Камень, ножницы, бумага, ящерица, Спок

Задача
В одной популяции животных нередко одновременно присутствует несколько разных, четко различающихся по внешним признакам, морф самцов. За счет чего может возникать такая ситуация и как она может поддерживаться?
Подсказка 1
И «возникновение такой ситуации», и то, «как она может поддерживаться», — разумеется, очень общие вопросы. Подумайте, какие варианты ответов подразумеваются.
Подсказка 2
Не сказано, сколько морф самок есть в популяции; подумайте, как полиморфизм самцов может быть связан с полиморфизмом самок.
Подсказка 3
См. название задачи — в нем содержится один из вариантов ответа.
Решение
Сначала уточним условие. Оно содержит несколько вопросов и подразумевает несколько ответов. Один вопрос — какие гены отвечают за различия морф и как наследуются эти различия? Почему в результате скрещиваний морфы не исчезают? Другой вопрос — как появлению четких морф могут способствовать условия среды, эволюционная история вида и особенности его поведения? Еще один, более частный вопрос, — какие признаки разных морф позволяют им не быть вытесненными из популяции из-за преимущества одной, самой приспособленной морфы? (Поскольку речь в вопросе идет о самцах — вероятно, эти признаки должны быть как-то связаны с размножением, хотя и не обязательно).
Давайте сначала представим себе возможные ответы на вопрос, откуда могут взяться несколько разных морф в популяции — не важно, самцов или самок; ведь в условии не сказано, что полиморфизм есть только у самцов. Для примера дальше мы часто будем говорить об окраске, но та же логика применима и к другим признакам.
Начнем с того, что многие наследственные признаки, вообще-то, дискретны. Грегору Менделю, как известно, удалось подобрать семь пар таких моногенных «менделирующих» признаков у гороха — например, горошины у его сортов были либо желтые, либо зеленые, но не желто-зеленые. А если такие признаки оказываются нейтральными и не влияют ни на какие другие свойства организма, в достаточно большой популяции такой полиморфизм будет сохраняться бесконечно долго (см. Закон Харди — Вайнберга). Исходно новые нейтральные аллели могут повысить свою концентрацию из-за дрейфа генов — например, в период низкой численности или сильной подразделенности большой популяции на микропопуляции.
Если окраска (или другой признак морфы) определяется одним геном с двумя аллелями, то морф будет две при полном доминировании и три — при неполном. При трех аллелях и неполном доминировании морф будет уже шесть, и т. д. По очень многим аллелям равновесие Харди — Вайнберга наблюдается в реальных популяциях, и среди них встречаются, конечно же, и аллели окраски (и других внешних признаков). Кстати, если нейтральный признак сцеплен с полом, а его рецессивный аллель достаточно редок, может возникнуть впечатление, что полиморфизм есть только у гемизигот (самцов или самок). Например, красно-зеленая цветовая слепота (полная или частичная) в Западной Европе встречается, по данным Википедии, у одного из 12 мужчин и у одной из 200 женщин. Взяв выборку в 50 особей, ученый с высокой вероятностью обнаружил бы полиморфизм по такому признаку только у самцов!
Возможно, к нейтральным признакам относится диморфизм гениталий у богомолов рода Ciulfina. У большинства изученных видов в разных пропорциях встречаются самцы с зеркально симметричными «правыми» и «левыми» гениталиями. При этом гениталии самок симметричны, и не удалось выявить никаких преимуществ при спаривании у одной из морф самцов. Различия в хиральности гениталий не предотвращали и межвидовые скрещивания с тем видом, у которого все самцы были «правыми», несмотря на полную посткопулятивную изоляцию.
Еще один вариант (часто нарушающий соотношение Харди — Вайнберга) — миграции. Если смешиваются особи из нескольких разных популяций, где отбор действовал в разных направлениях,легко можно наблюдать четкие морфы. Так дело обстоит во многих популяциях человека — например, где-нибудь в Лондоне или в Москве рядом с Российским университетом дружбы народов. И хотя изменчивость расовых признаков на самом деле непрерывная (см. Послесловие), это не так легко обнаружить. Потомков представителей разных морф мы, скорее, будем поначалу воспринимать как еще одну, третью морфу. Если предположить, что миграционная активность выше у самцов (что часто так и есть), то полиморфизм мы можем в таких случаях наблюдать только у одного пола.
