Интеллект у растений есть!
Наталья Резник,
кандидат биологических наук
«Химия и жизнь» №9, 2024

От Дарвина до наших дней
Чарльз Дарвин не одной теорией эволюции прославился. Круг его научных интересов был весьма широк, в него входили в том числе движения растений, которые произвели на естествоиспытателя большое впечатление. В 1880 году в работе «Сила движения растений» он писал: «В растениях нет более замечательного — в том, что касается функции — органа, нежели кончик корешка... Едва ли будет преувеличением сказать, что кончик корешка действует как мозг какого-нибудь из низших животных, получая впечатления от органов чувств и направляя в соответствии с ними свои движения».
Мозга у растений нет, это мы знаем точно, а вот есть ли у них интеллект? Этот вопрос интересует многих ученых, о некоторых исследованиях мы уже писали (см. статью «Зеленый ум», «Химия и жизнь» 2016, №6). Сейчас принято считать, что растения способны обучаться, у них есть память и они могут корректировать свое поведение в зависимости от обстоятельств.
Однако вопрос окончательно не решен, и для его конструктивного обсуждения прежде всего необходимо договориться, что такое интеллект. Определения этого сложного явления много лет собирали два специалиста в области искусственного интеллекта, Шейн Легг и Маркус Хаттер (Shane Legg, Marcus Hutter), и собрали их более семидесяти. Основываясь на этих определениях, они предложили свое собственное. Интеллект — это свойство, которым обладает отдельный агент при взаимодействии со своей средой или средами. Эту формулировку взял за основу профессор Корнельского университета, эколог Андре Кесслер (André Kessler), изучавший защитные стратегии растений и задумавшийся над тем, есть ли у них интеллект. Формулировка тем хороша, что не предполагает наличия нервной системы; агентом может быть любое существо и даже нечто неодушевленное, так что для растений она подходит.
Согласно Леггу и Хаттеру, интеллект проявляется во взаимодействии со средой, а именно он «определяет способность агента достигать целей в широком диапазоне сред». Какие же цели у растения? Многие ученые полагают, что цель живого существа состоит в достижении максимальной приспособленности. Иными словами, живое существо должно наилучшим образом адаптироваться к окружающим условиям, порой весьма изменчивым, чтобы оставить как можно больше жизнеспособных потомков. Взаимодействия растений со средой сложны и разнообразны, и мы рассмотрим лишь одно — взаимоотношения с травоядными насекомыми, которых далее для краткости будем называть просто травоядными. Но вы, пожалуйста, помните, что речь идет о личинках бабочек и жуков, а не о коровах. Исследований на тему «растения и травоядные» очень много, я пользовалась преимущественно публикациями Андре Кесслера и профессора Калифорнийского университета Ричарда Карбана (Richard Karban), энтомолога.
Применительно к взаимодействию с травоядными интеллект растений должен проявляться как способность адекватно среагировать на появление вредителей. А как на них реагировать? Защищаться надо, потому что убежать растение не может. А защищаются они по-разному.
Многие растения постоянно синтезируют разнообразные защитные соединения. Примером могут служить глюкозинолаты, которые вырабатывают представители семейства Капустные. Глюкозинолаты состоят из трех блоков: фрагмента β-тиоглюкозы, фрагмента сульфированного оксима и вариабельной боковой цепи. Защитные свойства глюкозинолатов усиливаются при гидролизе растительным ферментом мирозиназой. Когда гусеница жует лист, глюкозинолаты, которые хранятся в клеточных вакуолях, вступают в контакт с мирозиназой, а мирозиназа их расщепляет с образованием ядовитых серосодержащих продуктов.
