В середине юрского периода птерозавры встали на четвереньки

Птерозавры, гигантские летающие рептилии мезозойской эры, доминировали в небесах нашей планеты по меньшей мере 150 миллионов лет (с позднего триаса до конца мела). Множество особенностей — от облегченного скелета до специфического строения конечностей — указывают на глубокие адаптации к воздушному образу жизни. Однако, как показывают многочисленные следы, сохранившиеся по всему земному шару, с определенного времени эти рептилии начали активно осваивать и наземное перемещение, чувствуя себя на четырех конечностях не менее комфортно, чем пролетая на широко распахнутых крыльях над головами динозавров. В новом исследовании, опубликованном в журнале Current Biology, международный коллектив исследователей смог связать окаменелые следы с тремя группами птерозавров, которые их оставили. Судя по полученным результатам, птерозавры начали адаптироваться к передвижению на четырех лапах еще со средней юры, и к позднему мелу часть из них превратилась в полуназемных хищников, активно ищущих корм на суше.
Несомненно, главенствующими летающими позвоночными мезозоя были птерозавры, отличавшиеся огромным разнообразием форм и размеров. Поскольку анатомически и, очевидно, физиологически эти животные были глубоко адаптированы к летающему образу жизни, палеонтологи в первую очередь уделяли внимание их летным качествам (см., например, Полет птерозавров в ходе эволюции неуклонно совершенствовался, «Элементы», 02.11.2020; Крылатые гиганты прошлого были неспособны к динамическому парению, «Элементы», 27.05.2022), считая способности к наземному передвижению невысокими и связывая их образ жизни в первую очередь с морскими экосистемами (P. Wellnhofer, 1987. Terrestrial locomotion in pterosaurs).
Однако на рубеже тысячелетий начали накапливаться свидетельства в пользу того, что птерозавры много и охотно передвигались по земле, забираясь вглубь суши и оставляя там свои характерные отпечатки (D. Unwin, 1996. Pterosaur tracks and the terrestrial ability of pterosaurs; J. Mazin et al., 2003. Ichnological evidence for quadrupedal locomotion in pterodactyloid pterosaurs: trackways from the Late Jurassic of Crayssac (southwestern France); M. Lockley et al., 2008. The fossil trackway Pteraichnus is pterosaurian, not crocodilian: Implications for the global distribution of pterosaur tracks). Также последовали переоценки морфологии скелета, разнообразия питания и летных качеств, что показало, насколько широкий спектр экологических ниш занимали крылатые рептилии на мезозойской Земле (M. Witton, D. Naish, 2008. A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology; J. Bestwick et al., 2020. Dietary diversity and evolution of the earliest flying vertebrates revealed by dental microwear texture analysis; M. Witton, M. Habib, 2010. On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness). По-видимому, упор на развитие полетных характеристик не ограничил совершенствование альтернативных способов перемещения, и наравне с динозаврами в сухопутных экосистемах птерозавры выполняли роль как мелких хищников, так и добычи.

Рис. 2. Следы птерозавров легко отличить от следов динозавров: их задние ноги оставляли отпечатки четырех прямых узких пальцев, передние — трех небольших когтей на сгибе крыла. Здесь представлена ктенохазма (Ctenochasma elegans). Рисунок из обсуждаемой статьи
За последние тридцать лет было найдено множество новых образцов следов птерозавров, причем в некоторых местах их явно больше, чем ископаемых костей (J. Jung, M. Huh, 2024. New Pterosaur Tracks from the Hwasun Seoyuri Tracksite (Turonian) of South Korea: Implications for their Ecological Niche and Habitat). Это порядком затрудняет определение принадлежности следов хотя бы до уровня семейства (см. картинку дня Следы невиданных зверей), и в результате даже активное накопление ископаемого материала дает не так много сведений о биологии таксонов. Обнаруживая следы крылатых ящеров, ученые приходят к выводу лишь о присутствии неопределенных птерозавров в регионе в какой-то период времени, но не используют эти сведения при изучении биогеографии или макроэволюции конкретных клад (Y. Yu et al., 2023. Complex macroevolution of pterosaurs).
Конечно, на то есть веские причины, и в первую очередь это сложности с отнесением нового ихнотаксона к конкретной кладе птерозавров. Чаще всего ископаемые отпечатки относят к продвинутым птерозаврам-птеродактилоидам (В Аргентине обнаружили переходное звено между примитивными и прогрессивными птерозаврами, «Элементы», 13.12.2024), хотя этот подход не общепринятый и может потребовать пересмотра (M. Witton, 2015. Were early pterosaurs inept terrestrial locomotors?). Редкие попытки отнести отпечатки к конкретной группе основываются на косвенных доказательствах, таких как абсолютный размер следов или их стратиграфический возраст (K. Hwang et al., 2002. New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea), либо же на ассоциации следов с костными остатками, которые и считаются «следообразователем» (Y. Li et al., 2025. First deciphering of large pterosaur footprints and their trackmaker in the Junggar Basin, China).

Рис. 3. Разнообразие следов птерозавров, найденных по всему миру. Рисунок из статьи Y. Li et al., 2021. A new pterosaur tracksite from the Lower Cretaceous of Wuerho, Junggar Basin, China: inferring the first putative pterosaur trackmaker
В новом исследовании ученые из Великобритании и США использовали 3D-моделирование, подробный количественный анализ и сравнение следов с костными остатками, чтобы выделить ключевые синапоморфии трех крупных групп птерозавров. Работа была проведена на 52 образцах, обнаруженных на всех континентах за исключением Австралии и Антарктиды; возраст отпечатков составлял от 161 до 66 миллионов лет.
В первую очередь палеонтологи сосредоточились на отпечатках стоп, поскольку отпечатки передних конечностей птерозавров сравнительно консервативны по форме и несут меньше таксономически ценной информации. Учитывая, что все сохранившиеся отпечатки передних конечностей отличаются чрезмерно длинным третьим пальцем, авторы пришли к выводу, что отпечатков таксонов с более пропорциональными пальцами (например, анурогнатид; см. картинку дня Насекомоядный птерозавр) пока найдено не было.
Судя по сохранившемуся образцу птерозавра с мягкими тканями, у этих ящеров были «тощие лапки» с минимальным количеством мягких тканей вокруг ступни и пальцев, что характерно и для современных летающих позвоночных — птиц и рукокрылых (E. Frey et al., 2003. A new species of tapejarid pterosaur with soft-tissue head crest). Это облегчает работу ученых, которым приходится закладывать меньшие поправки на разницу между скелетной основой и следом живого птерозавра, и повышает точность измерений. Проанализировав все имеющиеся следы, исследователи отнесли их к поздним птерозаврам-птеродактилоидам, а также выделили три морфотипа следов, каждый из которых соотносится с конкретной кладой птеродактилоидов.

Рис. 4. Три морфотипа следов птерозавров и их распределение по континентам (отдельно выделены конкретные формации) и времени существования. Обратите внимание, что у всех морфотипов на отпечатке передней конечности третий палец (нижний) заметно длиннее остальных. Рисунок из обсуждаемой статьи
Морфотип I был отнесен к археоптеродактилоидам (Archaeopterodactyloidea), или ктенохазматоидам — ранним птеродактилоидам, существовавшим с конца юрского по ранний меловой периоды. Их остатки обнаружены в Африке, Евразии и Северной Америке. Многие виды этой клады отличались необычным строением черепа и зубов, в частности, именно к ктенохазматоидам относится птеродаустро (Pterodaustro guinazui; см. картинку дня Птеродаустро и его челюсть-щетка). Для морфотипа I характерны сравнительно крупные отпечатки стоп почти треугольной формы, длинные плюсневые кости и короткие пальцы. Первоначально эти следы считали принадлежащими крокодиломорфу, но затем была доказана их принадлежность птерозавру (K. Padian, P. Olsen, 1984. The Fossil Trackway Pteraichnus: Not Pterosaurian, but Crocodilian; M. Lockley et al., 2008. The fossil trackway Pteraichnus is pterosaurian, not crocodilian: Implications for the global distribution of pterosaur tracks). От отпечатков других птерозавров отпечатки морфотипа I отличаются соотношением длин плюсневых костей: первая длиннее второй, вторая длиннее третьей, третья длиннее четвертой. Такое распределение крайне нетипично для диапсидных рептилий, включая крокодиломорфов, у которых первая плюсневая кость сравнительно короткая.