Сходная картина полиморфизма у любого пола может возникнуть и при межвидовой гибридизации, когда гибриды будут восприниматься как отдельная морфа или морфы (подробно этот случай описан в задаче Где какой цвет в моде). Тут полиморфизм только у самцов может быть связан с тем, что у близких видов самцы различаются по окраске, а самки — нет.
В некотором смысле противоположный гибридизации пример — наличие нескольких морф самок и самцов и ассортативное (избирательное) скрещивание между ними. Такая ситуация может говорить о начавшемся симпатрическом видообразовании. Мы наблюдаем нестабильную ситуацию: со временем дизруптивный отбор сделает свою работу, и популяция распадется на два разных вида. Хороший пример — несколько видов цихлид, живущих в озерах Никарагуа, Манагуа и близлежащих небольших кратерных озерах. В каждом озере независимо возникли «золотые» светлые морфы самцов и самок, обычно малочисленные по сравнению со «стандартными» темными морфами (см. H. Kusche et al., 2015. Sympatric ecological divergence associated with a color polymorphism). Утверждается, что окраска — моногенный признак и за него отвечает ген с двумя аллелями и полным доминированием (золотистая окраска — доминантный признак). Из-за высокой избирательности скрещивания («золотые» самки предпочитают «золотых» самцов, темные — темных) между морфами возникли сравнимые с видовыми генетические различия, а также различия в форме тела, питании и других признаках.
Интересный пример поддержания полиморфизма окраски, связанный с научением и половым отбором, описан в новости Половой отбор может привести к видообразованию, если сыновья помнят, как выглядела их мать, «Элементы», 07.10.2019. Оказалось, что дочери лягушек-древолазов запоминают, как выглядела воспитавшая их мать, и преимущественно выбирают половых партнеров такого же цвета, а сыновья агрессивнее гоняют самцов этого цвета — почти по Фрейду. Модели показывают, что в этой ситуации возможно симпатрическое видообразование без дизруптивного отбора — только за счет полового.
Возможно, влияние научения на поддержание полиморфизма встречается и у других видов. Например, влияние запоминания на половые предпочтения самцов предполагается для орла-карлика, где светлая и темная окраска морф, видимо, наследуется по Менделю. Судя по частотам морф, светлые самцы-гетерозиготы могут чаще выбирать гомозиготных темных самок, руководствуясь импринтингом (совсем по Фрейду). Такое избирательное скрещивание между разными морфами называется дисассортативным и тоже может быть связано с поддержанием полиморфизма, если действует балансирующий частотно-зависимый отбор (см. Послесловие).
Еще один вариант возникновения и поддержания полиморфизма — искусственный отбор. Домашних животных мы здесь затрагивать не будем. Но оказывается, что в природе «искусственный» отбор тоже может приводить к полиморфизму — это отбор полиморфных по окраске колоний тлей, который, вероятно, ведут муравьи (см. Муравьи помогают тлям сохранять разнообразие окраски, «Элементы», 19.09.2016). Может быть, когда-нибудь найдется и подобный пример полиморфизма только среди самцов?
Нередко диморфизм или полиморфизм не связан ни с каким генетическими различиями. Тогда он называется полифенизм (см. Polyphenism). Например, сезонный полифенизм широко распространен у насекомых (см. обзор A. M. Shapiro, 1976. Seasonal Polyphenism) и хорошо изучен у некоторых бабочек. В России наиболее известен пример Araschnia levana — пестрокрыльницы изменчивой, у которой летняя морфа крупнее и настолько сильно отличается по окраске, что была описана как отдельный вид. При коротком световом дне и низкой температуре гусеницы превращаются в зимующих куколок, из которых выходит весенняя морфа; при длинном световом дне без диапаузы развивается летняя морфа (см. A. Vilcinskas, H. Vogel, 2016. Seasonal phenotype-specific transcriptional reprogramming during metamorphosis in the European map butterfly Araschnia levana и ссылки в этой статье про молекулярные механизмы этого диморфизма, а также заметки про сезонный полифенизм у других бабочек: Самцы и самки меняются ролями при смене погоды, «Элементы», 27.12.2012 и Генетические основы эволюции фенотипической пластичности начинают проясняться, «Элементы», 09.11.2020). Представим себе, что у такого вида самцы живут дольше самок (что вполне вероятно для бабочек с их ZW-механизмом определения пола) и что поколения самцов перекрываются. Тогда мы будем наблюдать одновременное присутствие в популяции двух морф только у самцов.
