, что если волна плотности окажется на орбите Венеры, то все летучие вещества (вода, водород, органические соединения) там испарятся: планета появится, а жизнь на ней нет. На орбите Марса слишком холодно. Равно как и на орбите Сатурна — недавно выяснилось, что поверхность Титана (спутника Сатурна) состоит в основном из сжиженных легких углеводородов. Планета, на которой живем мы, попала в самую точку. Здесь есть вода, легкие газы и т. п.
Вероятность появления планет на различных расстояниях от Солнца, возможно, определится после дальнейших расчетов, в которых используют дополнительные физико-химические факторы, влияющие на поведение газопылевого облака. Может быть, тогда станет понятно, где проходит граница жизни — до марсианской орбиты или за ней. И это знание сбережет массу средств и сил.
На Земле
Сразу же после того, как сформировались планеты и на их поверхность попало первое «допланетное» органическое вещество (и даже если не попало), на Земле начались те самые физико-химические процессы, которые породили жизнь. Для научной корректности, а также для того, чтобы можно было подсказать геологам, где искать следы первичной протожизни, сформулируем задачу — договоримся, что именно мы понимаем под феноменом «жизнь» и что могло быть ее первым проявлением.
Сегодня ученые считают, что нечто, называемое жизнью, должно отвечать нескольким условиям. Жизнь — это обязательно процесс, то есть функционирование за счет обмена веществом и энергией с окружающей средой. Живые объекты способны к размножению и воспроизведению себе подобных. Наконец, все живые объекты способны к прогрессивной эволюции в сообществе таких же объектов, благодаря наличию у них биологической памяти, способной запоминать признаки, благоприобретенные в ходе естественного отбора по Ч. Дарвину.
Причем должна выполняться вся совокупность условий — любое из них в отдельности не делает объект живым. Таким образом, несмотря на то что все процессы в живых организмах — химические, однако взятая отдельно химическая реакция не является жизнью, так же как и «воспроизведение» себе подобных. Например, рост кристаллов не что иное, как саморепликация подобных соединений и структур. Но это не жизнь. Простой обмен со средой веществом и энергией тоже не есть жизнь. К примеру, основной объект исследования Института катализа — каталитические химические процессы, в основе которых лежит именно обмен веществом объема с поверхностью катализатора. Но ведь и химический катализ — это тоже не жизнь.
Только тогда, когда появляется биологическая память, дающая возможность накапливать наследственную информацию и передавать ее дальше, можно говорить о жизни. Биологическая память — это основа для естественного отбора, в ходе которого организмы усложняются, адаптируются к окружающей среде и эволюционируют. Во всех живых организмах основой биологической памяти служат молекулы РНК и ДНК.
Мы задали себе вопрос: «А возможно ли, что существовали более простые предшественники РНК и ДНК, обладавшие тем не менее свойствами, сходными с молекулярной памятью?» На самом деле очень близки к биологическим автокаталитические системы. Это системы, в которых химическая реакция ускоряется собственными конечными продуктами реакции или предшественниками этих продуктов. Иными словами, в автокаталитических реакциях, как и в сообществе живых организмов, происходит размножение, то есть саморепликация молекул. Простейшая автокаталитическая реакция может быть записана как R + X —> 2 X. После реакции молекулы автокатализатора Х с «молекулой пищи» R получаются две молекулы автокатализатора. При достаточном количестве пищи в системе такое удвоение приводит к лавинообразному росту количества автокатализатора в системе.
А теперь посмотрим, как будет вести себя такая автокаталитическая реакция в открытой системе, где есть обмен веществом с окружающей средой, но количество «пищи» ограничено. Расчеты показывают, что существуют два стационарных состояния такой системы. В первом (неустойчивом) количество автокатализатора точно равно нулю. Это понятно: для того чтобы его концентрация росла, необходима исходная затравка в виде хотя бы одной предшествующей молекулы автокатализатора. Во втором стационарном состоянии концентрация автокатализатора линейно увеличивается с увеличением количества пищи. Но при этом концентрация пищи должна превышать некий минимальный уровень, зависящий от свойств конкретного автокатализатора. Если этого не происходит, то количество автокатализатора также станет равным нулю. Таким образом, для автокаталитических реакций существует критический предел, при котором еда еще есть, а автокатализатор уже исчез, то есть вымер. Причем полностью, до единой молекулы.