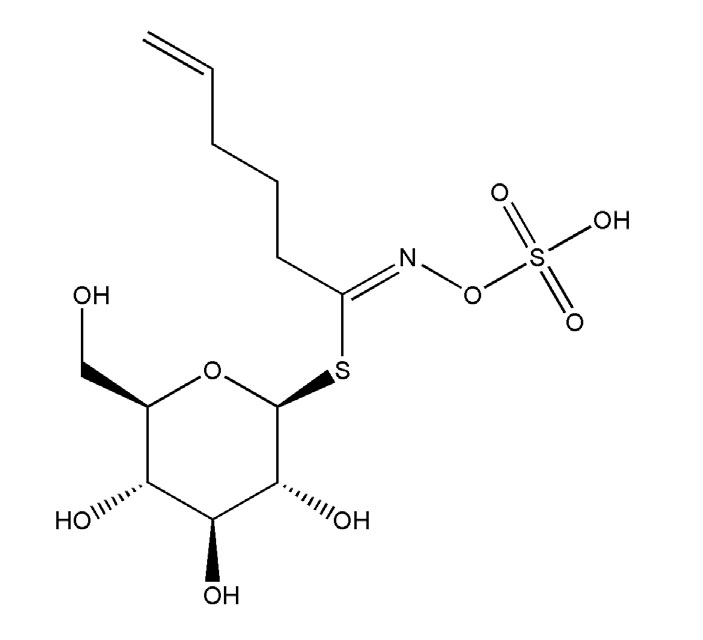
Глюкобрассиканапин. Глюкозинолаты — химическое оружие растений из семейства Капустные
Постоянную защиту называют конститутивной. Наряду с ней существует защита индуцированная, когда растение вырабатывает защитные вещества только при нападении травоядных. Эти способы друг друга не исключают, и при атаке вредителей та же капуста может усилить синтез одного или нескольких глюкозинолатов.
Сигналы тревоги
Чтобы реагировать на появление вредителей, нужно понять, что на тебя напали. Информацию об окружающей обстановке растения воспринимают с помощью химических сигналов разной природы: фрагментов макромолекул из разрушенных растительных тканей; слюны, содержимого полости рта, отрыжки или фекалий травоядных. Бобовые, например, распознают инцептины — фрагменты одного из растительных ферментов, которые оказываются во рту насекомого, переварившего лист. А у жующих гусениц малой совки во рту волицитин (N-17-гидроксилиноленоил-l-глютамин).
Эти сигнальные молекулы, попадая на поврежденный лист, взаимодействуют с растительными гормонами, которые в свою очередь активируют защитные гены растения. Фитогормоны жасмонаты — сложные эфиры и соли жасмоновой кислоты — обеспечивают устойчивость к тем, кто повреждает или жует листья: личинкам насекомых; грибам, вызывающим некроз ткани, и нематодам. Салициловая кислота повышает устойчивость к вирусам и некоторым видам насекомых, которые питаются соком растений, прокалывая жилки, например к тлям. Если одновременно активировать синтез обоих гормонов, они будут мешать друг другу. Но если активация происходит в разное время, взаимодействие жасмоновой и салициловой кислот усиливает защитный эффект.
Например, классический модельный объект арабидопсис Arabidopsis thaliana заразили бактерией Pseudomonas syringae. Эта инфекция активирует синтез салициловой кислоты, которая запускает гибель клеток вблизи места заражения. Местный некроз не позволяет бактерии распространяться, но при этом неизбежно привлекает других патогенов, которые питаются мертвой растительной тканью. Однако растение готово к такому повороту: вслед за активизацией салициловой защиты срабатывает жасмоновая, которая усиливает работу генов, противостоящих некрофилам. В данной ситуации оба защитных механизма действуют сообща.
Иногда к ним подключается еще один гормон — этилен. Его влияние также зависит от того, кто нападает на растение. Когда дикий табак Nicotiana attenuata заражают патогенные грибы, совместное действие жасмоновой кислоты и этилена вызывает синтез фунгицидов. Ситуация меняется при нападении гусениц табачного бражника Manduca sexta. Обычно в случае атаки насекомых растение синтезирует алкалоид никотин, губительный для гусениц. Но табачный бражник к никотину устойчив, и тратить силы на его синтез смысла нет. В такой ситуации фитогормоны действуют как антагонисты: этилен не позволяет жасмоновой кислоте повысить уровень никотина в табачных листьях; вместо этого срабатывают иные защитные механизмы.