К полиморфизму самцов может приводить и конфликт полов (Sexual conflict) (см. о нем, например, Самцы-насильники у гуппи производят низкокачественное потомство, «Элементы», 17.02.2012). Конфликт полов очень острый у жуков-плавунцов, у которых домогательства самцов нередко приводят к смерти самок (см. Самцы жуков-плавунцов — насильники и убийцы). Из-за этого появляются морфы самок с ребристой или гранулированной поверхностью надкрыльев, к которой самцам труднее прикрепляться своими присосками. А недавно на одном из видов, Graphoderus zonatus, было показано, что в ответ на это и среди самцов возникают морфы (см. L. Iversen et al., 2018. Sexual conflict and intrasexual polymorphism promote assortative mating and halt population differentiation). Правда, морфы эти не очень четко выраженные, и есть впечатление, что авторам работы очень хотелось их найти. У одной морфы крупнее три присоски наибольшего размера и многочисленнее остальные, более мелкие. Показано, что такие самцы чаще спариваются с «защищенными» самками с гранулированными надкрыльями, и их больше в популяциях, где процент таких самок выше. И модели, и длительные наблюдения показывают, что в этом случае может действовать балансирующий (см. Balancing selection) частотно-зависимый отбор, поддерживающий полиморфизм.
Можно представить себе, что у самок изменчивость (например, размеров) непрерывная, а у самцов есть две морфы — мелкая и крупная, и мелкая морфа успешнее спаривается с мелкими самками, а крупная — с крупными. Если размеры самок меняются (например, в зависимости от времени года или местообитания), то репродуктивное преимущество может переходить от одних самцов к другим. Возможно, так обстоит дело у некоторых морских бокоплавов.
Нередко полиморфизм возникает в связи с мимикрией — подражанием разным видам-моделям. Лучше всего он изучен у бабочек. Обычно разные морфы у бабочек-подражателей есть только среди самок, а самцы вообще не мимикрируют; есть разные возможные объяснения этого (см, например, J. R. G. Turner, 2008. Why male butterflies are non-mimetic: natural selection, sexual selection, group selection, modification and sieving). Но как минимум у одной бабочки — африканской нимфалиды Pseudacraea eurytus — есть миметические морфы, которые встречаются только среди самцов (хотя у самок таких морф гораздо больше). С чем связано это исключение — неизвестно.
А иногда полиморфизм связан с комбинацией мимикрии под разные модели и избирательным скрещиванием (см., например, Зафиксирован начальный этап видообразования у тропических бабочек, «Элементы», 09.11.2009). Естественно, возможны и другие комбинации причин...
Но наиболее распространенный вариант полиморфизма самцов, вероятно, все-таки связан с альтернативными репродуктивными стратегиями (когда поведение детерминировано генетически) или тактиками (когда оно определяется не генотипом, а чем-нибудь еще). Отчасти к ним можно отнести и ассортативные скрещивания, описанные выше; но нередко самцы отличаются не тем, каких самок они выбирают, а тем, какие способы используют для овладения ими.
Иногда тактики настолько сильно различаются, что самцы двух морф принадлежат к одной популяции, но никогда не встречаются друг с другом. Например, у общественной пчелы Macrotera portalis, строящей под землей крупные гнезда, есть две морфы самцов — крупные монстры, с огромной головой и острыми челюстями-мандибулами, и более мелкие, обычного для пчел облика. Мелкие вылетают из гнезда, сидят на цветках мальвовых рода Sphaeralcea (единственных растений, с которого самки собирают пыльцу), гоняют с них всех самцов своего и других видов пчел и спариваются с прилетающими самками. Драки между мелкими самцами жестокие, но не смертельные. Крупные самцы вообще не летают и на всю жизнь остаются в гнезде. У них есть крылья, но редуцированы летательные мышцы (зато очень мощные мышцы мандибул!) Сидя обычно в боковых ходах гнезда, эти самцы спариваются с самками, когда те прилетают в гнездо с пыльцой после окончания трудового дня, и незадолго до откладки самками яиц. Такая тактика может быть связана с приоритетом спермиев: у многих перепончатокрылых яйца преимущественно оплодотворяются спермиями последнего кавалера.