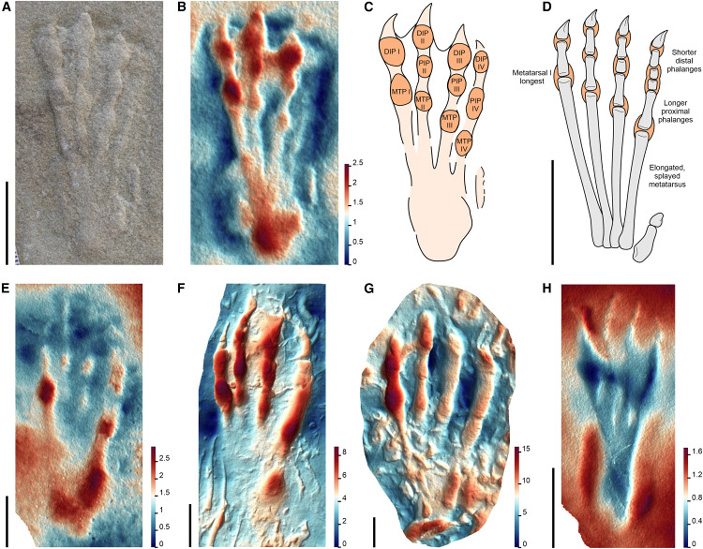
Рис. 5. Морфотип I. A–D: фотография позднеюрского естественного слепка отпечатка стопы Pteraichnus isp.; цифровая модель, показывающая глубину отдельных частей следа и расположение пальцевых подушечек; прорисовка следа и прорисовка костей стопы с выделенными синапоморфиями. E–H: примеры следов морфотипа I из США (E, F) и Европы (G, H). Рисунок из обсуждаемой статьи
Морфотип II отнесли к джунгариптеридам (Dsungaripteridae), семейству птерозавров из раннего мела Евразии и Южной Америки. В данном исследовании рассматривались два ихнотаксона из раннего мела Китая, Pteraichnus wuerhoensis и Pt. junggarensis. Эти следы отличались от следов морфотипа I относительными длинами плюсневых костей (кость I примерно равна кости II, и обе сравнительно короче, чем в отпечатках ктенохазматоидов) и костей пальцев (первый палец укорочен, остальные три примерно равны по длине, в то время как у морфотипа I все четыре пальца примерно одинаковы, см. рис. 2). Такое соотношение костей пальцев характерно для джунгариптерид Dsungaripterus weii и Noripterus complicidens и птеранодонов Pteranodon, однако пропорции плюсневых костей у последних отличаются, что позволяет отнести отпечатки морфотипа II именно к джунгариптеридам. Впрочем, поскольку результаты анализа основаны на крайне ограниченной выборке (четыре отпечатка и одна полностью сохранившаяся стопа джунгариптерида), воспринимать их следует с некоторой осторожностью.

Рис. 6. Морфотип II. Фотография раннемелового естественного слепка отпечатка стопы Pteraichnus wuerhoensis (A), цифровая модель, показывающая глубину отдельных частей следа и расположение пальцевых подушечек (B); прорисовка следа (C); прорисовка костей стопы с выделенными синапоморфиями (D). Рисунок из обсуждаемой статьи
Наконец, следы морфотипа III были наиболее распространены географически и были отнесены к неоаждархиям (Neoazhdarchia), подгруппе аждархоидов (Azhdarchoidea), крупных меловых птерозавров со сравнительно короткими крыльями. Следы такого морфотипа отличались узкой удлиненной стопой со слабыми отпечатками пальцев и хорошо отпечатавшейся пяткой, на которой виден след пяточной подушечки — явной адаптации к наземному передвижению. Соотношение плюсневых костей у следов такого морфотипа отличается от предыдущих (плюсневая кость I примерно равна по длине плюсневой кости II или короче ее), и все плюсневые кости тесно сжаты. Поскольку хорошо сохранившиеся отпечатки такого морфотипа — большая редкость, а развитые подушечки затрудняют оценку длин плюсневых костей, авторы рекомендуют при определении принадлежности таких следов использовать явную синапоморфию — хорошо отпечатывающуюся пятку с развитой подушечкой.

Рис. 7. Морфотип III. Цифровая модель раннемелового естественного слепка отпечатка стопы Pteraichnus yanguoxiaensis (A), прорисовка следа (B) и прорисовка костей стопы с выделенными синапоморфиями (C). (D) и (E) — левая и правая ноги неопределенного аждархоида (образец SMNK PAL 3830) с сохранившимися пяточными подушечками. Рисунок из обсуждаемой статьи
В настоящее время наши знания о следах птерозавров все еще очень ограничены: к примеру, к ихнотаксону Pteraichnus относятся следы, которые в данном исследовании отнесены к разным морфотипам и, соответственно, кладам птерозавров. Используя примененную авторами методологию, можно уточнять время существования, экологию и особенности поведения представителей конкретных клад, расширяя наши знания о биологии птерозавров.
Несмотря на относительно многочисленную выборку, ни один из изученных отпечатков не был с уверенностью отнесен к ранним птерозаврам-рамфоринхоидам (Rhamphorhynchoidea), для которых характерен хорошо развитый пятый палец стопы, сопоставимый по размерам с остальными. Все задокументированные следы относятся к временному периоду от средней юры до позднего мела (а появились первые птерозавры еще в триасе), причем костные остатки первых птеродактилоидов несколько моложе древнейших следов морфотипа I (C. Zhou et al., 2017. Earliest filter-feeding pterosaur from the Jurassic of China and ecological evolution of Pterodactyloidea). Впрочем, это соответствует результатам филогенетических исследований, предполагающих появление птеродактилоидов в среднем юрском периоде.
Можно предположить, что рамфоринхоиды в основном лазали по деревьям и скалам, не спускаясь, к примеру, на полосы прибрежной грязи и ила, в которых часто сохраняются ископаемые отпечатки, тогда как у первых птеродактилоидов развились стопы и кисти, лучше приспособленные для передвижения по земле, — и в ископаемой летописи появились первые следы птерозавров (R. Smyth et al., 2024. Hand and foot morphology maps invasion of terrestrial environments by pterosaurs in the mid-Mesozoic). Стоит, правда, отметить, что не все птеродактилоиды были хорошими ходоками: у представителей некоторых групп, например, аньянгуэрид (Anhangueridae), были редуцированные кости таза, сравнительно короткие ноги и маленькие ступни, так что вряд ли они часто оставляли свои отпечатки в палеонтологической летописи.