Теперь представим, что структура или состав молекулы могут как-то меняться. (Не будем пока называть это мутацией.) Изменение структуры и состава молекулы приводит к изменению ее свойств. Поэтому для каждой измененной молекулы будет своя критическая концентрация пищи. Если концентрация пищи будет уменьшаться до значений ниже критических, то сначала исчезнут те автокатализаторы, для которых эти критические значения количества пищи были максимальны. Если количество пищи снова увеличится, то исчезнувшие типы автокатализаторов не восстановятся уже никогда, потому что исчезли их затравки (рис. 3). Чем это не естественный отбор, аналогичный отбору в биологических популяциях?

Рис. 3. «Естественный» отбор автокатализаторов Xi при уменьшении и последующем увеличении концентрации «пищи» R. Выживает только автокатализатор Х3, у которого критическая концентрация пищи Rкр3 оказалась меньше, чем минимальный необходимый уровень пищи (светлая точка)
Читатель сразу спросит: «А что, уже известны такие автокаталитические реакции с мутациями автокатализатора и с элементами «естественного отбора»?». Известна по крайней мере одна, и довольно хорошо — это так называемая «формозная» реакция Бутлерова, которая была открыта в России почти 150 лет тому назад. Синтез сахаров из молекул формальдегида
n CH2O → (CH2O)n
протекает в присутствии ионов кальция или магния при комнатной температуре в водных растворах. Автокатализаторами в реакции Бутлерова служат синтезируемые в ней же сахара. Интересно, что эту реакцию активно исследовали в 70-х годах прошлого века, поскольку хотели с ее помощью получать искусственную пищу во время длительных полетов на Марс. Но безуспешно: направить реакцию Бутлерова на синтез сахаров какой-либо заранее заданной структуры так и не вышло. Всегда получался целый набор продуктов самой различной структуры, который включал не только полезные, но и ядовитые сахара. Проблему бросили, так и не решив.
Между тем большой набор сахаров (рис. 4) — это не что иное, как прототип мутаций. Изображенные на рисунке сахара одинаковы по атомному составу, но при этом совершенно различны по свойствам. А раз так, то разными должны быть и критические концентрации формальдегида, при которых можно ожидать вымирания сахаров-автокатализаторов при понижении концентрации формальдегида.
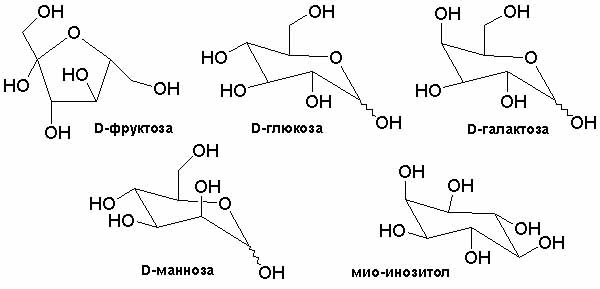
Рис. 4. Структура моносахаров состава С6, которые образуются в реакции Бутлерова
Теперь вспомним, что полные химические названия РНК и ДНК — рибонуклеиновая кислота и дезоксирибонуклеиновая кислота, и корень обоих названий происходит от слова «рибоза». Рибоза — это сахар с пятью атомами углерода, который лежит в основе нуклеотидов, формирующих РНК и ДНК (рис. 5). Нуклеотиды РНК и ДНК отличаются от просто сахаров дополнительными фосфатными группами и азотсодержащими органическими соединениями. Отличие нуклеотидов РНК от нуклеотидов ДНК еще меньше — немного различаются азотистые основания, да в сахарном остатке ДНК не хватает одной гидроксигруппы. Причем если уже есть молекула сахара, то фосфорные и азотные соединения, необходимые для построения нуклеотидов, присоединяются к ней сами без серьезных проблем. Надо отметить, что и другой важный химический компонент живых организмов — переносчик энергии АТФ (аденозинтрифосфат) — тоже содержит моносахарид рибозы. То есть сахара — основа всего живого. И именно молекулы на основе сахаров, а не аминокислот (и следовательно, не белки) ответственны за биологическую память, то есть за отличие живого от неживого.

Рис. 5. Моносахарид рибоза — основа структуры нуклеотидов РНК и ДНК, а также аденозинтрифосфата (АТФ). Показаны структуры сходных нуклеотидов уридила (РНК) и тимина (ДНК)
Для того чтобы экспериментально подтвердить наши предположения, несколько лет назад мы возобновили исследования реакции Бутлерова в Институте катализа. Прежде всего надо было выяснить, какие же типы сахаров — наиболее активные автокатализаторы в этой реакции. Это могло бы подсказать и ответ на вопрос, действительно ли возможен естественный отбор в реакции Бутлерова. Следующий принципиальный вопрос — какую роль в реакции играют ионы кальция или магния и каков механизм их каталитического действия.