Между собой крупные самцы устраивают схватки, часто заканчивающиеся смертью одного из них. Но в крупных гнездах самцов может быть много; видимо, они как-то уживаются, если на каждого приходится как минимум по одной самке.
У крупных самцов явно более высок шанс оплодотворить самок. Но в новых гнездах до появления первого потомства, в течение трех недель, крупных самцов нет. А самки всё это время летают за пыльцой и спариваются с мелкими... Кроме того, самки могут сами «решать», сколько им сделать крупных и мелких самцов. Для выкармливания крупных явно требуется большая затрата сил (например, больше пыльцы в ячейке). И самке в определенных условиях может быть выгодно «наделать» побольше мелких самцов, даже если толку от них меньше.
Оказывается, так бывает нередко. Похожий диморфизм — летающие мелкие и остающиеся в гнезде или в соплодии, нелетающие, драчливые, способные поубивать друг друга самцы — есть также у разных видов фиговых ос, муравьев и пчел другого семейства.
Иногда разделение ниш самцов не такое четкое. Например, у нескольких видов саламандр найдены две морфы самцов — одни в период размножения отращивают специальные усики и ищут самок заранее, в лестной подстилке; другие отращивают мышцы челюстей (и чаще кусают других самцов) и ждут самок в ручьях, где те будут откладывать икру.
Очень поучительный пример — тактики самцов у клеща Caloglyphus berlesei. Две морфы самцов этого клеща различаются в основном строением ног третьей пары. Одни (назовем их «драчуны») имеют утолщенные и заостренные ноги, которыми они могут убивать соперников. У «мирных» самцов ноги обычные, как у самок. «Драчуны» появляются только при низкой плотности колоний клеща, при высокой плотности их развитие подавляется химическими сигналами. В эксперименте создавали колонии из двух драчунов и двух мирных (и четырех самок) и крупные колонии, где было по 30 самцов каждой морфы (и 60 самок). Оказалось, что в мелких колониях в большинстве случаев один из драчунов убивал всех стальных самцов и получал неограниченный доступ к самкам, и репродуктивный успех драчунов в целом был выше. Но в крупных колониях драчуны чаще вступали в схватки и убивали друг друга, в живых оставались мирные самцы, и теперь уже их репродуктивный успех был выше. Получается, что «первым парнем на деревне» легче стать за счет грубой силы, зашугав всех потенциальных соперников; но в городе такая тактика не слишком перспективна...
Во многих случаях, как выяснилось, морф с альтернативными стратегиями-тактиками не две, а три — и тогда между ними могут возникать отношения «камень, ножницы, бумага». Некоторые такие примеры описаны в статье Натальи Резник с таким названием. Например, у многих ящериц агрессивные, имеющие большую территорию самцы («камень») побеждают самцов с меньшей территорией («ножницы»), но проигрывают похожим на самок воришкам («бумага»). Самцы с меньшей территорией охраняют ее бдительнее и воришек не пропускают.
Пожалуй, у ящериц цветовой триморфизм изучен лучше всего, и по ним недавно вышел обзор (см. D. Stuart-Fox et al., 2020. Convergence and divergence in lizard colour polymorphisms). Поэтому кое-что можно добавить про этот интересный случай.
Оказалось, что триморфизм самцов с красным (оранжевым), желтым и голубым (серым/белым) горлом независимо появлялся едва ли не во всех главных эволюционных линиях ящериц. Физиологическая основа его во всех случаях одна и та же, а вот генетическая — видимо, во многих случаях разная (тут мы снова сталкиваемся с тем, что «гомологические ряды» не совсем гомологические, см. новость Параллелизмы — результат быстрой эволюции сенсорных рецепторов, «Элементы», 20.09.2011).