Судя по распределению следов, ктенохазматоиды были неотъемлемой частью прибрежных морских экосистем и, судя по обилию отпечатков, в свое время буквально кишели на отмелях, отфильтровывая из воды мелкую живность или охотясь за рыбешкой. Джунгариптериды, в свою очередь, тяготели к мелководным рекам и озерам, где, судя по их характерным челюстям с беззубыми кончиками и крепкими зубами, искали моллюсков с твердыми панцирями. Наконец, следы неоаждархий обнаруживаются исключительно в континентальных отложениях до самого конца мелового периода, после чего они начинают попадаться в прибрежных регионах. Любопытно, что в тех местах, где следы неоаждархий сохраняются вместе со следами сухопутных позвоночных, направление следовых дорожек птерозавров совпадает с дорожками динозавров, которые, натолкнувшись на водоем, воспринимали его как препятствие и обходили по краю — то есть, даже натолкнувшись на преграду, птерозавры не поднимались на крыло, а предпочитали обойти ее пешком (L. Xing et al., 2015. Reanalysis of Wupus agilis (Early Cretaceous) of Chongqing, China as a Large Avian Trace: Differentiating between Large Bird and Small Non-Avian Theropod Tracks).
Подводя итоги, с помощью количественного анализа и выделения ключевых синапоморфий палеонтологи теперь смогут определять следы птерозавров не до уровня отряда и подотряда, но до более узкой клады, что позволит больше узнать об их экологии, а также географическом и временном распределении. Судя по полученным данным, птерозавры начали активно вторгаться в наземные экосистемы уже в среднем юрском периоде, адаптируясь к передвижению по суше, что в конечном итоге привело к появлению «цапель-монстров» позднего мелового периода (см. картинку дня Крылатые чудовища древней Трансильвании), наводивших ужас на мелких динозавров и других сухопутных позвоночных по всему земному шару.
Конечно, ни одно летающее позвоночное животное не проводит жизнь исключительно в воздухе, спускаясь на землю хотя бы для размножения. Возможно, ранние птерозавры, подобно современным мелким воробьиным вроде чижей или клестов, в основном отдыхали и размножались на скалах и ветвях деревьев, откуда им было удобно взлетать, спрыгивая прямо в воздух: об этом свидетельствуют их цепкие кисти с крупными когтями и относительно короткие стопы с длинными хватательными пальцами. Поздние птерозавры начали больше времени проводить на земле, как нелетающие четвероногие, чему способствовали более длинные ноги, редукция хвостовой перепонки, удлиненные и крепкие кисти и стопы: подобно куликам или голубям, они могли искать пищу на земле, поднимаясь на крыло только в случае опасности или при необходимости добраться до удаленного места (R. Smyth et al., 2024. Hand and foot morphology maps invasion of terrestrial environments by pterosaurs in the mid-Mesozoic).
Наконец, стоит отметить, что активно передвигаться по суше птерозавры могли начать еще в раннем мезозое. К примеру, британский палеонтолог Марк Уиттон (Mark Witton) отмечал, что отсутствие следов птерозавров с триаса по среднюю юру может и не отражать их привычки в передвижении (M. Witton, 2015. Were early pterosaurs inept terrestrial locomotors?): остатков крылатых рептилий из этого временного периода в принципе меньше, чем многих современных им четвероногих, что может быть следствием особенностей экологии (например, птерозавры в основном заселяли леса и горы, где остатки сохраняются плохо) или биогеографии (на ранних этапах своей эволюционной истории птерозавры могли быть распространены нешироко). Вдобавок, ранние птерозавры были в основном некрупными животными размером от галки до орла, так что их следы были невелики и неглубоки: возможно, мы их просто еще не нашли, а если и нашли, то не распознали?
В таком случае обилие следов птеродактилоидов во второй половине мезозоя — это не только следствие их анатомической приспособленности к наземному передвижению, но и результат активного освоения прибрежных и речно-озерных экосистем, где они искали корм подобно современным куликам. Рамфоринхоиды же — в основном насекомоядные и рыбоядные виды, искавшие добычу в открытом море, — с меньшей вероятностью стали бы бродить вдоль линии прибоя или по берегу озера, оставляя следы в иле или мягкой грязи. Так что, удивляясь отсутствию их ископаемых следов или объясняя это исключительно анатомическими ограничениями, мы удивляемся отсутствию следов двугорбого верблюда в песчаных дюнах или на дне высохшей австралийской реки.
Источник: R. S. H. Smyth, B. H. Breithaupt, R. J. Butler, P. L. Falkingham, D. M. Unwin. Identifying pterosaur trackmakers provides critical insights into mid-Mesozoic ground invasion // Current Biology. 2025. DOI: 10.1016/j.cub.2025.04.017.
Анна Новиковская
-
Ну, страусы же крыльев лишились, почему бы и нет? Другое дело, если эти птерозавры перестали летать, то скорее всего и кожаные перепонки у них на крыльях редуцировались, ибо вещь это достаточно тонкая и уязвимая...
-
ответ на Ваш коммент дали ещё в 50х годах физики (по легенде это был Ландау, хотя называют и другие имена) в дискуссии с Лысенко: девственную плеву рвут, рвут уже давно - но это не приводит к её исчезновению.
Так и Ваше "скорее всего" происходит от бытового ламаркизма.
-


Всё, как раз таки, наоборот - более плотная среда позволяет (и вынуждает) иметь более плотные и меньших размеров крылья (плавники). А причиной тому - увеличение сопротивления среды и давления её потоков: при более плотной среде труднее махать такого же размера крыльями (плавниками) и тяжелее сопротивляться её потокам (ветрам и течениям).
Парусообразные (легкие и большой площади) - как у птерозавров - крылья выгоднее иметь, как раз таки, при меньшей(!) плотности воздуха. При увеличении плотности воздуха, потоки ветра легко сносили бы (и, вообще, уносили бы прочь) перепончатокрылых птерозавров, затрудняя им полет.
Еще бОльшие проблемы более плотная атмосфера доставила бы летающим насекомым - потому что у них другая механика полета - они не парят, как птицы, а часто машут своими плоскими крыльями. Гигантские стрекозы, меганевры пообломали бы себе полуметровые крылья, если бы смогли ими часто махать в еще более плотном, чем сейчас воздухе. Скорее, наоборот, воздух тогда был более разряженным, если позволял им часто - как требуется насекомым для обеспечения подъемной силы и маневренности - махать их огромными крыльями. Можно почувствовать на себе усилие, которое надо было бы прилагать меганевре при современной плотности воздуха - просто часто-часто помашите веером.
Поэтому, именно увеличение плотности атмосферы поспособствовало завоеванию неба птицами, с их более жесткими и относительно меньших (если не считать оперение) размеров крыльями. По-видимому, это также поспособствовало распространению цветковых растений, опыляемых в т.ч. ветром.
Таким образом, резюмируем, что при более плотной атмосфере необходимо иметь более плотные и жесткие (а значит и более тяжелые) и меньших размеров (чтобы снизить сопротивление среды) крылья (почти плавники), которые не увеличивают подъемную силу и не дают никакого стимула для увеличения общих размеров тела по сравнению с современными летунами. При этом, очевидное преобладание, во времена динозавров, летунов с легкими парусообразными крыльями (выгодными и необходимыми летунам только при более разряженной среде) и гигантсткие размеры самих летунов подталкивают на мысль, о том, что одновременно бОльшие размеры летунов и меньшая плотность воздуха (в котором они могли бы махать своими гигантскими хлипкими парусообразными крыльями) могли обеспечиваться только за счет меньшей(!), чем сейчас, гравитации. Всё говорит в пользу этого.
Ведь, при меньшей гравитации плотность атмосферы уменьшается, а это снижает подъемную силу. Значит, для при увеличении общей массы и размеров тела летуна для обеспечения достаточной подъемной силы последней необходимо относительно увеличить площадь крыльев, не увеличивая значительно их массу, т.е. приобрести парусовидные (большие, но легкие) крылья.
