Асинхронность цветения и активности опылителей влияет на направление отбора

Даже в пределах одного сезона давление отбора на растения может меняться. Одним из примеров такой ситуации служит фенологическая асинхронность между цветением и активностью опылителей: одни растения цветут рано и насекомых на них почти нет, а другие зацветают позже, когда опылителей много. Такие временные рассогласования могут менять связь между признаком растения и его приспособленностью, то есть влиять на то, какие растения оказываются более успешными в размножении. Недавнее исследование, опубликованное в журнале Annals of Botany, показало, как это работает на примере Stigmaphyllon paralias — растения, выделяющего масло для специализированных пчел. Оказалось, что растения, цветущие в пик сезона, при нехватке визитов пчел (сильное лимитирование пыльцой) получают преимущество за счет крупных цветков — у них больше семян и больше визитов. Поздноцветущие растения в условиях достаточного количества опылителей, напротив, теряют это преимущество: крупные цветки требуют много ресурсов, а семян образуется меньше. В итоге направление отбора менялось с течением времени: в начале сезона выгоднее иметь большие цветки, в конце — мелкие.
Давление естественного отбора не является постоянным: оно изменяется во времени и пространстве, определяя направление эволюционных преобразований. Согласно концепции географической мозаики коэволюции (The Geographic Mosaic of Coevolution), разные популяции одного и того же вида могут вступать в коэволюционные отношения с партнерами по-разному, поскольку характер взаимодействий варьирует в зависимости от географического контекста. В результате эволюция носит локальный характер: в одних популяциях отбор оказывается особенно сильным и направленным («горячие точки»), тогда как в других популяциях он ослаблен или вовсе отсутствует («холодные точки»; см. Неядовитые змеи вырабатывают устойчивость к смертоносному яду тритонов, «Элементы», 21.03.2008). При этом постоянный поток генов между популяциями объединяет эти локальные сценарии, и коэволюция предстает не как единый процесс, а как мозаика пространственно разнородных взаимодействий.
Не менее изменчиво и селективное давление во времени. Разные агенты отбора могут влиять на популяцию с разной силой в разные годы или даже в разные месяцы, ускоряя или замедляя эволюционные процессы. Колебания возможны даже в пределах одного репродуктивного сезона. Особую роль здесь играют биотические взаимодействия, например, партнерство между растениями и их опылителями. Если пики цветения растений и активности опылителей не совпадают, возникает временная асинхронность. Она может быть вызвана конкуренцией за опылителей между разными видами растений или смещением сроков цветения — в том числе под влиянием изменения климата. В результате одни растения в популяции цветут тогда, когда насекомых мало, а другие — когда их много. Такое рассогласование влияет не только на число завязавшихся семян, но и на то, какие признаки оказываются выгодными именно в этот момент.
Недавнее исследование, опубликованное в журнале Annals of Botany, было посвящено тому, как совпадение или рассогласование во времени между цветением растения и активностью опылителей влияет на лимитирование пыльцой и, в конечном счете, на связь между признаками цветка и репродуктивным успехом.
Исследование проводилось в Национальном парке Катимбау на северо-востоке Бразилии в 2020 году, в разгар цветения местного растения Stigmaphyllon paralias. Этот кустарник или полукустарник из семейства Мальпигиевые цветет в период с октября/ноября по май/июнь, а пик его массового цветения обычно приходится на декабрь–март. У каждого отдельного растения «праздник цветения» длится около месяца, а отдельный цветок живет всего до трех дней. Цветки S. paralias — желтые, зигоморфные (то есть асимметричные), с пятью лепестками (рис. 1). Лепесток состоит из двух частей — верхней широкой, называемой отгибом, и нижней узкой, похожей на черешок листа и называемой ноготком. У одного из пяти лепестков — «флагового» — этот ноготок толще, чем у остальных. Чашелистики цветка снабжены особыми масляными железками (элайофорами, см. Elaiophore). Ради этого масла цветок и посещают его главные опылители — пчелы из группы Centridini (семейство Apidae). Поведение этих пчел стереотипно: они крепко захватывают флаговый лепесток челюстями, оставляя на нем заметный след — характерное некротическое повреждение. Этот след невозможно спутать с чем-то другим, поэтому он надежно показывает факт визита. Затем они соскабливают масло с железок специальными щеточками на передних и средних ногах (рис. 1, С, D). Одновременно на их теле оседает пыльца, которая переносится к другим растениям. Без таких пчел кустарник размножиться не может: он самонесовместим, то есть собственная пыльца не приводит к образованию семян. Плоды S. paralias содержат до трех семян.