Реакцию мы исследуем в довольно простых «проточных» устройствах, которые тем не менее способны моделировать открытые системы с обменом веществ (рис. 6). В стеклянный реактор с хорошо перемешиваемым содержимым подают исходные компоненты, а раствор с продуктами реакции непрерывно вытекает из реактора. Самое важное и сложное — химический анализ того, что получается из формальдегида.

Рис. 6. Схема эксперимента по исследованию реакции Бутлерова в стационарных условиях
Уже первые исследования показали, что далеко не все типы сахаров одинаковы. Кетозы, то есть те из сахаров, в молекулах которых один из атомов кислорода находится в строго определенном месте — активнее, чем альдозы. А самые активные — маленькие сахара с двумя и тремя атомами углерода: гликолевый и глицериновый альдегиды, а также дигидроксиацетон. При этом оказалось, что состав сахаров, которые получаются при стационарном протекании реакции, не зависит от природы вводимого в небольших количествах сахара — затравки. В качестве затравки мы использовали самые разные сахара, но химический анализ на очень чувствительных хроматографах каждый раз показывал, что состав продуктов абсолютно одинаков. Мы обнаружили в продуктах более 14 различных сахаров, но только три из них — глюкоза, сорбоза и эритроза — широко известны. Четыре продукта неизвестны до сих пор, и это вопрос, на который предстоит ответить в ближайшее время. Ответ на него может оказаться очень важным, поскольку биохимики считают, что для образования первичных нуклеотидов не нужна была именно рибоза, все могло начаться и с других ее аналогов. Откуда взялись самые первые молекулы сахаров, которые запустили автокаталитическую реакцию, мы уже знаем. Согласно нашим экспериментам, упомянутые выше простейшие сахара можно получить и без реакции Бутлерова, просто облучив водные растворы формальдегида УФ-светом.
С исходной пищей — формальдегидом тоже все довольно просто. В первичной атмосфере Земли, практически не содержавшей кислорода, формальдегид, несомненно, присутствовал, причем в больших количествах, поскольку возможны несколько путей его образования. Например, он мог получаться в древней атмосфере Земли при мощных разрядах молний или на раскаленных каталитически активных лавах. Хорошо известно, что в этой атмосфере присутствовали водород, оксид углерода, метан, водяной пар, углекислый газ, аммиак. В таких условиях для образования формальдегида достаточно горячей, содержащей железо поверхности.
За эти годы прояснилось, почему для синтеза сахаров нужны именно ионы кальция или магния. Оказалось, что все участники реакции образуют промежуточные соединения — сначала получается комплекс ионов кальция или магния с одной молекулой сахара, а потом к нему присоединяется молекула формальдегида. Возможно образование комплексов с несколькими молекулами сахара. В самые последние месяцы вообще обнаружилась фантастическая вещь. Если вместо катионов кальция взять его соль с фосфат-анионами (обычный природный апатит), то в реакции формальдегида с простейшими сахарами (которые, как было доказано, могли образоваться под действием света) почти селективно (!) получается... долгожданная рибоза! Вот так! Природа, вероятно, очень ловко обошлась малыми средствами. Замечу, что в апатите есть и фосфатные группы, так необходимые для создания нуклеотидов. Входят фосфаты или нет в синтезируемые сахара — покажет будущее.
Нельзя исключить, что реакция Бутлерова — не единственная автокаталитическая реакция, положившая начало естественному отбору и «жизни». Однако другие такие реакции пока не известны — надо искать.
Из всего сказанного можно сделать следующие выводы. Во-первых, в отличие от общепринятого мнения, естественный отбор на Земле мог начаться не с РНК и ДНК, а с гораздо более простых соединений. Во-вторых — и в этом важное отличие нашей теории, пребиотический бульон необязательно должен быть густым, насыщенным разными органическими соединениями (что очень трудно предположить), а, наоборот мог быть, весьма разбавленным — только тогда начиналась конкуренция молекул автокатализаторов за пищу. Я полагаю, что это намного естественнее и правдоподобнее. Более того, первичный синтез сахаров в разбавленном пребиотическом бульоне по целому ряду термодинамических обстоятельств предпочтительнее, чем первичный синтез белков из аминокислот, который обычно обсуждают как первичный элемент зарождения жизни. Ведь при синтезе сахаров из молекул формальдегида не отщепляются молекулы воды, как это происходит при синтезе белков из простых аминокислот. Это значит, что сахара, в отличие от белков, стабильны даже в очень разбавленных водных растворах. Белки же в разбавленных растворах опять распадаются (гидролизуются) на аминокислоты.
