Необыкновенные приключения водорослей

Задача
Водоросли-динофлагелляты (они же панцирные жгутиконосцы) — одна из важнейших групп протистов. Существуют свободноживущие, паразитические и симбиотические виды этих протистов, а среди свободноживущих есть как гетеротрофные (не способные синтезировать органические вещества из неорганических), так и фотоавтотрофные (использующие свет для получения энергии) представители. Фотосинтезирующие динофлагелляты в мировом океане — одни из главных продуцентов органики. Привлекают они внимание ученых и тем, что часто вызывают цветение воды — «красный прилив», и среди видов, дающих такие вспышки, есть сильно ядовитые. Некоторые виды выделяют смертельно опасный сакситоксин, а некоторые — менее опасные, но всё равно малоприятные токсины, вызывающие диарею. В организм человека эти яды могут попасть, если их накапливают поедающие динофлагеллят съедобные двустворчатые моллюски.
И вот один из таких ядовитых видов водорослей-динофлагеллят никак не удавалось для более подробного изучения получить в лабораторных культурах. Вообще-то, большинство водорослей культивировать легко: нужна стандартная питательная среда, содержащая необходимые растениям минеральные соли да еще пару-тройку витаминов. Но наши динофлагелляты упорно не желали расти на такой среде, хотя нормально росли в лаборатории в смешанных пробах планктона.
Проблему удалось частично решить, когда к культуре водорослей добавили один из видов морских инфузорий. В этих условиях динофлагелляты росли и размножались, но через некоторое время двухвидовые культуры угасали. А вот если в культуру добавляли еще один-два определенных вида морских одноклеточных водорослей — тогда динофлагеллят удавалось культивировать долго. Как можно объяснить такую необычную «привередливость» панцирного жгутиконосца?
Подсказка 1
Инфузории мельче динофлагеллят, а «дополнительные» водоросли мельче инфузорий.
Подсказка 2
Все эти виды имеют похожую красно-коричневую окраску.
Подсказка 3
Инфузорий не удается культивировать в темноте.
Решение
Действующие лица нашего детектива — это динофлагеллята Dinophysis acuminatа (динофизис), инфузория Myrionecta rubra (мирионекта) и криптофитовые водоросли (криптомонады) Geminigera и Teleaulax. Сюжет их «необыкновенных приключений» лучше всего описывается поговоркой «вор у вора дубинку украл». Почти целиком этот сюжет описан (на английском) в научном блоге Sceptic Wonder, а русский перевод есть здесь. Вкратце этот сюжет заключается вот в чем: инфузории едят криптомонад, а динофизисы — инфузорий. От съеденных жертв внутри хищников остаются работающие хлоропласты. «Ворованные» хлоропласты (по-научному — клептопластиды; см. Клептопластия) передаются по пищевой цепочке. Ни мирионекты, ни динофизисы не выживают без фотосинтеза клептопластид, получаемых от криптомонад.
Теперь рассмотрим эту детективную историю поподробнее.
Инфузория Myrionecta rubra — один из самых массовых видов морских планктонных инфузорий, иногда даже вызывающий красные приливы (см. также Red tide). До самого конца XX века предполагали, что M. rubra имеет постоянных фотосинтезирующих эндосимбионтов, передающихся потомству при делении клетки. Но в 2000 году было доказано, что M. rubra — облигатный миксотроф. Выяснилось, что для роста и размножения она должна постоянно поедать криптомонад.
Почти все органические вещества для роста инфузории при этом получают благодаря фотосинтезу съеденных жертв — из-за этого их не удается долго культивировать в темноте.
Самое удивительное в питании инфузорий — то, что они не переваривают не только пластиды, но и другие органоиды жертв (см. рис. 1). Работающие хлоропласты криптомонад сохраняются в цитоплазме инфузорий месяцами и даже, по некоторым данным, делятся. Возможно, это потому, что в клетке мирионекты сохраняются транскрипционно активные ядра криптомонад. Если клептопластия распространена довольно широко, то использование ворованных ядер — уникальное явление, известное пока только для мирионекты. Авторы, описавшие это явление, назвали его кариоклептией.