PS. Если бы при массе тела 200 кг можно было бы летать в условиях современной гравитации, то человек уже давно бы, с помощью современных технологий и сверхлегких искусственных материалов, создал летающий аппарат, работающий только на мускульной силе, способный легко взлетать и переносить его на расстояния, которые преодолевали гигантские птерозавры. И покорил бы с их помощью небо.
-
Ну такой-то бред писать не надо! Маленькие насекомые как раз живут в условиях высокой вязкости воздуха (для них число Рейнольдса сильно меньше) и летать им легче. А маленькие парашютики семян и пауки на паутинке вообще без проблем летают даже ничем не маша, хотя сильно тяжелее воздуха. В разреженной атмосфере, когда вязкость мала, они бы просто лежали на земле. Про страшную плотную воду, создающую ужасное препятствие для подводных летунов я вообще молчу
У человека печальное количество мышц к общему весу тела - мы даже бегать толком не можем, сильно уступаем волкам и прочим в ваттах на килограмм тела. На мускулах мы никуда не улетим. Была бы среда поплотнее - может и смогли бы. При 1000 кг/м3 мы летаем даже без мускулолета-
число Рейнольдса сильно меньше
наоборот, больше. Число Re для насекомых очень велико, и они летают по вихревой модели.-
Нет. Для них числа Рейнольдса малы, а вязкость воздуха велика. По какой модели они летают, не знаю, но число Рейнольдса для них ниже чем для птеродактиля
-
Вязкость одинакова для всех, не так ли? Это свойство воздуха, а не объекта.
При этом в формуле для Re есть "характеристический размер" в знаменателе. Чем меньше размер - тем больше Re при прочих равных условиях.-
Вы заблудились в трех соснах. Бывает.
В знаменателе формулы для числа Рейнольдса стоит только вязкость среды.
Иногда стоит сверять свои озарения с википедией.-
путаница в слове "высоких". Высокие Re - это больше критерия Рейнольдса, а не большое значения критерия. Чем выше вязкость - тем меньше турубулентность. Чем меньше характеристический размер - тем выше. Так понятно?
Мелкие насекомые - летают именно по вихревой модели и именно по причине малого своего размера. Примеры для случая трубы не показательны, почему - разъяснить?
Я, хоть и 45 лет назад, но гидроаэромеханику учил, так что нет, не по вики. Объяснение границы турбулизации потока можно сделать буквально на пальцах. Формула - эмпирична, несколько различна для разных случаев и используется в конкретных условиях для оценочных суждений, а не всегда для всего и точного расчёта. К примеру, погранслой может быть турбулентным, а обтекание самолёта в целом - нет. При одной и той же скорости в одном и том же воздухе одной и той же вязкости.-
Чем выше вязкость - тем меньше турубулентность. Чем меньше характеристический размер - тем выше. Так понятно?
Совершенно верно. Вязкость в знаменателе, а характеристический размер в числителе.Мелкие насекомые - летают именно по вихревой модели и именно по причине малого своего размера.
Непонятно, что Вы понимаете под вихревой моделью. Крыло самолета создает подъемную силу за счет образования вокруг него огромного присоединенного вихря. Самолеты летают не по вихревой модели?
Хотя конечно, вихри у крыла насекомого и у крыла самолета разные.
-
-
-
-
-
-
Теперь все встало на свои места!
Родная планета была легче, искривляла пустоту меньше, слой атмосферы был тоньше, плотность её меньше, концентрация кислорода жиже. Стойкость против солнышка хуже, мутации размашистее и чаще; погода стабильнее, потому как температура кипения воды ниже, а значит облачность гуще и ниже...
Что там ещё должно по этим причинам быть не такое, как, сегодня?
То есть эти крылатые ребятки с очень прикольным дизайном дают возможность понять, что тогда было тут практически инопланетное сообщество!
Надо переописать-переобдумать и конструкцию всех зверей тех времен!
Оч плодотворное соображение!
Кажется, должно быть сегодня возможным посчитать конкретную плотность атмосферы и кривизну пустоты в те времена, исходя из формы и размеров этих гиганских летучих.
А потом посчитать годовой прирост массы Земли, который привёл к сегодняшней массе, сегодняшней плотности атмосферы, сегодняшней кривизне пространства.
Отличный фронт работ!
Интересно, а как Луна себя вела в той кривизне? И земная орбита в системе?
Как много пользы несут кожистые крылышки птерозавров для понимания природы!
-
Планета не была легче. Было легче все то, что расположено на её поверхности. По закону всемирного тяготения это зависит от расстояния от центра Земли и гравитационной постоянной G (величина которой не очень-то и постоянна - судя по её измерениям)
Что за глупость(!) в комментарии (не вашем) -И что чудесно, выведи за скобки сей "гравитационной гипотезы" этих гигантов, и... нет "гипотезы" ))
Очевидно же, что первыми на ком могло сказаться увеличение гравитации - это именно гиганты, особенно сухопутные. Какое рыбам дело до изменения гравитации?)
Хотя и их это, очевидно, КОСнулось) Увеличение веса/давления воды, судя по всему, и привело к распространению в океанах, в тот период, костистых рыб, имеющих более прочный скелет.
Так что, да, условия среды могут (и, наверное, должны) определять внешний вид и строение тел её обитателей. Сухопутным плавники ни к чему ;)
"Странные", однако, совпадения имели место в один период - вымирание динозавров и всей гигантской флоры и фауны, появление/распространение опыляемых ветром цветковых растений, распространение костистых рыб в морях и океанах, появление/распространение птиц, переход в водную среду обитания предков современных китообразных и многое (долго перечислять) другое. И всё это из-за падения какого-то, там, астероида/кометы?) Коих падало на Землю, и раньше, и позже немало штук. Или из-за глобального похолодания в результате его падения?)
А гравитационная Гипотеза прекрасно работает со всем перечисленным и многим, здесь, не перечисленным.-
Это Вы бросьте меня путать: как раз из гравитации исходя и нужно чтоб планета была менее массивной, иначе звери ну никак легче не могут стать. Откудова эта вспомянутая Вами "жи"? Из массы планеты берется!
Логика такая: чтоб громадные животные полегчали - нужно уменьшить силу тяжести, т.е. массу планеты.
Сегодня"жи" стала больше, и такие громадины не живут.
Вывод:
Земля за довольно короткий период времени ощутимо прибавила в массе, что привело к росту "жи" и вымиранию этаких крупненьких животинок.
Я помню, что второй (кроме массы) фактор - расстояние. Но представить, что наша планета за это время сильно уменьшилась в диаметре, мне как-то жутче, чем то, что она прибавила в массе. Массу хоть понятно где взять - из метеоритов и прочих болидов.
А как диаметр уменьшить? Это уж апокалипсис какой-то!...
-
-
1) Физика говорит на языке математики, все просчитывается и доказывается, наблюдается, в экспериментах проверяется. Ничего подобного мы от вас за годы не видели и не увидим.