Теперь вернемся к мутациям (все-таки воспользуемся этим биологическим термином, поскольку трудно подобрать более адекватное слово) сахаров-автокатализаторов и к тому как закрепляются эти мутации в нашей системе. Полезные мутации в живых организмах крайне редки, и еще реже вероятность их фиксации. Поэтому естественный отбор в живой природе идет очень медленно. В тех условиях, в которых сейчас мы изучаем реакцию Бутлерова, и полезные, и вредные мутации происходят за очень короткие промежутки времени. По-видимому, это считанные минуты, а более точно покажут исследования. В любом случае это совершенно другой масштаб времени и совершенно другая скорость первичного химического отбора, чем предполагалось ранее. Это означает, что первые прототипы живых объектов действительно могли появиться буквально за считанные миллионы, а может быть, даже сотни тысяч лет. Миллиарды лет были для этого не нужны, о чем свидетельствует и геология.
Более того, гипотеза о важной роли автокаталитического синтеза сахаров может пролить свет и на появление хиральности в живом мире. Хорошо известно, что существуют «правые» и «левые» изомеры сахаров и в живых организмах почему-то представлены только правые (а вот аминокислоты, напротив, только левые). Если исходить из нашей гипотезы, то получается, что присутствие в живых организмах именно правых изомеров — случайность. Молекулы практически всех сахаров обладают свойством хиральности, поэтому первая же молекула сахара-автокатализатора, которая имела существенные «эволюционные» преимущества над другими, очень быстро могла съесть остальные. После этого природе уже ничего не оставалось, кроме как использовать эту случайную правую молекулу для построения более сложных, а затем и живых организмов.
Наконец, опираясь на сказанное выше, можно дать более широкое физико-химическое определение понятию «жизнь». Это не формализм. Ведь важно, чтобы мы в поисках примитивной жизни в космосе не прошли мимо того, что следует воспринимать как жизнь или ее первичные проявления. По нашему мнению, жизнь — это фазово-обособленная форма существования функционирующих автокатализаторов, способных к химическим мутациям и претерпевших достаточно длительную эволюцию за счет естественного отбора.
Мы пока не затрагивали вопрос о том, откуда берутся фазово-обособленные формы. Можно надеяться, что исследование реакции Бутлерова и других автокаталитических реакций поможет ответить и на этот вопрос. Однако уже сейчас известно, что такие системы существуют — например, реакции каталитического синтеза полимеров (полипропилена или полиэтилена). Полимер формируется сразу в виде отдельной фазы — микрогранулы или глобулы, внутри которой работает катализатор, наращивающий продукты вокруг себя.
Из всего этого следует еще один заранее неочевидный, но очень важный вывод: если все происходило именно так, как мы говорили выше, то появление жизни на основе именно РНК и ДНК предопределено. То есть других форм жизни появиться не могло. И значит, РНК и ДНК образуются на любой планете, геологическая история которой сходна с историей Земли.
Автор благодарит всех, кто принимает участие в этой работе: сотрудников Института катализа СО РАН, специалистов Института биофизики, физических и математических институтов Сибирского отделения РАН и особенно дипломников, а теперь уже аспирантов Новосибирского государственного университета, которые проводят многие химические эксперименты для подтверждения обсуждаемой гипотезы.


-
Разлагались и вновь синтезировались неорганическим путем.
Нам ведь неважно, весь ли углерод переходил в углеводороды, достаточно и малой его части.-
Если это статья о том, как зародились планеты - да, соглашусь, что это НЕВАЖНО. Но если стать о "Новое в теории появления жизни", то это становится тем самым вопросом, который ставился всегда: "как синтезировались углеводороды в условиях Земли". Т.к. после прибывания Земли в расплавленном состоянии (а эта теория, пока, общепринята) углеводороды какой-либо сложности не могли сохранится. Это, кстати, относится и к ссылке об органических ископаемых в статье.
-
-
По-моему ты не дочитал либо не внимателен! на счет рибозы ниже было написанно: "Если вместо катионов кальция взять его соль с фосфат-анионами (обычный природный апатит), то в реакции формальдегида с простейшими сахарами (которые, как было доказано, могли образоваться под действием света) почти селективно (!) получается... долгожданная рибоза!"