Мирионекты, видимо, фагоцитируют своих жертв (хотя подробно процесс их питания, кажется, не описан). А затем загадочным путем органоиды жертв — ядра, хлоропласты и митохондрии — «секвестрируются» в особые компартменты внутри клетки инфузории, окруженные мембраной — ядра отдельно, а митохондрии с хлоропластами отдельно (рис. 1). Как нужные белки направляются из ядер криптомонад в хлоропласты и митохондрии (если это действительно происходит) — неизвестно.
Динофлагелляты рода Dinophysis бывают гетеротрофными и фотоавтотрофными. Dinophysis acuminatа считались облигатными фотоавтотрофами. Но при этом попытки их культивирования в течение 15 лет не приводили к успеху: ни обогащенная среда, ни добавление разных мелких жертв (см. послесловие) не позволяли поддерживать культуру дольше, чем в течение 4–6 циклов делений клеток. Как написано в одном из обзоров, «такие работы сопряжены с высоким риском: никто не захочет, чтобы его PhD-исследователь месяцами возился над и не получил никакого определенного результата» (см. An IOC Newsletter on toxic algae and algal blooms, No 32 ).
Давно существовало подозрение, что пластиды динофизис «ворует». Его пластиды резко отличаются от «обычных» пластид динофлагеллят — имеют две, а не три наружные мембраны, не содержат перидинина, зато содержат фикобилины и хлорофилл c, характерные для криптомонад. Но среди динофлагеллят немало видов с третичными пластидами. Это значит, что свои вторичные пластиды, происходящие от красных водорослей (см. Возникновение и эволюция пластид, рис. 2 и послесловие) они утратили, а взамен приобрели какие-нибудь другие. Так что оставалось неясным, имеет ли динофизис собственные третичные пластиды или клептопластиды.
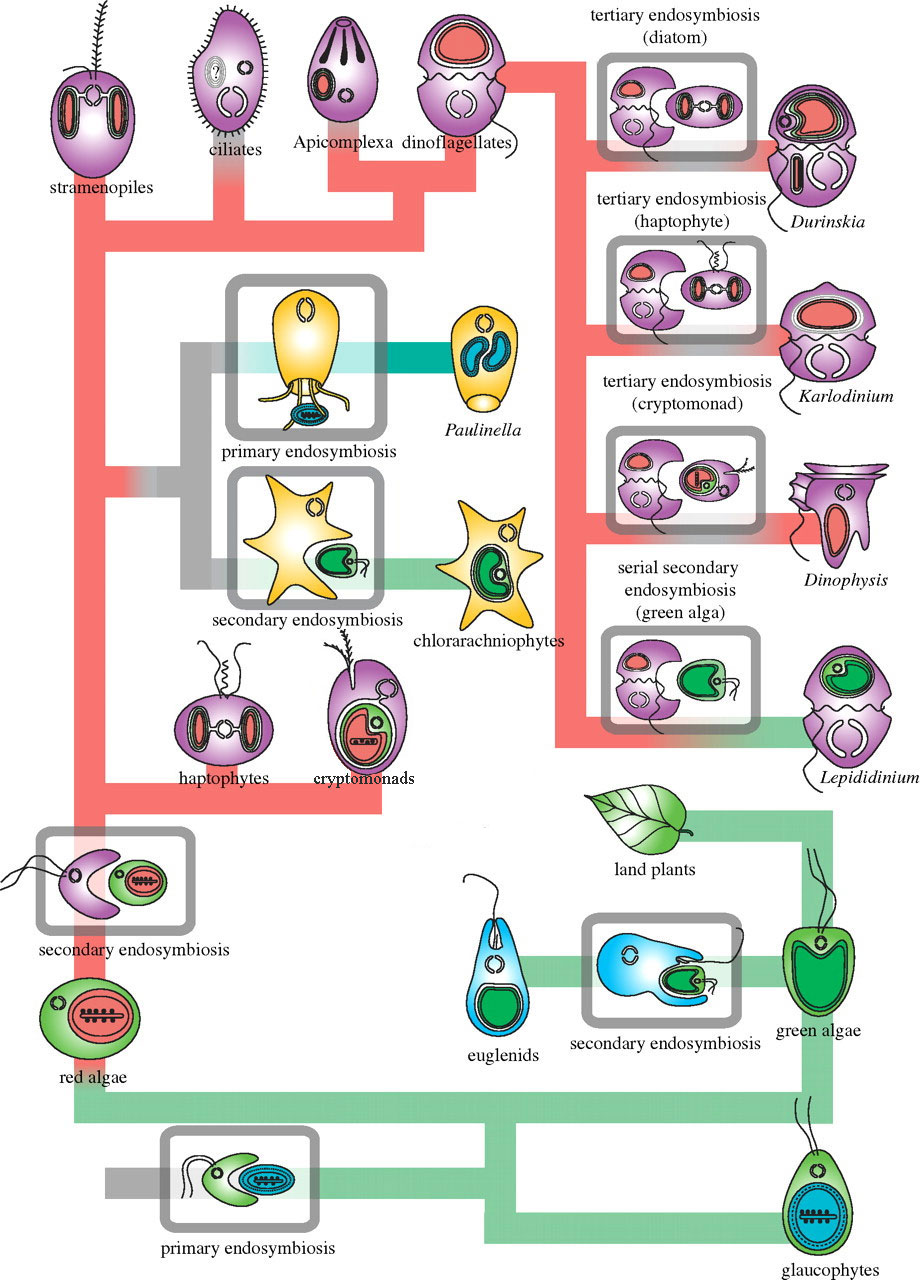
Рис. 2. Общая схема симбиозов среди пластидных организмов. Первичный симбиоз (primary endosymbiosis) происходил дважды: между праводорослью из царства Archaeplastida и неизвестной цианобактерией, от которой произошли все пластиды основной ветви, и между эукариотической Paulinella и цианобактерией из порядка Prochlorales. После первичного симбиоза царство Archaeplastida разделилось на три таксона: Glaucophyta, Rhodophyceae и Chlorophyta. После этого события произошел вторичный