Выяснилось также, что не цвет определяет репродуктивную тактику. У пятнистобоких игуан (Uta stansburiana), о которых речь идет в статье Натальи Резник, самые агрессивные и имеющие наибольшую территорию и гарем самцы — оранжевые. А у другой игуаны, Urosaurus ornatus, это «синие» самцы.
Различаются разные виды и по предпочтениям самок. Например, у Urosaurus ornatus синие самцы выбирают желтых самок (у этого вида они либо желтые, либо оранжевые), а желтые («воришки») спариваются безвыборочно. А у обыкновенной стенной ящерицы (Podacris muralis), у которой самки имеют те же три морфы, только бледные (рис. 1), друг с другом чаще спариваются особи одной морфы.

Кроме репродуктивной тактики цвет может коррелировать с другими признаками — например, у этого вида синие самцы имеют более узкий спектр пищевых объектов.
Также нужно иметь в виду, что морф самцов на самом деле бывает больше, чем три: кроме трех основных морф есть одна или две промежуточных. Так что при дальнейшем их изучении вполне могут выявиться «ящерица и Спок».
А еще оказалось, что у Uta stansburiana синие самцы имеют наибольшую приспособленность не среди желтых (как описано в статье Резник), а среди синих, причем сходных генетически! Похоже, что они совместно защищают территорию от внедрения других самцов и чуть ли не способны к альтруизму в отношениях (см. Tim Stephens, 2003. Cooperation between unrelated male lizards adds a new wrinkle to evolutionary theory и Tim Stephens, 2006. Greenbeards with blue throats shed new light on the evolution of altruistic behavior).
У рыб триморфизм самцов тоже встречается. Например, у цихлиды Astatotilapia burtoni есть территориальные самцы — более крупные, агрессивные, размножающиеся и окрашенные либо в ярко-желтый, либо в ярко-синий цвет. Есть также более мелкие, с покровительственной буро-зеленой окраской, похожие на самок самцы, не защищающие территорий и, видимо, вообще не размножающиеся. Выяснилось, что любой самец может поменять цвет — стать из желтого синим или буро-зеленым и наоборот, из территориального — бродягой и наоборот, и это зависит, в частности, от исходов драк. Таким образом, это пример полифенизма.
Кроме изопод, жуков, рыб, ящериц и турухтанов триморфизм самцов описан у стрекоз, сенокосцев и клещей. Но боюсь, что решение и так получилось слишком длинным, и пора остановиться. А жаль! Сколько еще хороших примеров полиморфизма самцов осталось — почти как советов у ребе, когда все куры сдохли! Вот, например, у паука-скакунчика Maevia inclemens (рис. 2) одни самцы имеют три хохолка из щетинок на голове, а у других хохолков нет, зато есть желтые полосочки — и никто не знает, почему...

Рис. 2. Две морфы пауков-скакунчиков Maevia inclemens. Фото с сайта researchgate.net
Послесловие
Полиморфизм часто привлекает внимание и ученых, и популяризаторов науки. В одной старой наукометрической статье утверждается, что полиморфизму посвящено больше эволюционных статей, чем непрерывной изменчивости. И сайт «Элементы» не исключение. Перед тем как писать задачу, я просмотрел сайт и нашел как минимум четыре задачи и множество статей на похожую тему. А когда увидел и прочел статью Натальи Резник «Камень, ножницы, бумага» — почти отчаялся. Но потом решил, что отступать некуда и нужно разобрать полиморфизм самцов в более широком контексте.
Как мы видели, полиморфизм может возникать и/или (как минимум некоторое время) сохраняться без всякого отбора. Но ученых гораздо больше привлекают случаи, когда можно предположить (и изучить) действие на морфы естественного отбора — этого гениального художника и скульптора, создающего живые формы и раскрашивающего их в причудливые цвета. Публике ведь тоже интереснее смотреть на «Черный квадрат» Малевича, чем на похожий квадрат, случайно напечатанный на принтере. Если полиморфизм связан с размножением, то для ученых он — как «Черный квадрат» в квадрате. А уж если он есть только у одного пола — это как «Черный квадрат» в кубе!
Кстати, полиморфизм самок, видимо, привлекает меньше внимания ученых, чем полиморфизм самцов; это утверждается, например, в цитированном недавнем обзоре по ящерицам (см. Решение). И тут мужской шовинизм! (Справедливости ради нужно отметить, что для ящериц как раз не известны примеры, когда полиморфизм есть только у самок, но у других животных такое бывает сплошь и рядом.)