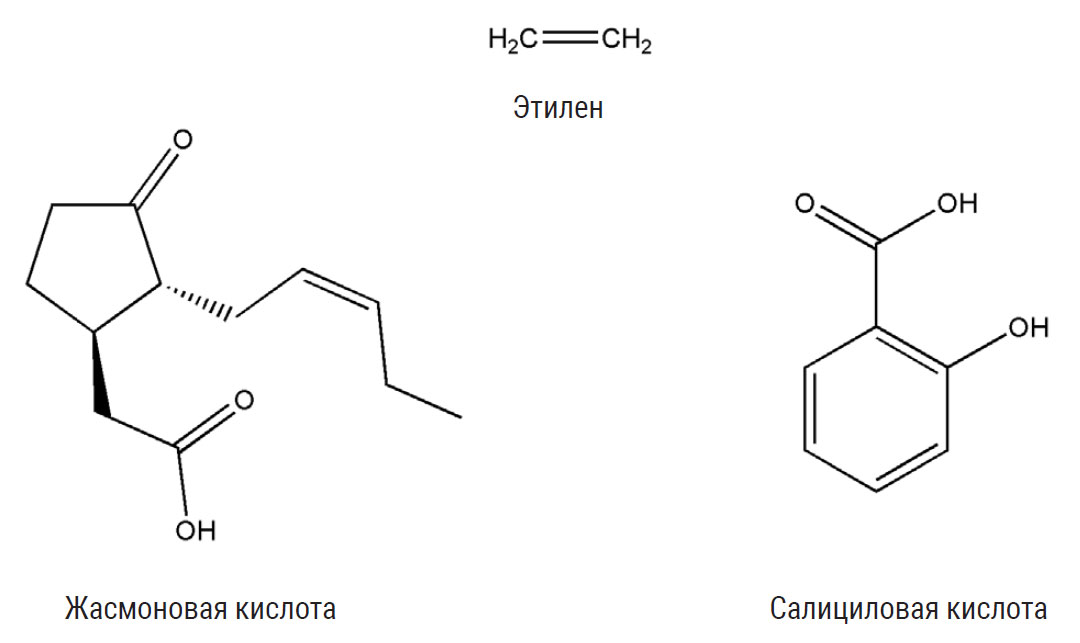
Эти фитогормоны участвуют в защите растений от нападения насекомых-вредителей
Ситуация, когда растение отвечает на атаку травоядных усилением защиты, называют оптимальной индуцируемой защитой. Как правило, она достаточно эффективна. Но есть и другие варианты индуцируемой защиты. Одна из них — индуцированное разнообразие. Суть ее в том, что растение в ответ на сигнал опасности меняет набор синтезируемых веществ. Они не обязательно токсичны, они просто другие — не те, на которые рассчитывает гусеница. У листа появляется непривычный вкус. Причем эти вещества еще и меняются.
Такая чехарда с набором питательных веществ затрудняет гусеницам пищеварение. Так, личинки непарного шелкопряда, выращенные на более разнообразном рационе, растут медленнее и развиваются дольше, чем гусеницы, выращенные на корме столь же питательном, но менее разнообразном. В результате рост численности вредителей замедляется. Такая изменчивость может повредить травоядным насекомым, но не обязательно принесет пользу растению, если вредители съедят изрядную его часть. Индуцированное разнообразие поможет растениям только в случае, если гусеницы умерят свой аппетит. Исследований, посвященных этому механизму, немного, и неясно пока, насколько он распространен в природе.
Третий способ индуцированной защиты — передача информации. Поврежденное растение испускает летучие сигналы, которые побуждают другие, пока еще целые ткани того же растения усилить сопротивление. До соседних растений эти сообщения тоже долетают, и они заблаговременно укрепляют защиту. Но и насекомые эти сигналы улавливают. Для них сигнал означает, что это растение уже занято конкурентами, неплохо защищено и нападать на него не стоит. Эти же сигналы как приглашение воспринимают хищники, которые охотятся на гусениц, и паразитоиды, откладывающие в них яйца. В этом случае эффект защиты зависит от того, какие животные и растения находятся вокруг.
Все три модели предполагают, что растения реагируют на сигналы, указывающие на повышенный риск нападения. Они могут передаваться от клетки к клетке, через сосудистую систему растения, а также соседям по воздуху и через почву, по корням и мицелию микоризных грибов, связывающих растения одного и разных видов. Так, растения фасоли, соединенные гифами грибов с сородичами, зараженными тлей, активно синтезировали репелленты, и тля заселяла их реже, чем не связанные гифами контрольные растения. Они также активно привлекали паразитоидов тли.