2) Гипотетическое изменение гравитации (особенно в 2-а раза, как вы не раз предлагали) это явление астрономического масштаба с соответствующими проявлениями в изменении орбит и скорости вращения небесных тел (не говоря о том, что даже размер звезд и их эволюция изменятся). Соответственно, изменится орбита Земли, Луны, скорость их вращения, а это влияет на такое масштабное планетарное явление, как приливо-отливные циклы в океане. Что в свою очередь повлияет на ритмичность приливо-отливных отложений. Такие циклы вполне хорошо фиксируются в осадках и даже в ритмичности роста раковин моллюсков, и изучаются. По ним рассчитывают продолжительность суток, лунного месяца и года и пр. И эти исследования ставят огромный крест на ваших бредовой фантазиях об изменение гравитации или орбиты Земли.
https://agupubs.onlinelibrary.wiley.com/doi/full/10.1029/2019PA003723
https://www.researchgate.net/publication/266208982_Late_Cretaceous_diurnal_tidal_system_A_study_from_Nimar_Sandstone_Bag h_Group_Narmada_Valley_Central_India
https://agupubs.onlinelibrary.wiley.com/doi/epdf/10.1029/1999RG900016
https://www.researchgate.net/publication/266208982_Late_Cretaceous_diurnal_tidal_system_A_study_from_Nimar_Sandstone_Bag h_Group_Narmada_Valley_Central_India
https://www.researchgate.net/publication/378861769_Bayesian_Estimation_of_Past_Astronomical_Frequencies_Lunar_Distance_a nd_Length_of_Day_From_Sediment_Cycles
https://www.mantleplumes.org/WebDocuments/MazumderESR2004.pdf
https://sci-hub.ru/10.1016/j.sedgeo.2013.08.001
https://sci-hub.ru/10.1016/j.earscirev.2009.09.002
Судя по комментариям за все годы, вы осознаете что пишете чушь. На одной теме так странно развлекаться несколько лет подряд, писать пространные скатерти текстов... Может стоит заняться чем-то более конструктивным, тем что вас возвысит в глазах окружающих, а не как сейчас низводит до сумасшедшего?
увеличение сопротивления средыЭто влияет лишь на скорость. На подъёмную силу увеличение плотности влияет положительно.
-
Подъемная сила зависит от скорости набегающего потока воздуха). Т.е., если скорость (например, взлетная) уменьшается, то уменьшается и подъемная сила.
Предполагаю, что увеличение плотности воздуха увеличивает подъемную силу ровно на столько же, на сколько увеличивает сопротивление среды. Т.е. никакого выигрыша оно не даёт, в частности, для увеличения размеров и массы тела. Единственное, на что может повлиять - это на относительные размеры крыльев, т.е. по отношению к размерам тела. Что, по сути, означает уменьшение общих размеров тела при постоянной его массе.
Хотите оценить усилие, которое должен был бы прилагать птерозавр с размахом крыльев в 12 метров при обычной плотности атмосферы? Вы когда-нибудь катались на виндсерфере - доске с парусом? Высота паруса около 3-4 метров. Площадь паруса, думаю, не больше, чем у одного(!) крыла гигантского птерозавра. Вот, усилие от ветра, который давит на парус виндсерфера, я думаю, сопоставимо со взмахом крыла птерозавром. И оно, судя по всему, немалое, если серфингист перебарывает его лишь весом своего тела, а не силой/мышцами рук.
А, если еще больше увеличить плотность атмосферы, то тут понадобятся два серфингиста, на одну доску, перетягивающих парус виндсерфера (= одно крыло птерозавра) всем своим весом. )
Может быть причина в том, что атмосфера стала пожиже?Сила гравитации была слабее процентов на 80, люди на дом запрыгивали с разбегу. Ягоды росли на опушке с кота размером. Коты были с собаку, собака с корову, а корова как цех, а в цеху мужики в шахматы играли по методике ботвинника - конём мат ставили с первого хода! И атмосферное давление было 8 атмосфер.
-
Погодите... 8 - это же больше одной!
То исть атмосфера гуще, крыльями большими не помахать особо...
Так что было-то, давление меньше и меньше кривизн, или давление больше с большей кривизной Пространственно-Временного Континуума?
У Вас как-то нестройно - давление выше, масса меньше...
Доработайте, пожалуйста, Вашу идею)-
То исть атмосфера гуще, крыльями большими не помахать особо...
им и не надо особо махать, потому что каждый взмах в 8 раз эффективнее.У Вас как-то нестройно - давление выше, масса меньше...
у меня нет ничего про массу, может вес?Доработайте, пожалуйста, Вашу идею)
исправьте вашу школьную ошибку-
Я о гравитации. То есть об искривлении. Которое, как нынче говорят, с массой как-то там связано.
А раз кривит меньше (прыгать легче, а значит и масса планетки меньше) , то и низкое давление атмосферы должно наблюдаться бы.
Вы же констатировали лёгкость прыгать и высокое давление одновременно - сие противоречиво.
Потому и отправил на доработку)
А про искривление пространства, так
в школе моей этого не было (дальше Ньютона с его притягиваемым по-старинке яблоком мы в ней не ходили), посему исправить по Вашей просьбе тут ничего не смогу.
Будьте снисходительны)
-
-
-
Потому что
(А) раньше были млекопитающие сильно крупнее слона, но вымерли, однако-с. Максимальный вес млека на суше - порядка 20 тонн, современные слоны, спасибо людям и их ружьям, едва до 10 успевают дорасти;
(Б) динозавры были куда лучше "заточены" под гигантизм, в первую очередь благодаря пневматизированным (легким и прочным) костям и воздушным мешкам, помогавшим снабжать кислородом всю эту махину. К примеру, диплодок длиной около 25 метров весил бы всего 15-20 тонн, то есть был бы сопоставим с откормленным индрикотерием;
(В) крупнейшие динозавры, скорее всего, не были теплокровными в нашем понимании этого слова, то есть не тратили львиную долю получаемой из пищи энергии на самообогрев. Современный саванный слон тратит на сбор своей ежедневной порции корма 16–18 часов в сутки, а сколько в таком случае потребовалось бы теплокровному же аргентинозавру?..
(Г) плюс дополнительные причины типа проблем с живорождением (у млекопитающих чем крупнее мать, тем крупнее будет новорожденный детеныш, и в какой-то момент организм матери становится неспособен обеспечивать ресурсами слишком большой эмбрион) и проблем с перегревом (теплокровному млекопитающему нужно эффективно избавляться от «лишнего» тепла, тогда как у динозавров с этим хлопот было меньше).
Так что ответ, по сути, простой: слоны не вырастают как зауроподы потому, что они - слоны, а не зауроподы. :)-
Что-то мне кажется, что при огромной туше задача не нагреться, а охладиться. Я думаю, что у тех же травоядных динозавров в животе была та ещё печка (на днях УП рассказывал, что по изотопам кислорода это видно).
В шутку можно сравнить с нашим Солнцем, где в самой топке термоядерных реакций - удельное энерговыделение соответствует энерговыделению в тканях спящей игуаны. Но за счёт огромного объёма разогревается до десятков миллионов градусов.
-
-
ничего не получалось,
Это самое правдоподобное объяснение почему птицы "за 20" не летают, а птерозавры, вроде как да. Параметры атмосферы - довольно-таки "безумная идея", гораздо реальнее предположить, что они не летали на самом деле. По воздуху по крайней мере.-
Да понятно почему: крупные птицы взлететь не могут, а птерозавры могут.
Птицы взлетают за счёт лях, им нужно оттолкнуться и подпрыгнуть. А потом ляхи превращаются в мёртвый груз и их нужно таскать. Птерозавры подпрыгивали и махали 4 конечностями, поэтому таких ограничений не было.-
махали 4 конечностями, поэтому таких ограничений не было
Это может увеличить максимальный вес максимум вдвое (4 конечности работают вместо двух), но не в десятеро. Только "пламенный мотор", но ниоткуда не следует, что у птиц удельная мощность ухудшилась, тем более так сильно. -
1. Тут нельзя мыслить линейными категориями.
2. У птиц - ноги это мёртвый груз, у птерозавров - нет, они так же работали в полёте.
3. Появление более эффективных хищников в ходе эволюции создаёт давление сверху на гигантов. Примерно, как эпоху гигантских насекомых закончили те же самые птицы. Насекомые могут быть большими физиологически, но это не выгодно. -
-
Звери же запросто используют всякие инструменты!