При изучении полиморфизма есть где развернуться и теоретикам, которые строят модели эволюции, и полевым биологам, наблюдающим за поведением животных в природных условиях, и — в последние десятилетия — молекулярным биологам.
Поразительно, насколько широко распространились и глубоко проникли в полевую биологию молекулярные методы. Они позволяют установить, кто чей родственник, — например, сколько детей в каждом гнезде принадлежит самцу, который за ними ухаживает. О такой детализации недавно нельзя было даже мечтать! А еще они позволяют постепенно приблизиться к пониманию физиологических механизмов появления морф на молекулярно-генетическом уровне. В качестве примера сошлюсь на статью Натальи Резник про морфы самцов турухтанов (см. Три самца и одна инверсия). У них в результате инверсии и делеций возникли три варианта супергена, детерминирующего фенотипы трех морф самцов. Участок, содержащий множество генов, наследуется как единое целое, как будто признак моногенный. Неудивительно, что при таком наследовании окраска и прочие внешние признаки коррелируют со множеством других показателей — от плодовитости и состава пищи до тонких особенностей полового поведения. К сожалению, именно в таких случаях установить вклад разных генов в фенотип сложно.
Конечно, полиморфизм привлекает и любителей разложить всё по полочкам — дать разумную классификацию этого явления и придумать терминологию для его описания. Нужно сказать, что им приходится нелегко.
Как почти везде в биологии, встречается масса комбинированных и промежуточных вариантов изменчивости. Во многих случаях трудно отличить непрерывную изменчивость от полиморфизма. Например, до сих пор идут споры о том, есть ли цветовые морфы у самок нашей ящерицы-живородки Zootoca vivipara или ее изменчивость непрерывная.
Другой пример: у многих видов кальмаров есть две четких морфы самцов, которых принято называть «консорты» и «сникеры» («крадущиеся»), см. картинку дня Консорты и сникеры. Первые крупнее, спариваются с самками в параллельной позиции, сперматофоры запихивают в их мантийную полость, поближе к месту выхода яиц; вторые, мелкие, спариваются «лицом к лицу» (по-тихому, не вступая в конфликты с консортами), сперматофор прикрепляют к буккальной мембране. И форма сперматофора, и форма сперматозоидов, — ну всё у них разное, явно две четкие морфы. Но недавно у кальмара Doryteuthis pleii между этими двумя морфами обнаружилась «промежуточная» морфа — и оказалось, что переход от одной тактики к другой зависит от возраста и происходит по мере роста (см. L. H. Apostolico, J. Marian, 2019. Behavior of «Intermediate» Males of the Dimorphic Squid Doryteuthis pleii Supports an Ontogenetic Expression of Alternative Phenotypes и обзор J. Marian et al., 2019. Male Alternative Reproductive Tactics and Associated Evolution of Anatomical Characteristics in Loliginid Squid). Или вот цихлида Melanochromis auratus — посмотришь на картинку, и очевиден диморфизм самцов (рис. 3).
