Переизобретение фотосинтеза

Задача
В процессе фотосинтеза фермент рибулозобисфосфаткарбоксилаза, или Рубиско присоединяет к пятиуглеродному сахару молекулу углекислого газа. Скорость этой реакции зависит от концентрации углекислого газа в клетке. У многих цветковых растений возникли дополнительные механизмы повышения концентрации углекислого газа, известные как C4-фотосинтез и CAM-фотосинтез, когда углекислый газ сначала связывается с фосфоенолпируватом, образуя четырехуглеродную (C4) органическую кислоту, а потом в нужное время и в нужном месте четырехуглеродная органическая кислота высвобождает углекислый газ.
Задание 1. Посмотрите на филогенетическое древо (рис. 1), на котором показаны случаи возникновения механизмов повышения концентрации углекислого газа у семенных растений. И C4-, и CAM-варианты фотосинтеза возникали многократно (в частности, насчитывают более 45 случаев независимого возникновения C4-растений; конкретное число отличается в разных исследованиях). При этом в некоторых группах — например, среди злаков или маревых — механизмы концентрации углекислого газа возникают особенно часто.

Как вы думаете, что делает возможным многократное возникновение такого механизма? Почему одни линии могут быть более предрасположены к возникновению C4-фотосинтеза, чем другие?
Задание 2. Одна из главных проблем обычного C3-фотосинтеза — это сродство Рубиско не только к углекислому газу, но и к кислороду. В последнем случае пятиуглеродный сахар окисляется кислородом, что приводит к реакциям фотодыхания. Концентрация кислорода в воздухе гораздо выше концентрации углекислого газа; кроме того, кислород вырабатывается в ходе световых реакций фотосинтеза, поэтому фотодыхание значительно снижает продуктивность фотосинтеза у C3-растений. Почему аналогичной проблемы нет при C4-фиксации углекислого газа? Какие дополнительные адаптации, приводящие к снижению концентрации кислорода в непосредственной близости от Рубиско, есть у C4-растений?
Задание 3. Для того чтобы повысить эффективность сельского хозяйства, было бы здорово перевести C3-культурные растения на C4-путь. Однако это до сих пор не решенная задача. Как вы думаете, какие сложности откладывают ее реализацию?
Подсказка 1
Какой фермент осуществляет первичную фиксацию углекислого газа при C4- или CAM-фотосинтезе? Какие другие функции в клетке известны у этого фермента?
Подсказка 2
В какой форме углекислый газ может находиться в клетке? В какой форме его концентрация больше?
Подсказка 3
Чем отличается строение листа у C4-растений?
Решение
Задание 1
C4- и CAM-фотосинтез — это надстройка над C3-фотосинтезом, все реакции которого сохраняются и в C4-, и в CAM-растениях. Многократное возникновение этой надстройки стало возможно потому, что все белки, задействованные в C4-фиксации углекислого газа, преобразовании и транспорте получившихся органических кислот и расщеплении этих органических кислот с освобождением углекислого газа, уже есть у каждого растения. Они задействованы в других биохимических путях, прежде всего в цикле Кребса, синтезе аминокислот и азотном обмене.