Я - прям щас вот - торжественно позволяю птерозаврам пользоваться длинным балансирным шестом с оперением!
Что уж тут проще?
Ещё птенцом пусть каждый птерозавр определяет, что летать не выходит без шеста.
После чего легко находит решение: отламывает от древнго хвоща подходящей длины кусок и использует для полёта.
(уж как он его пристроит - пусть решает сам:
держит ли в зубах, или верхом на манер ведьмы на метле сидит,
или задними лапами держит, или вставляет куда...
А Эволюция завсегда поддержит: кто догадался как хвощ применять - выжил и детей научил. У кого фантазии не хватило, или недостаточно прочно хвощ крепил - безжалостно вычеркнут был из победителей, и за малочисленностью не оставил о себе для нас памяти в виде скелетов.
-
Ат и идея! Почему мы должны лишать их такой приспособы?
Строение тела птерозавров аэродинамически было ближе к коротконогим водоплавающим птицам, например, гусям и уткам: хвост у водоплавающих довольно короткий и крылья в полете расположены ближе к хвосту, чем к голове - почти, как у птерозавров.
Звери же запросто используют всякие инструменты!
Я - прям щас вот - торжественно позволяю птерозаврам пользоваться длинным балансирным шестом с оперением!
Из этого следует, что и те, и другие использовали длинную шею с головой в качестве активного противовеса, в частности, при взлете. И им не нужен (был) сзади никакой "длинный шест с оперением".
При взлете птерозавры, по-видимому, не подпрыгивали, а делали нырок-кивок головой, и, одновременно со взмахом крыльев, отталкивались от земли ногами. В отличие от них крупные птицы с длинными ногами подпрыгивают. Но у них и "шест с оперением" имеется в наличии.
При этом, гуси-лебеди тоже относятся к крупным птицам! Значит отсутствие сзади "длинного шеста с оперением" для них не создает проблем.
Так что его наличие-отсутствие, по-видимому, зависит от механики взлета/полета, а не от размеров летуна. Ну и, конечно же, от пропорций тела и расположения его центра тяжести.
Всю дорогу стремлюсь ни одного неуважения к собеседникам не высказать.
Всяко к логике оперирую.
И некомпетентность не скрывал свою ни разу, тем более по троллевскому принципу "сам дурак". (иначе говоря: ярлыками не пользуюсь. Честно скажу: считаю это недурным принципом. А Вы как к ярлыкам относитесь? Можете не отвечать, это риторика)
И, в целом, надеялся на взаимность.
-
Длинная шея у гусей и лебедей. Гуси травоядные птицы, щиплют траву, длинная шея позволяет легко дотянуться до земли и увеличивает радиус пастьбы с одной точки. Лебеди и гуси - птицы водоплавающие, едят водную растительность и мелкую водную живность, которых они достают опрокидываясь в воду головой вниз (они не плавают под водой), длинная шея позволяет дотянуться до еды поглубже и дает широкий радиус овата округи с одной точки.
Длинный хвост - руль, который нужен для точного управления полетом (хищные, насекомоядные, лесные птицы), гусям и лебедям не требуется виртуозное планирование, от того и хвост скромный.
Гуси и лебеди - тяжелые, жирные, с огромными желудками птицы - не попрыгаешь. Крупные хищные птицы - орлы, журавли - поджарые, легкие.-
Гуси и лебеди - тяжелые, жирные, с огромными желудками птицы - не попрыгаешь.
Горные гуси одни из самых высоко летающих птиц. Зафиксирован случай полёта горных гусей на высоте 10175 м, во время их перелёта из Средней Азии над Гималаями.
Лебеди тоже не промах! Был зафиксирован факт, когда они летели на высоте 8200 метров. Средняя высота полета лебедей 2500 метров.-
Ааааафигеть!!
Десять тысяч!!!
Это они, гуси, на самолетных эшелонах крылышками трясут???
Это надо запретить!
Во-первых, посбивают там пассажирские самолёты нафихх!
Во-вторых, там же холодно жутко! Если верить внутрисамолетным дикторам - так все минус пятьдесят!!! И кислорода с гулькин нос кот наплакал! Как там летать? О гусином здоровье кто будет беспокоиться?
Предлагаю незамедлительно:
Занести эшелон в красную книгу и отстреливать всякого гуся, забравшегося в незаконные высоты!
Или хоть сажать их на ближайший аэродром перехватчиками.
И прочими истребителяии с бомбардировщиками
-
-
Другой, совершенно другой, не современной, была растительность. Цветковые появились только под конец эпохи динозавров (дав шанс на лидерство млекопитающим, но это совсем другая история). Прочие виды растений, даже хвойные, намного хуже (в десятки раз!) умеют прокачивать воду от корней наверх.
Потому обильная растительность была только в сильно обводнённых местах. Таких мест было немало, потому что без растительности сильна эрозия, это образует обширные мелкие береговые зоны рек и морей. Но они всё же были ограничены в размерах. Сейчас такое называют болотами, но болота образуются с участием опять же цветковых. Так что "большие мелкие лужи" будет точнее.
И вот там было изобилие жизни. Причём в виде деревьев. Травы просто не было, мелкая поросль побыстрее прорывалась наверх и обзаводилась запредельным количеством лигнина для защиты от насекомых.
Вот такие острова. Площадью иногда в десятки тысяч квадратных километров, но острова. Между ними пустыня. Не пустыня-пекло, как мы сейчас видим, а просто сухие места, где в результате ничего не растёт.
Немалым подтверждением такой картины являются месторождения угля. Лигнин никто не ел (и сейчас не ест, за одним исключением), упавшие стволы уходили вниз сотнями и тысячами слоёв, образовав в конце-концов уголь. И вот этот уголь не покрывает всё сплошным слоем, а встречается именно островами-месторождениями.
Потом-то появилось то самое "исключение" - белая плесень, мукор. Которая лигнин ест (а потом уже получившееся доедают все остальные). И каменноугольный период кончился. А там и цветковые появились, которые сумели освоить бывшие пустыни. И трава, защищающаяся от насекомых не лигнином, а ядами-алкалоидами (никотин, морфин, кокаин, каннабиоиды, кофеин, теин и всё прочее, что мы так любим)
Именно этим объясняется гигантомания. В острове очень выгодно обезопасить себя размером. Это сейчас в степях такой слонопотам соберёт вокруг себя такие стаи хищников, что всё равно закусают. Тогда нет, неоткуда было им взяться.
Получается, описанная в статье четырёхногость отлично сочетается с идеей об изменении образа жизни с возрастом. Юная мелочь, весом не более пуда (это и для птиц граница полёта), летает и тем самым может расселяться по "островам". Выжив и отъевшись, растёт вплоть до гигантских размеров, перемещаясь уже на четырёх, пешком.
Есть и второй вариант расселения - если стать достаточно крупным, можно иметь достаточно ресурсов для перехода пустыни в следующий оазис (как это делают сейчас слоны). Но это уже про исходно не летающих.
-
-
это я ещё забыл добавить, что хвощам и папоротникам вода необходима и для размножения. Хвойным - нет, но в то время хвойные, хотя уже и распространились, не были основой этих лесов (что, опять же, показывают следы растений в угле). Так что наличие подпочвенной воды леса не создавало.
-
-
Да, пожалуй, Вы правы. Мхи, лишайники, хвощи - как раз иллюстрация того, что там, где слишком много воды, шанс лидировать имеют не-цветковые.
-
Спасибо, версия довольно интересная и заставляет подумать.
Из того, что сразу смущает: смешение эпохи динозавров и каменноугольного периода, которые порядочно разнесены во времени. Понятно, что иллюстрация, но тут есть проблема: когда растения отмирают и неразлагаясь лежат - уровень поверхности быстро поднимается и доступ к воде снижается.