Предупрежден — значит вооружен
Мы уже видели, что растения не всегда дожидаются, когда гусеница откусит кусок, и готовятся к обороне, ориентируясь на косвенные сигналы присутствия травоядных, которые еще не едят, но уже явились. Они чувствуют шаги насекомых, сминающих трихомы (тонкие волоски, опушающие растения), реагируют на экскременты насекомых и улиткину слизь. Бабочки-капустницы, откладывая яйца на лист горчицы, пачкают его бензилцианидом. Растение реагирует на бензилцианид и синтезирует вещества, которые могут убить яйца, вырабатывает соединения, ядовитые для гусениц, если они все-таки вылупятся, и в итоге дает больше семян, чем растение, на которое просто посадили гусениц. Такая подготовка защитной реакции называется праймингом.

С помощью этих сигнальных молекул, выделяемых насекомыми, растения распознают присутствие вредителей
Растения также распознают косвенные сигналы, например выделения слюнных желез и отрыжку тех же гусениц, приступивших к пожиранию растения. Они улавливают чавканье насекомых, по-научному — акустические вибрации, вызванные жеванием, и не путают его с шумом ветра или пением насекомых. Растения арабидопсиса, которые перед заражением гусеницами репной белянки Pieris rapae предварительно обрабатывали такой вибрацией, потом синтезировали больше защитных глюкозинолатов, чем необработанные.
С одной стороны, заблаговременное предупреждение об опасности дает возможность как следует к ней подготовиться. С другой стороны, оно, в отличие от несомненного укуса, может и не подтвердиться.
Во многих случаях вредители способны влиять на тревожную сигнализацию растений. Некоторые насекомые переносят микроорганизмы, мешающие передаче растительных сигналов. Например, растения, которыми питается гороховая тля (в данном случае посевные бобы Vicia faba), выделяют летучие вещества, привлекающие паразитических ос, а гороховая тля может переносить бактерии Hamiltonella defensa, которые уменьшают выделение растительных летучих веществ после нападения тли. В результате на зараженное растение слетается меньше ос-паразитоидов. Эти же бактерии предотвращают развитие личинок паразитических ос, чем заметно повышают число выживших тлей.
В других случаях травоядные сами маскируются под патогенные бактерии. Получив ложный сигнал, растения готовятся к защите от инфекции и при этом становятся более чувствительными к вредителям. Кладка яиц бабочки-капустницы Pieris brassicae вызывает в капустных листьях молекулярные изменения, очень похожие на те, которые вызвали бы бактерии. На месте кладки накапливается салициловая кислота, контролирующая защиту от грибков и бактерий. Ее активность мешает активности жасмоновой кислоты, которая имеет решающее значение для защиты от травоядных насекомых. Так сигнальные молекулы яиц обманывают растение.
Растения, в свою очередь, принимают меры, чтобы не стать жертвой «мошенников». Они не изготавливаются к защите после первого же сигнала, обычно их должно быть по крайней мере два подряд. Растения могут хранить информацию в течение нескольких часов. Полынь, например, готовится к сопротивлению, если получает сигналы от поврежденных соседей в течение часа и более. На потоки летучих веществ, испускаемых менее часа, растение не реагирует. Арабидопсис хранит память о неоднократном воздействии слабых сигналов. В каждом из них концентрация летучих веществ была слишком маленькой, чтобы вызвать ответ, но многократное повторение вызывает синтез защитных соединений в листьях.
Память растений обеспечивают несколько факторов, первый из которых — эпигенетические изменения. Это модификации ДНК (обычно присоединение или удаление метильных групп); модификации гистонов — белков, участвующих в укладке ДНК; подавление активности генов под действием малых молекул РНК. Такие изменения могут даже передаваться от материнского растения семенам, попадать во вновь развивающиеся ткани и сохраняться до двух поколений. Это долговременная память. А есть еще память кратковременная, ее обеспечивают накопившиеся фитогормоны или РНК защитного гена, которые не сохраняются долго. Так что в растении, вероятно, одновременно действуют несколько механизмов памяти.