симбиоз (secondary endosymbiosys): часть красных и зеленых водорослей стала эндосимбионтами нефотосинтезирующих организмов из других клад. Таким образом, водоросли из таксонов Dinoflagellata, Chromerida, Apicomplexa, Stramenopila, Haptophyta и Cryptophyta получили пластиду от красных водорослей, a Euglenida и Chlorarachniophyta — от зеленых. Помимо вторичного симбиоза в кладе Dinoflagellata происходил третичный симбиоз (tertiary endosymbiosis), при котором пластида от красной водоросли была заменена или, в случае рода Kryptoperidinium, дополнена пластидой из других водорослей. Серый цвет линии обозначает беспластидные организмы, зеленая и красная линии — соответственно ветвь зеленых и красных водорослей, ? — сомнения в однократности события. Иллюстрация к статье P. J. Keeling, 2010. The endosymbiotic origin, diversification and fate of plastids, с изменениями
Только в 2006 году удалось получить стабильную культуру наших динофизисов — для этого в среду добавляли инфузорий M. rubra и криптомонад. Удалось установить, что динофизисы как-то ловят мирионект, хотя те могут очень быстро и далеко «отпрыгивать» за счет ударов ресничек. (В 2016 году для другого вида динофизисов было показано, что они выделяют слизь, в которой запутываются инфузории, см. G. Gimenez Papiol, 2016. The use of a mucus trap by Dinophysis acuta for the capture of Mesodinium rubrum prey under culture conditions; возможно, это справедливо и для D. acuminatа). Потом динофизисы протыкают покровы клетки инфузории специальным хоботком и поглощают ее содержимое с помощью особого способа фагоцитоза — мизоцитоза (см. Myzocytosis). При мизоцитозе содержимое жертвы постепенно высасывается (его еще называют «клеточный вампиризм») и оказывается внутри клетки хищника. Динофизис становится всё толще, а от жертвы остается только «шкурка» в виде бесформенного комочка. Как питается динофизис, можно посмотреть на видео.
После переваривания инфузорий от пластид остается лишь их внутренняя часть: получается хлоропласт, окруженный двумя мембранами (рис. 1) и при этом содержащий набор пигментов, свойственных криптомонадам.
Забавно, что способ культивирования динофизисов был запатентован. Что ж, пожалуй, его и вправду можно считать изобретением.