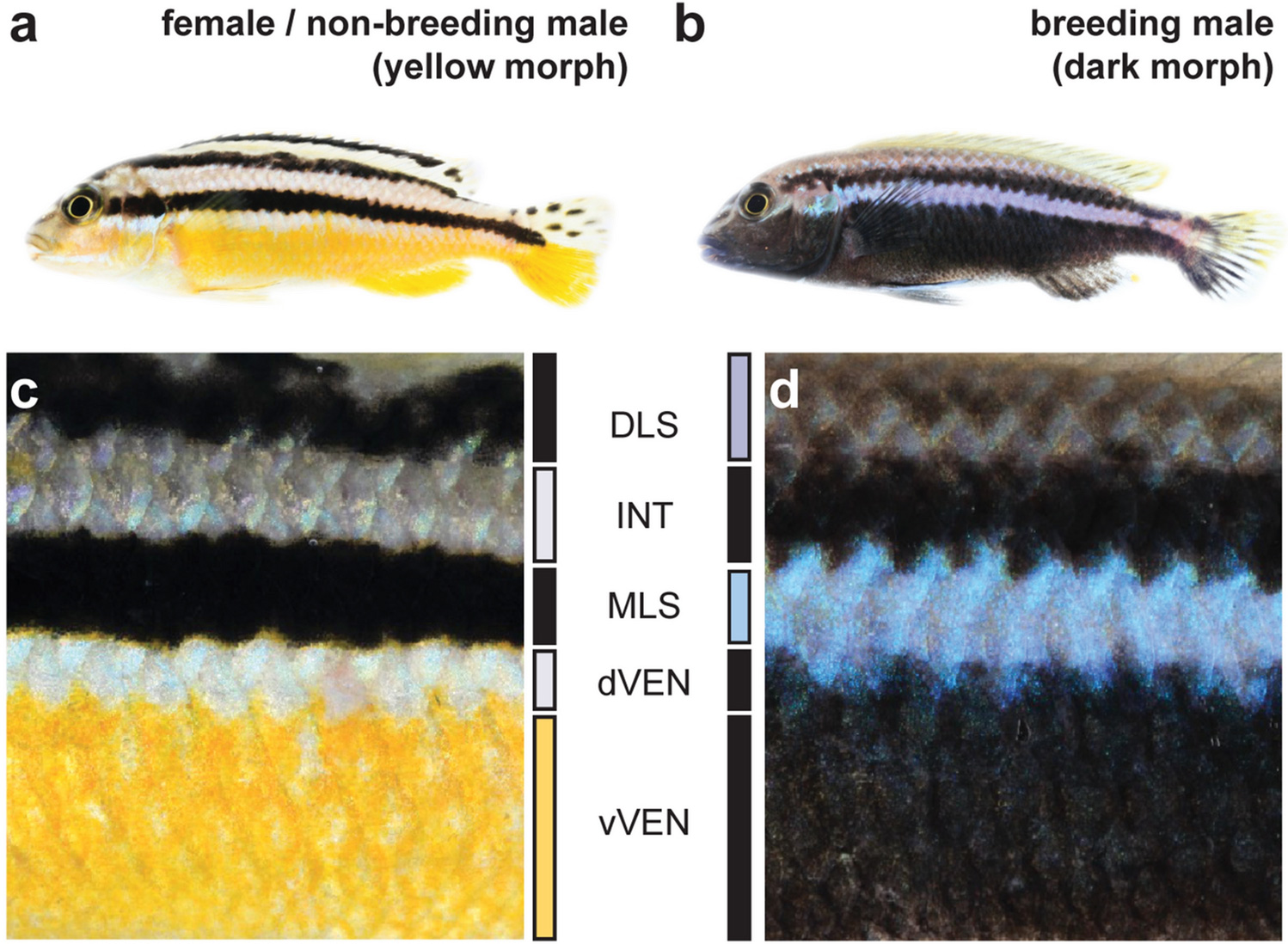
Рис. 3. Цветовые морфы самцов Melanochromis auratus — результат изменения окраски. Обратите внимание, что у доминантных самцов (справа) на месте белой полоски появляется черная, а на месте черной — голубая. Изображение из статьи Y. Liang et al., 2019. Neural innervation as a potential trigger of morphological color change and sexual dimorphism in cichlid fish
На самом деле цвет, как и в рассмотренном в решении примере, может меняться, и обычно он меняется с возрастом, когда самцы-субординанты становятся доминантными. Понятно, что меняется он не быстро, как у хамелеона или камбалы, — такие изменения окраски никто полиморфизмом не считает. При смене наряда меняется число клеток-хроматофоров, их структура и даже количество аксонов, иннервирующих чешуйки кожи, — у темной морфы их становится почти вдвое больше! Но значит, очевидно, есть и промежуточные варианты окраски...
Однако мы опять увлеклись примерами. А каковы все-таки наиболее частые механизмы подержания полиморфизма самцов и их альтернативных тактик/стратегий? Согласно одной из классификаций, при равной приспособленности двух морф это может быть либо балансирующий частотно-зависимый отбор, либо дизруптивный отбор. Если же приспособленности морф неравные, то равновесие возможно, если преимущество переходит от одной морфы к другой во времени или в пространстве или если наблюдается преимущество гетерозигот (см. Heterozygote advantage).