А вот пример того, как кратковременная память влияет на защиту растений. Несколько лет назад Андре Кесслер с коллегами посадил дикий табак рядом с трехзубчатой полынью Artemesia tridentata. У полыни отрезали несколько листочков, и она, будто насекомыми погрызенная, выделяла много биологически активных веществ, в том числе метилжасмонат. В табаке, растущем рядом с такой полынью, метилжасмонат активировал гены защиты от травоядных насекомых. Немедленного эффекта, то есть резкого увеличения количества защитных белков, исследователи не обнаружили. Но позже, когда на табак посадили гусениц бражника, растения вырабатывали ингибиторы трипсиновой протеиназы — пищеварительного фермента гусениц — быстрее, чем табак, не получавший сигналов от полыни. В результате они понесли меньший ущерб и погубили больше гусениц. Почему?
Дело в том, что у табака и полыни почти нет общих травоядных вредителей, а те, что есть, наносят табаку минимальный ущерб. Так что у табака нет оснований бояться полынных гусениц. Однако же сигнал бедствия полыни он улавливает и принимает во внимание. Табак не заготавливает защитные белки впрок, но усиливает синтез РНК защитных генов. И если на табак действительно нападут вредители в тот промежуток времени, пока он «держит в памяти» сигнал тревоги, эти РНК сразу отправятся в рибосомы, и начнется бурный синтез защитных белков. Так растения используют краткосрочный прайминг, чтобы минимизировать и вероятность ложных срабатываний, и затраты на синтез защитных белков, которые могут и не понадобиться.
Еще и конкуренция
Если цель растения заключается в достижении максимальной приспособленности, оно должно стать приспособленнее своих соседей. На первый взгляд, прайминг противоречит этой установке. Передача информации с помощью летучих соединений требует затрат от отправителя. Кроме того, отправитель, предупреждая соседа, создает для него конкурентное преимущество.
На самом деле, это не совсем так. Например, сигналы полыни воспринимают прежде всего ее близкие родственники. Как установил Ричард Карбан с коллегами, летучие вещества, выделяемые поврежденной полынью, сильно различаются у разных растений. Ученые обнаружили, что у генетически родственных особей профили летучих веществ похожи. К тому же популяции полыни организованы так, что растение, скорее всего, будет окружено близкими родственниками. Полынь воспринимает сигналы родичей, а посторонним их «подслушать» труднее.
Иначе ведет себя золотарник высокорослый Solidago altissima. Обычно он образует густые заросли и предупреждает о нападении всю популяцию. В этом случае всё его окружение становится более устойчивым и вероятность повторного проникновения вредителей на этот участок снижается, что принесет несомненную пользу и растению — источнику сигнала.
Растения могут оценить конкуренцию на основании света, отраженного от соседних растений. Хлоропласты преимущественно поглощают красный свет (600–700 нм), поэтому свет, прошедший через лист, сдвинут в сторону дальнего красного (710–800 нм). С помощью фоторецепторов фитохромов растения отслеживают изменения в соотношении красного и дальнего красного света и оценивают плотность растительности вокруг себя.
Если конкурентов много, растения ослабляют конститутивную и индуцированную защиты, но при атаке травоядных выбрасывают больше летучих соединений. Эти соединения привлекают хищных насекомых, что может уменьшить численность вредителей. Так растение избегает относительно дорогой прямой защиты в пользу менее дорогостоящей косвенной и экономит ресурсы в условиях конкуренции.
Цена вопроса — важный фактор, влияющий на выбор защитной стратегии. Принято считать, что индуцированная защита требует меньше затрат, чем постоянная. И концентрация защитного вещества при этом может быть гораздо выше, чем при постоянном синтезе. Однако затраты на синтез и распространение сигнальных молекул бывают так высоки, что превышают затраты на постоянную защиту. Кроме того, эти вещества зачастую отпугивают опылителей, а это плохо. Сравнительное исследование растений семейства Пасленовые показало, что виды, менее зависимые от опылителей, имеют более сильную индуцируемую защиту, в то время как виды, которые в опылителях нуждаются, полагаются на конститутивную защиту.
В общем, индуцируемая защита, оказывается, стоит дорого, она не экономит затраты, а является стратегией, альтернативной конститутивной защите. Выбор защитной стратегии определяется не только экономией, он зависит от стадии жизненного цикла и контекста окружающей среды.