-
В добиотическлй фазе химической эволюции начальные условия абиогенеза это раствор мономеров в воде предполагается готовым. В природе около 500 мономеров аминокислот, точность опустим, нужно учитывать такие, которые могут образовывать полимеры, учитывается оптическая изометрия, главное что бы мономеры способны образовывать при синтезе полимеры. В клетке как сформированной системе самовоспроизводящегося автомата используются 20 протеиногенных аминокислоты с L-изомерами и D-изомеры пентозы в нуклеотидов. Но в природе все изомеры хирально грязные смеси, среда открытая, не в пробирке.
Для объясняя синтез белка, обусловлена необходимость источника информации для такого синтеза. В добиотической фазе такого источника информации не было. В клетки для полноценной работы деления необходимы миллионы белков и десятки тысяч уникальных видов-последовательностей с согласованными функциональными свойствами. В добиотической фазе должны синтезироваться одновременно и очень быстро, учитывая превосходство по скорости экзотермической реакций гидролиза пептидной связи над медленными эндотермическими реакциями дегидратации, помня что ферменты только синтезируются, и ускорить синтез нечем. Это как бы шаг вперед в эволюции и два шага назад. Все миллионы независимых синтезов белков в добиотической фазе должны протекать одновременно и в пространстве липидного микропузыря пространства первичной протоклетки. Ясно, что все события синтеза уникальных полимеров в первичной луже случайные, если конечно гипотезу Бога считать лишней сущностью. Чтобы ускорить эволюцию клетки, в начальных условиях введём допущение, что в растворе первичной лужи, где возникла клетка все 500 видов мономеров растворены равномерно, тогда вероятность присоединения уникального мономера равна 1/500= 0,002. Вероятность 1/20=0,05 возможна только в сверхтехнологичной лаборатории в пробирке, нужно очистить раствор, сделав его хирально чистым и в растворе должны равных пропорций растворить 20 протеиногенных аминокислотных растворов. В природе всегда грязный раствор всех возможных мономеров. Для успешного объясняя синтез белка в добеотической фазе, необходим источника информации для такого синтеза, которым в живой клетке является ДНК. Но в добиотическй период синтеза полимеров благодаря броуновскому движению молекул в жидкости, скачкообразное движение мономеров вероятность скачка уникального мономера к амино концу полимера при 500 разных вариантов мономеров равна 1/500= 0,002. Это можно смоделировать на компьютере настроив генератор случайных чисел на 500 равновероятных чисел. Этот генератор и будет источником информации для уникального синтезе уникальных белков и ферментов. Достаточно знать число необходимых уникальных полимеров необходимых для сборки клетки, в клетки дрожжей по последним исследованиям 42000000 миллиона белков. В первичной клетки для её функционирования как молекулярный самовоспроизводящийся автомат нужны миллионы белков синтезировать с уникальными и согласованными свойствами, и попросить законы природы притормозить гидролиз пептидных связей, чтобы дать фору дегидратации. Так вот для порядочного физика и надеюсь и химика вероятность возникновения в грязной лужи куда меньше вероятности с сотнями миллиардов нулей после запятой. Если в качестве источника информации использовать генератор случайных чисел. А вот непорядочные биологи нейтрализуют сверх малые числа фактически нулевой вероятности своими коэффициентами больших чисел своих пожеланий. Вот почему для абиогенеза гипотеза Бога необходимое сверхестественно условие. Кунин подсчитал вероятность синтеза уникального полинуклеотида длинной 102 мономера один случай на всю нашу видимую вселенную до 50 миллиардов Галактик как наша за 10 миллиардов лет эволюции. И это гвоздь в гроб гипотезе РНК-мира, второй гвоздь это автогидролиз в РНК связей между фосфатами цепи полимера. Жизнь если и могла возникнуть без участия Бога только повторно, поле того как Бог создаст Человека химика, а люди создадут сверх технологическую лабораторию и сверхсуперкомпьютер. Невозможно создать нанороботы, с прямом ми передачей информации с коммуникацией с суперкомпьютером для своевременной доставки уникальных мономеров. Поэтому возникновение жизни это будет всегда почти абсолютным чудом Божьим с вероятностью много большей многих сотин миллиардов девяток после запятой, фактически с точки зрения физики равна 1, фактически является сверхъестественным явлением во Вселенной.
Нужна работоспособная универсальная модель. Она дудет представлена в 2013 г. предварительно с идеей и концепцией можно познакомиться на блоге http://dna-space-life.blogspot.com/ Будет дан ответ на ключевой вопрос - Почему природа выбрала нуклеотид дезоксирибозы в качестве главглго и единственно приемлимого строительного материада для ДНК.











Рис. 1. Зарождение жизни в потоках энергии и химических веществ по Опарину—Холдейну