При балансирующем частотно-зависимом отборе преимущество получает более редкая морфа, поскольку каждый тип самцов проигрывает в окружении себе подобных. Есть множество примеров такого отбора (в том числе полового), один из них был описан в послесловии к задаче Улитка в Зазеркалье для наземной улитки Amphidromus inversus. Похоже, что у самых разных животных этот тип обора может приводить к сходным результатам.
Чаще всего встречается вариант, когда есть территориальные самцы, которые честно вкладываются в «самцовые» признаки, в защиту территории и «своих» самок, и подражающие самкам обликом и поведением «воры», которые прокрадываются на чужую территорию и оплодотворяют часть самок. Подобный диморфизм описан у самых разных животных — от стрекоз и жуков-навозников до ящериц, птиц и орангутанов. Но при этом «самцы-воришки» похожи на самок далеко не во всём. Чтобы это проиллюстрировать, рассмотрим еще один пример.
У северной рыбы-мичмана (Porichthys notatus) одни самцы крупные, строят на литорали гнёзда, привлекают самок пением и сторожат кладки; другие мелкие, ничего не строят, кладок не стерегут и не поют. Кося под самку, они либо принимают «позу откладки яиц» и оплодотворяют часть яиц, откладываемых в этот момент самкой, либо просто лежат неподалеку от чужого гнезда, выпуская сперматозоиды в воду. Крупные самцы иногда тоже пытаются подбираться к чужим гнездам; но их быстро прогоняют хозяева, что наглядно демонстрирует преимущество мелких самцов при подобной тактике.
У крупных самцов лучше развиты «голосовые» мышцы и крупнее управляющие ими нейроны. Зато относительные размеры семенников у мелких самцов в 9 раз больше, чем у крупных! И похоже, что это тоже обычная составляющая их «тактики».
Тут наша история приобретает неожиданный поворот. Ведь все, о ком шла речь до сих пор, — никакие не самцы! Это бесполое диплоидное поколение, временное вместилище для настоящих самцов-сперматозоидов. Да-да, гаметы — это вовсе не клетки, а отдельное, гаплоидное поколение нашего жизненного цикла. Вот у кого нужно искать полиморфизм! Ведь сперматозоиды тоже конкурируют за право оплодотворения самок-яйцеклеток (см. Половой отбор сделал сперматозоиды дрозофил самыми длинными в мире, «Элементы», 01.06.2016), подвергаются половому отбору и даже способны проявлять альтруизм, помогая сородичам (см. Взаимопомощь среди сперматозоидов основана на семейственности, «Элементы», 29.01.2010). И конечно же, у гамет полиморфизм тоже известен — как среди самок, так и среди самцов. Сперматозоиды могут брать не только числом, но и умением. Например, у рыб мелкие самцы-воришки (рыбы, а не гаметы) обычно выпускают свои сперматозоиды дальше от гнезда, и логично предположить, что они должны лучше плавать и/или дольше жить. Часто это так и есть. А у цихлиды Lamprologus callipterus всё наоборот — ее крупные «самцы-буржуи» (которые во много раз крупнее самок) натаскивают в гнездо раковины улиток, в которые залезают самки для нереста, и выпускают сперматозоиды снаружи; мелкие «самцы-воришки» залезают внутрь и выпускают сперматозоиды в среднем в 6 раз ближе к икринкам (см. M. Taborsky et al., 2018. Alternative male morphs solve sperm performance/longevity trade-off in opposite directions). У карликовых самцов и сперматозоиды помельче, и они стараются поскорее доплыть до икринок по прямой, но быстро выбиваются из сил. Крупные сперматозоиды крупных самцов сначала сильнее виляют, но зато дольше сохраняют приличную скорость и дольше живут.
Даже беглый взгляд на литературу показал, что полиморфизм сперматозоидов — явление распространенное и довольно разнообразное. Например, нередко несколько типов сперматозоидов-самцов производит один самец-многоклеточное (см. Sperm heteromorphism). Так что ждем описания стратегий «камень, ножницы, бумага» среди сперматозоидов.





Рис. 1. Самцы Podacris muralis чаще спариваются с самками того же цвета. Фото с сайта uv.es