Растения, которые могут сильно пострадать даже от единственной встречи с травоядным насекомым (например, растения на стадии рассады), или обитающие там, где вредителей всегда много, где они малоподвижны и постоянно сидят на растении, или когда нападает несколько видов насекомых сразу, выбирают конститутивную защиту. Если вредителей немного, они мобильны, налетают внезапно и ненадолго, причем каждый вид требует специфического защитного средства, предпочтительнее защита индуцированная, даже если это означает, что большую часть времени растения не будут должным образом защищены от нападающего. Уменьшить затраты на индуцированную защиту помогает правильно организованный прайминг.
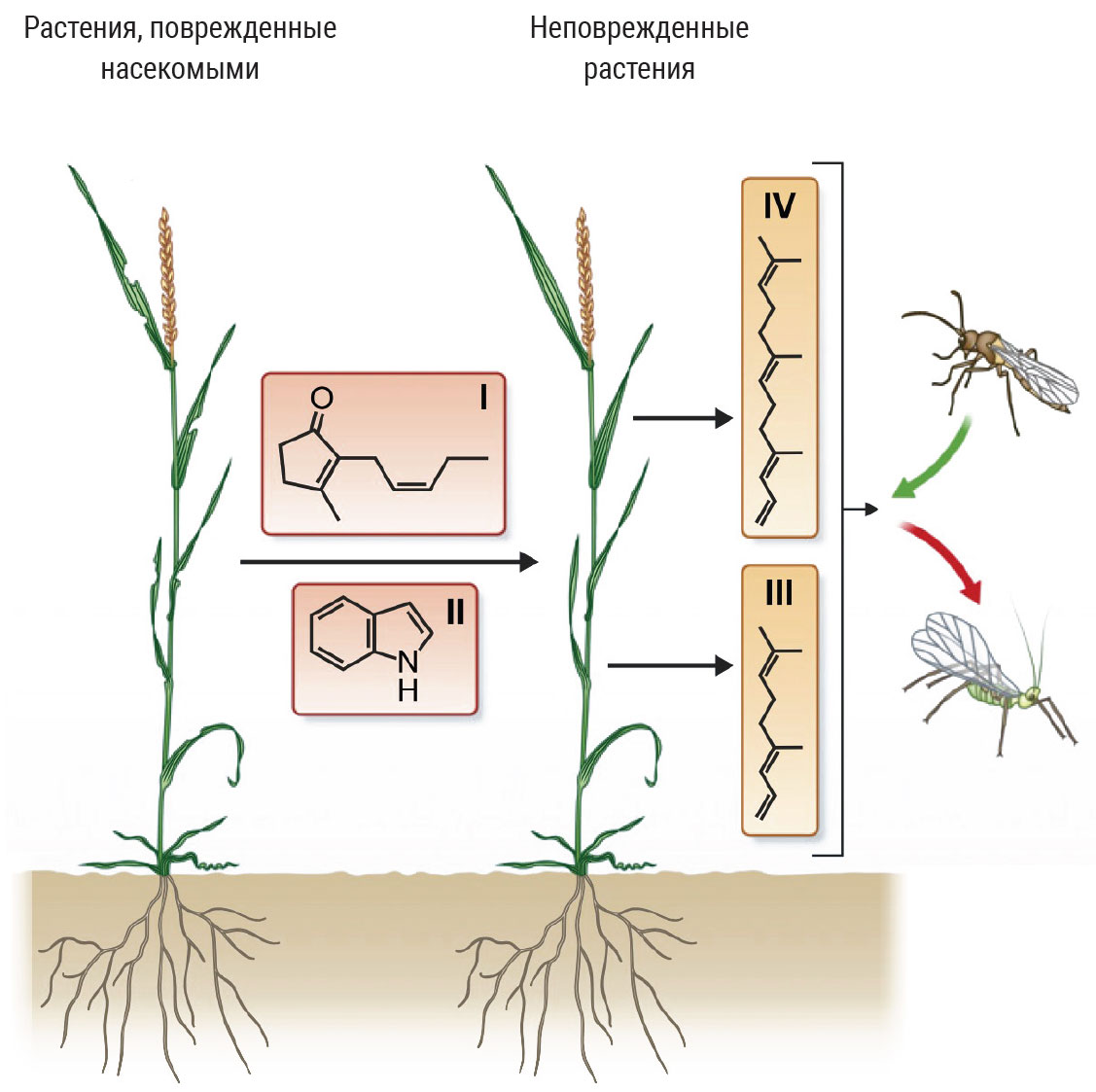
Система защиты растений бывает многоступенчатой. Растения, поврежденные травоядными насекомыми, выделяют стрессовые сигналы I и II. Их воспринимают соседние, еще не тронутые растения, и синтезируют защитные и сигнальные молекулы. Соединения III и IV отпугивают вредителей и в то же время привлекают хищников или паразитоидов, нападающих на этих травоядных. I — цис-жасмон, II — индол, III — (E)-4,8-диметил-1,3,7-нонатриен, IV — (E,E)-4, 8,12-триметил-1,3,7,11-тридекатетраен
Люди пользуются чужим интеллектом
Язык растений сложно расшифровывать. Например, поврежденная полынь выделяет более ста соединений, и понять, какое из них что означает, пока не удается. Информация может зависеть как от комбинации этих веществ, так и от абсолютных и относительных доз. А еще мы не знаем, как растения воспринимают сигналы риска. Есть несколько исключений — рецепторы к этилену и АТФ, вытекающим из поврежденных клеток, и рецепторы к веществам, которые срыгивают гусеницы.
