
 Microtus oeconomus) из рода Серых полевок (Microtus) – один из двенадцати исследуемых автором видов. ">
Microtus oeconomus) из рода Серых полевок (Microtus) – один из двенадцати исследуемых автором видов. ">Гомологическая изменчивость у полевок – результат эпигенетической регуляции.
А.А.Поздняков составил спектры изменчивости первого нижнего моляра у 12 видов полевок. Различия в структуре гомологической изменчивости у разных видов отражает конструктивные особенности жевательной поверхности их зубов. Дополнив внутривидовую гомологическую изменчивость индивидуальной (асимметрия правых и левых зубов у одного зверька), автор обсудил возможность эпигенетического формирования гомологической изменчивости. Индивидуальная асимметрия правой и левой стороны целиком формируется не за счет генетических различий, а за счет онтогенетических вариаций. Фрагменты индивидуальной изменчивости составляются в характерные для вида гомологические ряды, из этого следует эпигенетический механизм формирования гомологических рядов изменчивости нижних моляров.
Даниил Марков • 2 комментария
К статье:

Дрожжи размножаются быстрее, если их попеременно то охлаждать, то нагревать
Температура 30-33°С оптимальна для размножения дрожжей, но только в том случае, если ее поддерживать постоянно. Экспериментально показано, что повышение температуры до сверхоптимальной (37,5-40°С) стимулирует ускоренное размножение дрожжей, если до этого они жили при более низкой температуре (14-30°С). При длительном воздействии повышенной температуры дрожжи теряют терморезистентность, однако способны восстановить ее за всего за одно поколение при 20оС. По-видимому, это свойство является адаптацией к суточным колебаниям температуры в природе. Предполагается, что механизм, позволяющий дрожжам проявлять терморезистентность, локализован в клеточной мембране и его функционирование зависит от энергетической и хемиосмотической систем клеток.







Последние выпуски
Популярные синопсисы

Полиморфизм гена оркокинина OK-A у Apis cerana от тропического Вьетнама до Российской тайги
Рустем Ильясов, Алла Ильясова
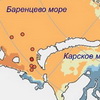
Минтай идет на запад: остановит ли его соленая вода Баренцева моря?
Сергей Баканев

Ритмы роста ствола, ветвей и хвои сосны обыкновенной
Сергей Шавнин, Андрей Монтиле
Мы, зимоспящие млекопитающие, но при этом ещё умеем летать
Виктор Илюха, Светлана Калинина, Екатерина Антонова