Рис. 2. Функциональная конвергенция в трех основных подтипах C4-фотосинтеза. C4-фотосинтез приводит к повышению концентрации углекислого газа вокруг Рубиско (в центре) в клетках обкладки проводящего пучка. Известны три варианта C4-пути, по разным ферментам, проводящим реакцию декарбоксилирования: NADP-ME (НАДФ+-малатдегидрогеназный тип), NAD-ME (НАД-малатдегидрогеназный тип), PEPCK (ФЕП-карбоксикиназный тип). Цифрами обозначены: 1 — карбоангидраза; 2 — фосфоенолпируват карбоксилаза; 3 — NAD/PH-малат дегидрогеназа, 4.1 — NADP-малик энзим, 4.2 — NAD-малик энзим; 4.3 — фосфоенолпируват карбоксикиназа; 5 — пируват, ортофосфат дикиназа; 6 — аспартат аминотрансфераза; 7 — аланин аминотрансфераза. Хлоропласты обозначены зеленым, митохондрии — оранжевым, клетки обкладки проводящего пучка — светло-желтым, клетки мезофилла — белым. Между мезофиллом и клетками обкладки проводящего пучка есть плазмодесмы (отмечены черным). Сокращения: ALA — аланин, ASP — аспартат, MAL — малат, NAD-ME — NAD-малик-энзим, NADPH-ME — NADPH-малик-энзим, OAA — оксалоацетат, PEP — фосфоенолпируват, PEPCK — фосфоенолпируват карбоксикиназа, PYR — пируват. Рисунок из статьи: B. P. Williams et al., 2012. Molecular evolution of genes recruited into C4 photosynthesis
Переход к C4-фотосинтезу связан с небольшими изменениями в аминокислотных последовательностях основных ферментов, но с существенными изменениями того, когда и где эти белки экспрессируются, и изменениями морфологии листа.
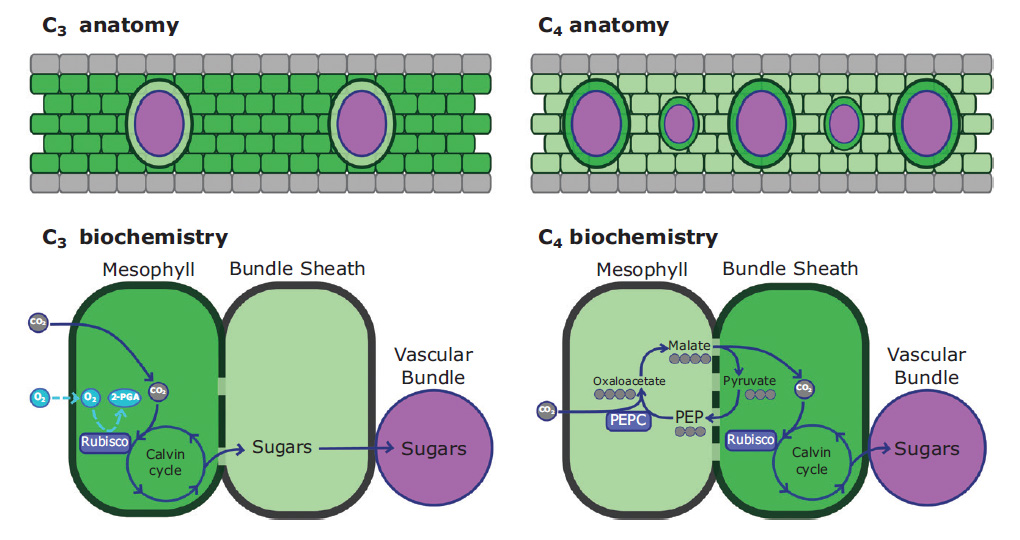
Рис. 3. Схематичное изображение типичной анатомии C3- и C4-листа. Яркий зеленый — клетки с многочисленными хлоропластами; светло-зеленый — клетки, где хлоропластов меньше; фиолетовый — клетки флоэмы; серый — прозрачные клетки эпидермиса листа. В C3-листе больше хлоропластов в клетках мезофилла, клетки обкладки проводящего пучка получают сахара и загружают их во флоэму. В C4-листе в мезофилле происходит лишь первичная фиксация углекислого газа, затем четырехуглеродная кислота транспортируется в клетки обкладки проводящего пучка и там расщепляется с выделением углекислого газа. Углекислый газ входит в цикл Кальвина, таким образом клетки обкладки у C4-растений — основное место синтеза сахаров. У C4-растений меньше объем мезофилла и больше объем клеток обкладки проводящего пучка (и проводящие пучки, то есть жилки, в листе расположены чаще). Изображение из статьи: S. Young et al., 2020. Why is C4 photosynthesis so rare in trees?
C4- и CAM-фотосинтез позволяют более эффективно использовать углекислый газ, а значит, сокращать газообмен через устьица и уменьшать потери воды. Но дополнительные шаги для фиксации и освобождения углекислого газа требуют дополнительных затрат энергии, которую растение получает прежде всего в ходе световых реакций фотосинтеза (поглощения света с последующим транспортом электронов по электронно-транспортной цепи хлоропласта, созданием протонного градиента и использования его для работы АТФ-синтазы). Растения с C4- и CAM-типом фотосинтеза выигрывают в конкуренции у C3-растений в засушливых условиях, при интенсивном освещении и особенно при повышенной температуре, когда C3-растения сильно страдают от фотодыхания. Именно в таких условиях эволюционировал C4- и CAM-фотосинтез — и это отчасти объясняет его неравномерное распределение среди семейств цветковых растений.