Хотя механизмы защиты растений изучены явно недостаточно, феномен зафиксирован, и его уже успешно используют. Многие африканские фермеры, живущие южнее Сахары, с удовольствием пользуются системой «притяни — оттолкни» (push — pull). Суть заключается в том, что ценную зерновую культуру — кукурузу, сорго или просо — сажают вместе с другим видом, который «отталкивает» вредителей. В этом качестве часто выступает кормовая трава Melinis minutiflora, выделяющая сильный репеллент, отпугивающий бабочек. А чтобы все эти бабочки не полетели откладывать яйца на кукурузу, по краям поля высаживают другие кормовые травы, которые обеспечивают «притяжение». В сумерки, когда бабочки особенно активны, эти травы выделяют гораздо больше привлекательных летучих веществ, чем кукуруза. Пары отталкивающих и притягивающих растений надо специально подбирать в зависимости от зерновой культуры и вредителей, но получается выгодно, потому что травы потом идут на корм скоту. Такая система особенно нравится фермерам-женщинам.
А что нам вообще до интеллекта растений? По мнению исследователей, ценность этой концепции заключается в способности порождать новые проверяемые гипотезы. Она, например, предсказывает, что растения в разных условиях окружающей среды должны по-разному стимулировать и вызывать устойчивость к одним и тем же видам вредителей, в зависимости от относительных затрат, необходимых в той или иной ситуации. Это также означает, что растения могут, находясь в относительно предсказуемой обстановке, действовать по шаблону, а в изменчивых условиях — проявлять поведенческую гибкость. Такой гибкостью прайминга и индуцированной защиты должны отличаться виды, занимающие обширный, разнообразный по условиям ареал. Эти и другие гипотезы еще предстоит проверить.
Согласно определению Шейна Легга и Маркуса Хаттера, интеллект — это способность достигать цели, а все живые существа имеют одну и ту же цель — успешное размножение. Это их объединяет и отличает от искусственного интеллекта, который пока не приобрел стремления к самораспространению, а его цели зависят от желаний человека. Из определения также следует, что интеллект представляет собой фундаментальное свойство жизни, присущее и животным, и растениям, и даже одноклеточным. Нервной системы у них может не быть, а интеллект присутствовать обязан.
-
« Интеллект — это свойство, которым обладает отдельный агент при взаимодействии со своей средой или средами.»
Ну если исходить из этого определения, то кактус действительно имеет интеллект, а пресловутый ИИ под это определение никак не попадает, хотя и может анализировать и сортировать информацию и даже как-бы «обучаться». -
Интеллект — это свойство, которым обладает отдельный агент при взаимодействии со своей средой или средами.
Трудно найти такой "отдельный агент", который при взаимодействии со средой (как определяется из них "своя", по реестру собственников?) не обладал бы хоть каким-нибудь свойством! Получается, что интеллект - это всё что угодно. Например, глупость. При взаимодействии со своей редакционной средой журналов "Химия и жизнь" и "Элементы" она, как выяснилось, обладает свойством публикабельности!
























Иллюстрация Петра Перевезенцева