Конечно, как во многих наших задачах, условие (даже вместе с подсказками) позволяет предположить и другие варианты ответа. Например, инфузории и динофлагелляты могут есть криптомонад, а динофлагелляты могут, кроме того, получать от инфузорий какие-то необходимые ростовые вещества, которые те выделяют в воду (можно придумать и множество других аналогичных вариантов).
Послесловие
Мы разобрали только современную часть нашего детектива. Но необыкновенные приключения водорослей начались давно. Точнее, очень давно: «давным-давно-предавно (когда свиньи пили вино, гуси (по другим версиям — утки или мартышки) жевали табак, а куры его глотали и от этого жёсткими стали)». Было это точно больше миллиарда лет назад, а может, и ближе к двум миллиардам. В роли гетеротрофных кур выступали некие древние эукариоты, а в роли фотоавтотрофного табака — цианобактерии. Так возникли первичные пластиды красных, зеленых и глаукофитовых водорослей, а клетки самих водорослей стали жесткими, приобретя клеточную стенку, — всё как в сказке.
Но сказка на этом не кончилась: «кур» через пару сотен миллионов лет наглотались другие персонажи («лисы», «волки» и «медведи»). Так возникли вторичные пластиды — у предков криптомонад и динофлагеллят из проглоченных красных водорослей, а у предков эвгленовых и хлорарахниевых — из зеленых (см. Возникновение и эволюция пластид).
Надо отметить, что «куры» внутри волков-криптомонад почти не изменились. Вторичные пластиды криптомонад сохранили общую схему строения эукариотической клетки. Они окружены четырьмя мембранами, причем между второй и третьей снаружи есть ядро (нуклеоморф) и эукариотические рибосомы (см. P. R. Gilson, 2001. Nucleomorph genomes: much ado about practically nothing). Правда, нуклеоморф больше уже не может обеспечивать жизнь хлоропласта: как и из цианобактерии, большинство генов из него переместились в «центральное правительство» — в ядро клетки криптомонады. Это и понятно: если в твоем желудке живут куры (да еще с табаком внутри, способным их прокормить!), лучше держать их размножение под строгим контролем.
Расшифровка строения пластид криптомонад — настоящий триумф теории симбиогенеза (см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). Уж такие-то органоиды (а это именно органоиды!) никак не могли возникнуть внутри клетки, эндогенным способом. И если любая клетка эукариот — мультигеномная система, то криптомонады мультигеномны в квадрате. В их клетке совместно функционируют четыре генома: 1) геном митохондрий, 2) геном ядра криптомонады, 3) геном первичной пластиды — бывшей цианобактерии, 4) геном нуклеоморфа — бывшего ядра кранной водоросли. Ну а мирионекты полигеномны в кубе. В их клетке постоянно присутствуют семь геномов: к геномам съеденных криптомонад добавляются геномы митохондрий инфузории и двух ее ядер, большого и малого.
В древних сериях «необыкновенных приключений» привередливость динофлагеллят тоже проявилась. Она проявилась в том, что многие из них избавились от своих «исконных» вторичных пластид (в прошлом красных водорослей), а потом приобрели третичные пластиды — из криптомонад, гаптофитовых, диатомовых или зеленых водорослей (см. P. J. Keeling, 2004. Diversity and evolutionary history of plastids and their hosts и рис. 2 из решения).
Правда, сейчас часть этих данных под вопросом. Может быть, и в других случаях (а не только у динофизиса) это не третичные пластиды, а клептопластиды?
Да и наш современный детектив далеко еще не закончен. До сих пор продолжают выходить статьи, доказывающие, что в мирионекте живут целые криптомонады, а у динофизисов есть собственные стабильные пластиды. Несмотря на применение современных методов геномики и ПЦР отдельных клеток, доказать или опровергнуть это достаточно сложно — особенно учитывая тот факт, что наличие собственных пластид или эндосимбионтов не мешает промышлять клептопластией (рис. 3). К тому же совершенно непонятно, как ядра и пластиды криптофитовых попадают в цитоплазму инфузорий и оказываются внутри мембранных компартментов. Так что сомнения в том, «кто преступник», остаются...

Рис. 3. Третичный эндосимбиоз, клептопластия или их сочетание?