Рис. 4. Географические центры происхождения 35 линий двудольных C4-растений, перечисленных в таблице 1. Изображение из статьи: R. F. Sage et al., 2011. The C4 plant lineages of planet Earth. Авторы отмечают, что определить географические центры происхождения однодольных C4-растений сложнее, но похоже, что Южная Америка была одним из основных мест возникновения C4-злаков
Задание 2
Углекислый газ не только растворяется в воде, но еще частично диссоциирует (эту реакцию ускоряют карбоангидразы, см. Carbonic anhydrase). Рубиско взаимодействует именно с молекулярным углекислым газом, а вот ФЕП-карбоксилаза присоединяет к фосфоенолпирувату углекислый газ в форме гидрокарбонат-аниона HCO3–, и, в отличие от Рубиско, не имеет сродства к молекулярному кислороду. При слабощелочном pH в освещенных хлоропластах концентрация гидрокарбонат-иона гораздо выше концентрации растворенного углекислого газа, поэтому, хотя в абсолютных цифрах сродство ФЕП-карбоксилазы к гидрокарбонат-аниону близко к сродству Рубиско к углекислому газу, ФЕП-карбоксилаза фиксирует углерод при более низких концентрациях углекислого газа, чем Рубиско (по Физиологии растений Зитте, 2007).
В большинстве C4-растений ФЕП-карбоксилаза экспрессируется только в клетках мезофилла, а Рубиско — только в клетках обкладки проводящего пучка. Таким образом, они не конкурируют за углекислый газ. Кроме того, у клеток обкладки проводящего пучка C4-растений обычно есть и другие особенности. Например, в них нет или снижено количество фотосистемы II — именно того комплекса, который осуществляет фотолиз воды с образованием кислорода. А их клеточные стенки могут быть пропитаны суберином, который будет препятствовать и диффузии кислорода из мезофилла в клетки обкладки, и диффузии углекислого газа из клеток обкладки.
Задание 3
Переход к C4-фотосинтезу включает в себя множество тонких настроек — изменений морфологии органелл, клеток, листа, а значит, времени, места и количества синтеза всех необходимых для этого белков — не только ферментов, непосредственно вовлеченных в биохимический C4-путь. Пока что нет даже полной картины того, экспрессию каких белков нужно менять, и тем более неизвестно, как менять регуляторные элементы, чтобы получить нужный результат. Известны только отдельные фрагменты этого огромного пазла. В его сборке помогает изучение растений, находящихся на «промежуточных» стадиях между C3- и C4-фотосинтезом.
Послесловие
Насчитывают более 8000 видов растений с C4-фотосинтезом (список по семействам можно посмотреть здесь). Большинство из них относятся к однодольным — злаки и осоки, около четверти — двудольные. C4-растения более полно используют растворенный в клетке углекислый газ, чем C3-растения, и в их органические соединения встраивается больше тяжелого изотопа — C13; это позволяет определить, к какой группе относится растение, не только по особой морфологии листа, которая может быть выражена не очень явно, но и по изотопной подписи. К C4-растениям относятся такие культурные злаки, как сахарный тростник, кукуруза, сорго.
Практически все C4-растения — травы, реже — кустарники и совсем редко — деревья (известные деревья с C4-фотосинтезом относятся к роду молочай — Euphorbia — и, по-видимому, относительно недавно произошли от трав). Выдвигают несколько возможных причин, почему C4-фотосинтез может быть плохо совместим с жизненной формой дерева. Среди них — затенение: для C4-фотосинтеза нужен интенсивный свет, но даже когда дерево растет не в лесу, а на открытом пространстве, то первое время медленного роста его затеняют травы, а потом оно в какой-то степени затеняет само себя. Другая возможная причина — необходимость кормить большой объем нефотосинтезирующих органов: ткани ствола, ткани корней. Клетки обкладки сосудистого пучка у деревьев очень активно загружают флоэму сахарами, и эта роль может быть плохо совместима с их ролью в C4-фотосинтезе; впрочем, пока что это всего лишь предположение, которое надеются уточнить, изучая загрузку флоэмы у древесных C4-молочаев (см. S. Young et al., 2020. Why is C4 photosynthesis so rare in trees?).

Рис. 5. Euphorbia olowaluana, одно из немногих C4-деревьев. Фото с сайта inaturalist.org
Несмотря на глубокое сходство биохимических путей C4-фотосинтеза и, в меньшей степени, морфологии листа C4-растений с разной эволюционной историей, одни и те же внешние признаки могут быть обеспечены разными генами и тем более разными регуляторами. C4-растений так много, что изучение их эволюционных путей далеко не исчерпано. Скорее, можно сказать, что адаптации к C4-фотосинтезу пока изучены частично, прежде всего — для самых интересных объектов.
Одним из таких объектов является род невзрачных сложноцветных Flaveria, в котором, помимо C3-растений, встречаются C4-виды и виды с переходными между C3 и C4 вариантами — у некоторых из них есть механизмы концентрации углекислого газа, выделившегося при фотодыхании, у некоторых есть полный, но не очень активный C4-цикл фиксации и освобождения углекислого газа, а у некоторых C4-цикл активен, но еще не произошло полного разделения функций между клетками мезофилла и клетками обкладки (см. J. Mallmann et al., 2014. The role of photorespiration during the evolution of C4 photosynthesis in the genus Flaveria). Изучение растений из рода Flaveria позволило предположить, что механизмы концентрации углекислого газа, выделяющегося при фотодыхании, в клетках обкладки сосудистого пучка — это преадаптация для возникновения C4-фотосинтеза (подробнее об этом см. здесь и здесь).

Рис. 6. Flaveria trinervia, фото с сайта commons.wikimedia.org
Другой интересный объект — злак Alloteropsis semialata. Некоторые популяции этого злака не способны к C4-фотосинтезу, некоторые обладают промежуточными характеристиками, а некоторые представляют собой полноценные C4-растения. Интересно сравнить разницу в экспрессии генов в листьях между этими тремя вариантами; в исследовании 2019 года обнаружили, что промежуточный C3–C4- и полный C4-фенотипы ассоциированы с повышением экспрессии всего нескольких десятков генов (L.T. Dunning et al., 2019. Key changes in gene expression identified for different stages of C4 evolution in Alloteropsis semialata). Осталось дождаться работ, в которых в разных популяциях Alloteropsis сравнят экспрессию генов в мезофилле и клетках обкладки, и, возможно, мы многое поймем про то, как наименьшими усилиями обеспечить все необходимые морфологические изменения.
Кроме этого, Alloteropsis semialata знаменит 59 фрагментами генома, полученными как минимум от девяти других видов злаков в ходе горизонтального переноса генов (L. T. Dunning et al., 2019. Lateral transfers of large DNA fragments spread functional genes among grasses). Гены в этих фрагментах участвуют в C4-фотосинтезе, формировании устойчивости к заболеваниям и абиотическому стрессу. Как и у других растений, у Alloteropsis semialata уже есть все необходимые для развития C4-фотосинтеза гены, но заимствованные им у C4-злаков гены уже были оптимизированы под C4-фотосинтез и, видимо, поэтому оказались полезны (см. обзор D. A. Wickell, F.-W. Li, 2019. On the evolutionary significance of horizontal gene transfers in plants).
После того как формируется активный C4-цикл и высокая концентрация углекислого газа около Рубиско, становится не так важно, чтобы Рубиско мог работать при низких концентрациях углекислого газа — другими словами, давление отбора на этот признак уменьшается. Оказывается, что в этом случае становится возможно увеличить максимальную скорость фиксации углекислого газа Рубиско. Наблюдаемые при этом две ключевые аминокислотные замены в позициях 291 (аланин → серин) и 309 (метионин → изолейцин) многократно повторяются в разных линиях C4-растений (M. V. Kapralov et al., 2012. Rubisco Evolution in C4 Eudicots: An Analysis of Amaranthaceae Sensu Lato). Такие растения могут синтезировать значительно меньше Рубиско и поэтому более экономно расходовать азот (P.-A. Christin, C. P. Osborne, 2014. The evolutionary ecology of C4 plants). И это еще одна причина думать о том, как перевести C3-культурные растения на C4-рельсы.


Спекулятивно, можно и взять водоросли и перенести их методы фотосинтеза на растение. И будет вам клубника с красными листьями, растущая в подвале.
Улучшение эффективности сельхоз растений нужно, как то надо всех накормить, а фермерством мало кто хочет заниматься. Тот же золотой рис, все боялись, что он страшный ГМО, а теперь продаётся в магазинах. Главное, что чиновники не знают, что он ГМО, а то уберут из полок;)






Рис. 1. Распределение случаев возникновения C4- и CAM-фотосинтеза по филогенетическому древу (по Harris, Davies, 2016) семенных растений. Синие круги обозначают появление C4-фотосинтеза, зеленые — разные варианты CAM-фотосинтеза; диаметр круга пропорционален оценке числа независимых появлений признака в филогенетической линии, собранной по разным исследованиям. Изображение с сайта nph.onlinelibrary.wiley.com