Так же, в условиях "островной" системы вряд ли могут появиться гигантские хищники, так как в пределах островов им просто не хватит питания. Они должны быть адаптированы для длительных переходов в неблагоприятных условиях к другим островам, но вроде таких адаптаций у них нет.
С другой стороны, может быть так, что травоядные гиганты с длинными шеями - как раз адаптированы к таким переходам? :) Могут идти по прямой, и в то же время подъедать всю растительность на пути.-
смешение эпохи динозавров и каменноугольного периода, которые порядочно разнесены во времени
есть расхождение, конечно :-) Но уголь - просто для иллюстрации того, что покрытие не было сплошным. Каменноугольный закончился, когда появился мукор, а цветковые начали распространяться уже под конец динозавровых времён. В этом промежутке растения были в принципе те же (хотя хвойных становилось постепенно больше).когда растения отмирают и неразлагаясь лежат - уровень поверхности быстро поднимается и доступ к воде снижается.
в маленьком пруду - да. Если бы доступ к воде снижался - отмершие стволы оставались бы на поверхности и в конце-концов сгорали бы.
Так что куда-то они все уходили, иначе не было бы слоёв каменного угля в десятки метров толщины.Так же, в условиях "островной" системы вряд ли могут появиться гигантские хищники, так как в пределах островов им просто не хватит питания
Как пример возможного размера оазиса: площадь кузбасского угольного месторождения - 27тыс км².Они должны быть адаптированы для длительных переходов в неблагоприятных условиях к другим островам, но вроде таких адаптаций у них нет.
А какие адаптации у слонов, кроме размера?
https://elementy.ru/kartinka_dnya/1868/Pustynnye_slony
По-моему, это показательный пример.Могут идти по прямой, и в то же время подъедать всю растительность на пути.
а вот с этим проблема :-) пустыня же. И травы нет пока и в помине.-
Читать, читать и еще раз читать
https://repository.geologyscience.ru/bitstream/handle/123456789/38880/Mcal_04.pdf?sequence=1&isAllowed=y
https://sci-hub.ru/https://doi.org/10.1016/S0016-7878(59)80068-7
https://www.researchgate.net/publication/46256368_Broad-Scale_Patterns_of_Late_Jurassic_Dinosaur_Paleoecology
https://d1wqtxts1xzle7.cloudfront.net/36751407/Hinz_et_al_-_2010_-_Palaeobio_Palaeoenv_-_A_high-resolution_three-dimensi onal_reconstruction_of_a_fossil_forest-libre.pdf?1424773869= &response-content-disposition=inline%3B+filename%3DA_hig h_resolution_three_dimensional_reco.pdf&Expires=17477324 07&Signature=B4y2aMT2fvEtgICHVjXA81FfZgWgiLuInSjsBZrum2a zfm1lAF-1LO96wgsg5bj727km2c4M1FSYWAM5H62fjK9bSc9ODEHAyPY76cM vmBOY-o3Zf81fCRi03OyJ0FUKuWnOw-GybYYO8H-tXE2VxcyQ2VA9av7-5ZK KYk7VcjXHWQi~QS9LSKRPukdDyU15Sp7XL1Fe-8bjMKm7~dzrMflaQcva1Pw QUY~rHgdUx-V6-y2X7sJ3xrYRgdgahpssH1mEsdPLgBC3KXcDNY4R8Sbn3dl 07MJDodjZJe8prO~USJRNwIfjrLYTAtKVsYEm6syFyQV--2F7bKYImCd-BQ_ _&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA
https://www.researchgate.net/publication/322766661_Panalesis_Towards_global_synthetic_palaeogeographies_using_integratio n_and_coupling_of_manifold_models
https://sci-hub.ru/https://www.sciencedirect.com/science/article/abs/pii/S0895981121001711
-
-
-
-
-
-
-
Именно этим объясняется гигантомания. В острове очень выгодно обезопасить себя размером.
человек просто бредит. Беда... )
https://elementy.ru/novosti_nauki/433805/Ostrovnye_karliki_i_giganty_rezultat_deystviya_kompleksa_ekologicheskikh_i_evol yutsionnykh_faktorov Европейские экологи СОБРАЛИ ВСЕ ИМЕЮЩИЕСЯ ДАННЫЕ по размерам тела островных видов позвоночных и их материковых родичей. На этом обширном материале они подтвердили реальность островного эффекта: КРУПНЫЕ ЖИВОТНЫЕ НА ОСТРОВАХ МЕЛЬЧАЮТ, А МЕЛКИЕ УКРУПНЯЮТСЯ. Составив многофакторные корреляции, ученые объяснили формирование островных карликов и гигантов действием целого комплекса причин. На размер тела влияет множество факторов, так что механизм островного эффекта и должен включать множество компонентов.
По-видимому, это происходит по причине отсутствия на островах большого количества крупных хищников, либо их там нету, вообще.
Правило Фостера (или островной эффект) — ХОРОШО ИЗВЕСТНАЯ ЭВОЛЮЦИОННАЯ ЗАКОНОМЕРНОСТЬ, согласно которой размер обитателей островов имеет тенденцию изменяться по сравнению с их материковыми прародителями. Некрупные виды увеличиваются (это так называемый островной гигантизм), а крупные, наоборот, уменьшаются (это островная карликовость). Иными словами, РАЗМЕРЫ ЖИВОТНЫХ СТРЕМЯТСЯ К УСРЕДНЕНИЮ.
При их наличии, удобнее быть мелким и незаметным и прятаться, в частности, в норах, либо быть очень крупным и сильным.
Средними быть удобнее только, если вокруг нету крупных хищников, которые могут запросто поймать и сожрать; на острове им убежать (и, вообще, избегать встреч с хищниками) сложнее. Среднего размера животные для них предпочтительная добыча: мяса много, справиться легко, доставать из нор не надо.
А почему на островах нету крупных хищников? Можно было бы и не искать ответа на этот вопрос. Но я, всё же, попытаюсь. Судя по всему, крупные хищники, даже, если они там есть, быстро переловят всю живность (возможно, кроме слонов) и помрут/вымрут сами. А, случайно выжившая/(хорошо спрятавшаяся) мелкая живность, в отсутствии крупных хищников (и, возможно, съеденных ими более мелких), быстро расплодится и вырастет в размерах. При этом, становиться/быть слишком крупным в отсутствии крупных хищников и при ограниченном питании нет необходимости/не выгодно.
Резюмирую: на многие вопросы не надо искать слишком сложные ответы; ответ на них может быть довольно простым и лежать на поверхности.-
человек просто бредит. Беда... )
Спасибо, Вы хорошо определили свои способности поддерживать уровень дискуссии.а крупные, наоборот, уменьшаются (это островная карликовость)
вопрос размера острова/оазиса. И да, наличия хищников. В отличие от морских островов, где обмен особями между островами практически невозможен, миграция между оазисами по суше была возможна, хотя и не для всех. Пример - птицы и слоны, вполне мигрируют через пустыни. -
-
Пример - птицы и слоны, вполне мигрируют через пустыни.
Мигрировать через пустыни пешком будучи очень крупным не очень-то просто - это большой расход энергии, воды.
Ответ ИИ:Вес саванного слона колеблется от 5 тонн у самок и до 7 тонн у самцов, при росте от 3 до 3,8 метра. Длина тела достигает 6–7,5 м, высота в плечах — 3–3,8 м.
Как я и предполагал.
Пустынные слоны, в свою очередь, отличаются несколько меньшим размером тела, более длинными ногами и широкими подошвами. Их физические способности позволяют пройти до 70 км в день в поисках воды по невыносимой жаре.