Одна из интересных проблем — поддержание активности «чужих» органоидов внутри клетки-хозяина. Если инфузории, вероятно, используют для этого «ворованные ядра» (в них первые 20–30 дней после попадания в инфузорию сверхактивно экспрессируются гены, производящие белки для хлоропластов криптомонады), то у динофизисов такой способности нет — а чужие пластиды работают в их клетке до трех месяцев. Естественно, начались поиски «пластидных» генов криптомонад в ядре динофизиса. Как минимум один такой ген нашли (нашли и другие пластидные гены, но из других водорослей). А в недавней работе утверждается, что и в клетках динофизиса клептопластиды делятся (P. M. Rusterholz et al., 2017. Evolutionary transition towards permanent chloroplasts? — Division of kleptochloroplasts in starved cells of two species of Dinophysis (Dinophyceae)). Если это правда — это первый доказанный случай деления клептопластид в отсутствие ядер их исконной клетки. Но со временем число и объем клептопластид всё равно снижаются, и насколько им помогают ядерные гены динофизиса, неизвестно.
Неясная картина и с ролью ядерных генов в культивировании пластид у «фотосинтезирующего моллюска» Elysia chlorotica (см. картинку дня Фотосинтезирующий моллюск-листик и ссылки в ней). Похоже, ни один известный медведь, съевший и переваривший волка, не научился как следует культивировать его кур...
Исследования динофлагеллят несколько размыли понятия «своего» и «чужого» (речь идет только о пластидах, а не об общечеловеческой морали!). Но размытой оказалась и еще одна важная граница — между продуцентами и консументами.
Все мы должны знать со школы о пищевых цепях. И в школе учат, что в них поток вещества и энергии направлен всегда от продуцентов к консументам. Оказывается — ничего подобного! Очень многие (а может быть, и все!) динофлагелляты, имеющие вполне нормальные хлоропласты — миксотрофы. некоторые по-настоящему охотятся на жертв. Так что выражение «хищная водоросль» уже давно не режет слух морским экологам.
Многие процессы в океанах, в том числе образование опасных «красных приливов», могут быть лучше объяснены и предсказаны с учетом миксотрофного питания динофлагеллят (рис. 4). Они поедают самых разных гетеротрофов — от бактерий до крупных инфузорий и других динофлагеллят. У некоторых есть даже два рта — один для крупной добычи в районе пояска, в центре клетки, и один маленький, для бактерий, на ее переднем конце. Это — волк, коза и капуста в одном лице. А в биологии почти никогда не бывает резких границ...

Рис. 4. Миксотрофное питание может объяснять высокую скорость размножения многих динофлагеллят. Потребляя азотфиксирующих цианобактерий и гетеротрофных бактерий, богатых фосфором, они получают дефицитные биогенные элементы. Из статьи H. J. Jeong et al., 2010. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs







Рис. 1. История клептопластид Dinophysis. Инфузория M. rubra, поедая криптомонад, длительное время сохраняет в цитоплазме их вторичные пластиды, ядра и митохондрии. Когда Dinophysis поедает инфузорий, он сохраняет только бывшую цианобактерию — внутреннюю часть пластид криптомонад, две наружные мембраны и нуклеоморф разрушаются. 1 — ядро криптомонады; 2 — ее вторичный хлоропласт; 3 — нуклеоморф; 4 — митохондрия криптомонады; 5 — ядро и цитоплазма криптомонады, окруженные мембраной инфузории (а поверх нее — еще двумя мембранами эндоплазматической сети инфузории. — Прим. С.Г.); 6 — ядро инфузории (на самом деле в клетке M. rubra, как и у других инфузорий, два ядра — малое и большое. — Прим. С.Г.); 7 — пластидно-митохондриальный комплекс, окруженный мембраной инфузории; 8 — митохондрия инфузории; 9 — ядро динофлагелляты; 10 — клептопластиды; 11 — митохондрия динофлагелляты. Вверху — фотоснимки клеток, длина масштабного отрезка — 10 мкм. Из статьи J. H. Wisecaver and J. D. Hackett, 2010. Transcriptome analysis reveals nuclear-encoded proteins for the maintenance of temporary plastids in the dinoflagellate Dinophysis acuminata