Ученые хотели понять, как совпадение или рассогласование во времени цветения растения и активности пчел влияет на его размножение. Для этого они организовали два «раунда» наблюдений продолжительностью приблизительно по 10 дней: первый — в период массового цветения, когда растений с цветками было много, но пчелы почти не прилетали; второй — через 3–4 недели, когда цветущих растений осталось меньше, зато активность опылителей резко выросла. При этом число раскрытых цветков на растение между раундами не различалось. В каждом раунде исследователи работали с 90 кустами (всего 180), расположенными не ближе трех метров друг от друга, чтобы избежать отбора клонов.
Чтобы разобраться, как особенности цветков Stigmaphyllon paralias связаны с их успешным размножением, исследователи внимательно измеряли и сравнивали несколько параметров. Их интересовали три вещи: цветочный фенотип (то есть внешние характеристики цветка), лимитирование пыльцой (насколько недостаток пыльцы мешает образованию семян) и частота посещений опылителями. Для этого на каждом кусте помечали от одного до трех свежераспустившихся цветков. Главным показателем фенотипа авторы выбрали размер цветка, так как у многих растений именно крупные цветки лучше привлекают опылителей. Кроме того, у S. paralias размер лепестков положительно связан с размером масляных железок, ради которых и прилетают специализированные пчелы. Чтобы получить точные данные, исследователи фотографировали цветки с масштабной линейкой и измеряли расстояние между крайними точками боковых лепестков.
Далее они проверяли, насколько растениям хватает пыльцы для нормального семенного воспроизводства. Для этого использовали классический метод — сравнение естественного опыления и искусственного (ручного) опыления. В каждой серии наблюдений половину кустов оставляли на волю природы (естественное опыление), а другую половину люди брали под свой контроль (ручное опыление). В последнем случае в первый день раскрытия цветка на его рыльце наносили свежую пыльцу, собранную с другого растения, расположенного не ближе трех метров. Такой выбор исключал возможность, что донор и реципиент окажутся клонами или близкими родственниками. Для опыления использовали пыльники только что раскрывшихся цветков: их аккуратно извлекали и втирали в рыльце.
Через 15 дней ученые проверяли, образовались ли семена. Сравнивая результаты между двумя группами, можно было понять, насколько в естественных условиях растения страдают от нехватки перекрестного опыления. Этот показатель называется «лимитирование пыльцой» и выражается формулой: 1 − (среднее число семян при естественном опылении)/(среднее число семян при ручном опылении). Значение, близкое к 1, означает сильный дефицит пыльцы (естественное опыление работает плохо), а значение, близкое к 0, говорит о том, что пыльцы хватает.
На второй день жизни каждого помеченного цветка ученые фиксировали, есть ли на флаговом лепестке некротическое повреждение ткани, оставленное челюстями пчелы. Так получали простую бинарную характеристику: цветок либо хотя бы раз посетили маслособирающие пчелы, либо он остался незамеченным.
Кроме этого, исследователи подсчитывали, сколько плодов образовалось на каждом растении, — это служило обобщенным показателем его приспособленности. Исследователи проверяли, как размер цветка связан с успехом размножения. В качестве показателя успеха они использовали относительную приспособленность. Для каждого растения ее считали как произведение числа плодов на долю семяпочек, превратившихся в семена. Этот показатель нормировали, то есть сравнивали со средним значением по всей популяции, чтобы было удобно видеть, кто успешнее, а кто хуже среднего. В анализ включили только те растения, которые опылялись естественным образом, без вмешательства человека (87 особей).
В результате выяснилось, что отбор по размеру цветков зависит от времени цветения. У растений, цветущих в разгар сезона, больше шансов на успех имели крупные цветки. А у тех растений, что зацветали позже, наоборот, преимущество оказалось у мелких цветков. Иными словами, направление отбора меняется в зависимости от времени цветения (рис. 2). Более того, сила отбора тоже была разной: в пик сезона давление отбора на размер цветка было заметно сильнее, чем в конце. Если же рассматривать всю популяцию целиком, без учета времени цветения, то статистически значимого отбора на размер цветка не выявлялось.

Рис. 2. Влияние стандартизированного размера цветка (Zflower size, по горизонтальной оси) на относительную приспособленность (W, по вертикальной оси) у растения Stigmaphyllon paralias в разные периоды цветения. Наблюдается сильный корреляционный отбор: направление отбора на размер цветка (Z) положительно для особей пикового (Peak, желтым) и отрицательно для особей позднего (Late, фиолетовым) периода цветения. Рисунок из обсуждаемой статьи
Лимитирование пыльцой тоже было изменчиво во времени. В пик цветения нехватка пыльцы была очень высокой (индекс лимитирования пыльцой — 0,74). Растения, которые получили дополнительное ручное опыление, завязывали семян в три раза больше, чем те, что остались на естественном опылении. Зато у поздноцветущих растений лимитирования пыльцой почти не было (индекс лимитирования пыльцой — 0,03). Их завязывание семян почти не зависело от «помощи» человека и было сопоставимо с результатами ручного опыления в пике цветения (рис. 3).
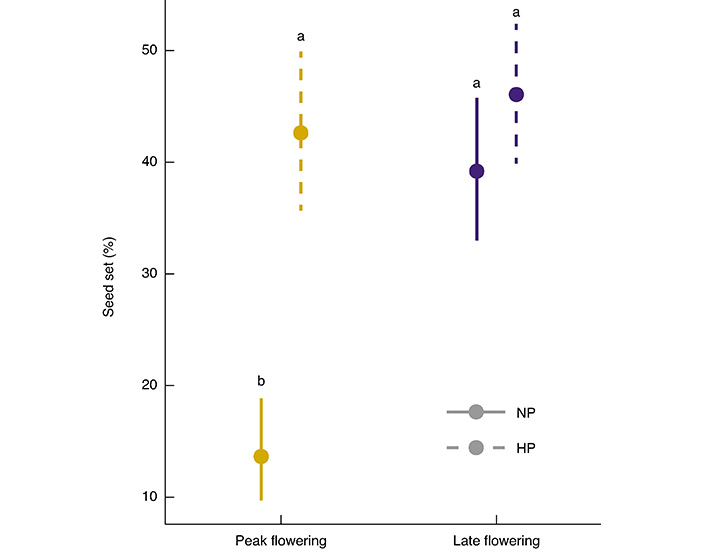
Рис. 3. Процент завязывания семян (seed set, по вертикальной оси) у особей пикового (peak flowering) и позднего (late flowering) периодов цветения при естественном опылении (NP) и дополнительном ручном опылении (HP) как оценка уровня лимитирования пыльцой в популяции Stigmaphyllon paralias. Разные буквы обозначают статистически значимые различия в завязывании семян. Рисунок из обсуждаемой статьи
Во время наблюдений были зарегистрированы три вида маслособирающих пчел: Centris (Centris) aenea, C. (C.) caxiensis, C. (Trachina) perforator. Частота визитов опылителей резко различалась между периодами наблюдения (рис. 4). У растений, цветущих в пик, следы визитов пчtл (некротические отметины на флаговых лепестках) были лишь на 7,5% цветков (из 134). А у поздноцветущих растений — уже на 93,6% цветков (из 140). То есть разница была огромной.
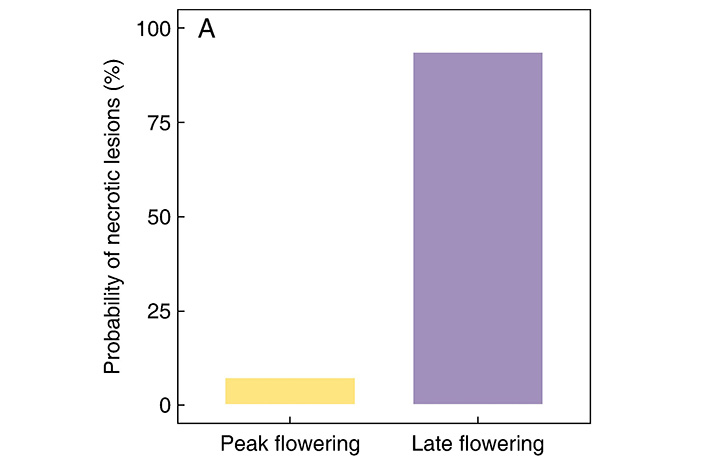
Рис. 4. Различия в доле некротических повреждений на флаговом лепестке в качестве индикатора посещений (по вертикальной оси) между растениями Stigmaphyllon paralias пикового (peak flowering) и позднего (late flowering) периодов цветения. Рисунок из обсуждаемой статьи
Кроме того, оказалось, что пчелы чаще выбирали более крупные цветки. То есть размер напрямую влиял на вероятность визита, и чем больше был цветок, тем выше шанс, что его заметят. Более крупные цветки играют роль честного сигнала — они обещают опылителю больше нектара, масел или смол, даже если те остаются невидимыми до посадки. В первый период наблюдения, когда опылителей было мало, крупные цветки действительно давали своим владельцам преимущество: они чаще привлекали пчёл и обеспечивали больше семян.
Однако при высокой активности опылителей различия между фенотипами теряли значение. Все растения получали достаточно визитов, лимитирование пыльцой исчезало, и возможности для отбора почти сводились на нет. Более того, в таких условиях ограничивающим фактором становились уже не визиты пчел, а ресурсы среды. В дефиците влаги или питательных веществ, характерном, например, для сухих лесов, даже обильное опыление не гарантирует высокий урожай семян. У растений с крупными и броскими цветками это приводит к обратному эффекту: значительная часть ресурсов уходит на рекламу, тогда как на формирование семян сил остается меньше. В результате крупный цветок превращается из преимущества в обузу, а связь между размером цветка и репродуктивный успехом может становиться отрицательной. Однако, возможно, крупные цветки сохраняют преимущество в другой плоскости — через мужскую функцию, обеспечивая лучшее качество пыльцы или ее более успешную конкуренцию, но такие аспекты пока трудно измерить.
Таким образом, в пределах одного сезона цветения давление отбора не остается стабильным. Оно может меняться от положительного к отрицательному в зависимости от того, совпадает ли активность опылителей с пиком цветения и насколько ресурсы ограничивают растения. Это ведет к тому, что опылители не всегда формируют устойчивый и предсказуемый отбор, а скорость эволюционных изменений в таких условиях может снижаться. В будущем важно понять, как продолжительность сезона цветения влияет на скорость таких изменений: чем короче период цветения, тем меньше вариаций в селективных давлениях, а значит, и эволюция может идти по иным траекториям.
Источник: Liedson Tavares Carneiro, Isabel Cristina Machado. Evolutionary consequences of flowering–pollinator asynchrony: the case of a floral oil-producing plant and its oil-collecting bees // Annals of Botany. 2025. DOI: 10.1093/aob/mcaf126.
Елена Устинова
Последние новости













Рис. 1. Цветки Stigmaphyllon paralias (A, B) и их взаимодействие с маслособирающими пчелами (C, D). A — вид спереди; fs — измерение размера цветка (красная пунктирная линия). B — вид сбоку; stg — листовидные рыльца; stm — тычинки (показаны три из десяти). Красная стрелка указывает на некротическое повреждение на флаговом лепестке, оставленное хватательным поведением маслособирающих пчел. C — пчела Centris caxiensis; красные стрелки показывают, как пчела захватывает флаговый лепесток верхними челюстями и использует передние и средние ноги для сбора масла. D — вид сзади; детальное изображение сбора масла с элайофоров (ela) особью C. caxiensis. Длина масштабных отрезков — 3 мм. Рисунок из обсуждаемой статьи