Таким образом, САВАННЫЙ СЛОН значительно КРУПНЕЕ ПУСТЫННОГО.-
Как у вас красиво "несколько меньший размер" превратился в "значительно крупнее" :-)
На самом деле пустынные слоны - иллюстрация того, что размер позволяет пересекать пустынные пространства, но несколько избыточная. Они не только пересекают пустыню, но и живут там. Находясь же в более благоприятных условиях они, как всего лишь экологически отличающиеся, наберут тот же вес.-
Это всё ответ ИИ
Я от себя ничего не добавлял.
PS. Интересно, и как это противоречит тому, чтоМигрировать через пустыни пешком будучи очень крупным не очень-то просто - это большой расход энергии, воды.
?-
Помнится, абсолютным доказательством считалось "так в газете написано". Потом "по телевизору сказали". Потом "это же из интернета". Теперь для вас доказательно "это выдал ИИ" :-D
-
ИИ же берет информацию откуда-то. Кому понадобилось её искажать?
"Bedal так сказал" для меня точно не является доказательством.-
Нейросети похожи на людей в том числе тем, что легко галлюцинируют.
Не может быть источника, опровергающего "значительно крупнее", а вот источник, это утверждающий, хотелось бы увидеть. Потому как большая часть ответа цитирует вики, но вот такого вывода там и близко нет. Зато есть указание, что пустынные слоны - тот же вид, что и саванные, отличия только экологические. Как люди во времена голода становятся ниже ростом, еды не хватает вырасти.-
Еще ИИ эволюционируют, причем экспоненциально.
PS.Новый ИИ-метеоролог Microsoft превзошел ведущие мировые центры прогнозирования 22.05.2025
Корпорация Microsoft разработала искусственный интеллект Aurora, способный предсказывать погоду, качество воздуха и траектории тропических штормов точнее и быстрее традиционных методов. Результаты исследования, опубликованные в журнале Nature, показали, что новая система генерирует 10-дневные прогнозы с меньшими затратами и опережает ведущие мировые метеорологические центры. Aurora, пока не коммерциализированная, успешно спрогнозировала все ураганы 2023 года, включая разрушительный тайфун Доксури, который обрушился на Филиппины. В то время как официальные прогнозы предполагали его движение севернее Тайваня, ИИ точно определил место и время удара за четыре дня до события.
Источник: New-Science.ru https://new-science.ru/novyj-ii-meteorolog-microsoft-prevzoshel-vedushhie-mirovye-centry-prognozirovaniya/ -
Да, и уровень галлюцинаций при этом вырос. Всё как у людей...
Не пытайтесь меня удивить достижениями ИИ. Я, грешным делом, писал нейросеть ещё тогда, когда и слова такого не было. А ИИ называлось нечто, лежащее в основе ПРОЛОГа. Обратная раскрутка логики. На поиграться - забавная идея, но провальная.-
-
предъявить источник информации о том, что пустынные слоны ЗНАЧИТЕЛЬНО меньше саванных можете? Нет? Не морочьте голову.
-
Спросите у ИИ )
PS. Если имеется тенденция к снижению размеров, то она может сохраниться - если не будет контактов между двумя видами.-
Вы юлите. Зачем? Между "несколько" и "значительно", да ещё и капсом, различия не понимаете? Юлите.
Пустынные слоны - пример того, что размер позволяет переходить из одного оазиса в другой без специальных режимов организма (как у верблюдов) и, значит, без потери возможности благоденствовать в оазисе (тот же верблюд в оазисе другим животным быстро проигрывает).
Итого, как уже указывали здесь, Вы просто троллите, не отдавая себе труда что-то узнавать и думать. Но троллей кормить нельзя - и я больше с Вами переписываться не буду.
-
-
-
-
-
-
-
-
-
-
-
-
Например, этого - https://avatars.dzeninfra.ru/get-zen_doc/96748/pub_5c47a9ad9
и этого - https://img.mix.com.tj/img/b/7/22/53428.jpg
Если это реконструкции (куклы), то выглядят они очень реалистично.
Особенно удивляет строение крыла птерозавра на первом фото, в нем отчетливо видно, что крыло изламывалось назад для ходьбы с опорой на 4 конечности. Была ли известна такая их особенность в то время, когда была сделана эта фотография?
-
Фейк, причем современный: https://karlshuker.blogspot.com/2021/01/elucidating-two-civi
l-war-pterodactyl.html?m=1
Фотографии состарены, а в самих "птерозаврах" куча ошибок. Например, на последнем фото у "птерозавра" крылья летучей мыши. Также у настоящих птерозавров были закругленные кончики крыльев.-
Про второе(в статье) фото знаю, что это фейк - https://avatars.dzeninfra.ru/get-zen_doc/1582174/pub_5ec01cd
bbe2dce5b920f2f42_5ec023ecfce1527a111c4a02/scale_1200
Где-то в интернете сегодня эту модель в руках современно одетых киношников видел.
PS. Из комментариев к статье:
"крылья были цифровым воспроизведением из эпизода сериала BBC "Прогулки с динозаврами"
Ну, тогда понятно почему они выглядят так реалистично )
Хотя кто-то в статье утверждает, что модель птерозавра цифровая, наличие тени под ним говорит о том, что это должна была быть кукла.
А, вот, у второго фото отсутствие тени от головы говорит о том, что это может быть фотомонтаж.
PPS. Интересен вызвал последний комментарий к статье:
"Мы с женой сидели на заднем крыльце. Был прекрасный летний день, голубое небо с небольшими пушистыми облаками, разбросанными повсюду. Когда мы смотрели на облака, мы заметили вдалеке небольшой самолет, летящий между облаками. Мы думали, что это самолет,
пока он не захлопал крыльями. Мы оба не могли себе представить, какая птица может вырасти до размеров частного самолета." -
Например, на последнем фото у "птерозавра" крылья летучей мыши.
Если предположить существование двух видов птерозавров - сухопутных и лазающих по деревьям, то у птерозавра-древолаза крылья, теоретически, могли бы быть похожи на крылья летучей мыши, потому что ему складывать их, выворачивая наружу, не надо было бы. А, вот, сухопутному, ходящему на четвереньках, крылья надо было выворачивать наружу, и тут (3 или 4?) лишних пальца мешались бы - складки перепонки болтались бы, как у сложенного зонта. Когда назад загибается только один палец (мизинец), несущий все крыло, то это получается более компактно - перепонка на пальце все так же натянута, ничего не болтается - и не мешает ходить на четвереньках.
Судя по всему, именно этим объясняется разное строение крыльев у [большинства] птерозавров и летучих мышей - птерозавры с выворачиваемыми назад крыльями были сухопутными, в первую очередь, самые крупные из них.-
Проблема в том, что не найдено ни одного птерозавра с анатомией крыла, отличающейся от остальных представителей отряда. Собственно, это как если бы ученые обнаружили оленя с лапами, а не копытами. Так что на фотографии всего лишь увеличенный до безобразия крылан с головой птерозавра. Современный новодел.
-
Понятно )
Зато когда-то существовал пакистанский "олень", вернее кит, вернее предок китов, с копытцами на пальцах лап ;)
PS. Уточнил: как утверждает предполагаемый автор того фото, на нем изображены крылья фруктовой летучей мыши )
Крылан, похоже, был пририсован здесь - https://avatars.mds.yandex.net/i?id=776c7fd86e6f6627b21c7d253778fbb5-5602191-images-thumbs&n=13
-
-
-






Рис. 1. Поздние птерозавры, такие как арамбургиания (Arambourgiania), вполне могли быть полуназемными хищниками. Рисунок из статьи D. Naish, M. Witton, 2017. Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators