Главный орган головоногих

Задача
Разнообразие и сложность организации и поведения головоногих моллюсков поражает воображение: древние наутилусы с красивыми раковинами, быстрые кальмары, умные осьминоги, загадочные глубоководные вампиротеутисы и спирулы — все они относятся к классу цефалопод. На равных конкурирующие с морскими позвоночными, а кое в чем их и превосходящие, они, тем не менее, являются родственниками медлительных, а то и вовсе неподвижных созданий, ползающих по дну или всю жизнь проводящих в донном иле, — улиток и двустворок.
Головоногие моллюски возникли в позднем кембрии, более полумиллиарда лет назад, и очень быстро стали хозяевами древних морей, заняв самые разные экологические ниши — от активных хищников до гигантских фильтраторов, плавающих в толще воды. Но, судя по палеонтологическим данным, предками головоногих моллюсков были медленные и примитивные «улитки» из класса моноплакофор, населявшие дно кембрийских морей. Они несли на спине широкую, похожую на колпачок раковину и не были способны ни к плаванию, ни вообще к быстрому передвижению. Как эти примитивные и вполне обычные моллюски превратились в стремительных свободно плавающих животных с хорошо развитым мозгом, которых писатель И. Акимушкин прозвал «приматами моря»?
Головоногие моллюски стали такими сложно организованными и высокоразвитыми организмами благодаря эволюции в толще воды — в трехмерном пространстве, полном конкурентов, хищников и добычи. Чтобы начать освоение этого пространства, предкам головоногих нужно было оторваться от дна и научиться плавать. Они сделали это благодаря появлению особого органа, которого нет у других моллюсков. Какой именно орган появился у предков цефалопод в позднем кембрии и стал причиной их удивительного прогресса?
Подсказка 1
Несмотря на то что этот орган фактически «создал» головоногих моллюсков, он сохранился далеко не у всех современных цефалопод.
Подсказка 2
Изучение этого органа у вымерших головоногих является довольно сложной задачей из-за того, что в ископаемом состоянии в основном сохраняются только твердые, минерализованные части тел животных.
Решение
Хотя среди современных головоногих моллюсков есть множество форм, у которых раковина частично или даже полностью редуцирована, а оставшийся от нее рудимент, если он есть, находится внутри тела моллюска, все древнейшие раннепалеозойские головоногие имели наружную раковину. Эта раковина была разделена специальными перегородками (септами) на отдельные отсеки-камеры. Во внешней, самой большой и открытой наружу камере (она называется жилой камерой) находилось тело самого моллюска, а камеры позади него (эту часть раковины объединяют под названием фрагмокон) были заполнены газом и обеспечивали плавучесть всего организма. Среди современных головоногих такая раковина сохранилась только у наутилусов (см. картинку дня Мохнатый наутилус), но в первой половине палеозоя все головоногие моллюски имели наружные раковины, организованные по такому принципу, хотя и очень сильно различавшиеся внешне.
Еще в раннем кембрии, полмиллиарда лет назад, ползавшие по дну кембрийских морей близкие к моноплакофорам примитивные моллюски научились строить перегородки в своих раковинах (см. G. A. Brock, J. Paterson, 2004. A new species of Tannuella (Helcionellida, Mollusca) from the Early Cambrian of South Australia). Но раковина, разделенная перегородками на отдельные отсеки, была лишь первым шагом к появлению головоногих, необходимым, но не достаточным. Сама по себе перегородка лишь отделяет определенный объем раковины от тела моллюска, но ни закачать воду в этот изолированный объем, ни откачать ее оттуда после формирования перегородки уже нельзя (рис. 1). Следовательно, такая раковина хоть и может служить поплавком, но всплывать и погружаться по собственному желанию моллюска она не позволяет.
Для того чтобы разделенная перегородками раковина превратилась в функциональный гидростатический аппарат, нужно было, чтобы возник специальный орган — сифон (англ. Siphuncle). Именно о нем и идет речь в этой задаче.

Сифон — это трубка, проходящая через все отсеки-камеры в раковине головоногого и связывающая их с телом моллюска. Именно появление сифона (рис. 2) позволило древнейшим цефалоподам в позднем кембрии превратить свои раковины в аппарат поддержания нейтральной плавучести, оторваться от дна и начать освоение толщи воды.

Рис. 2. Срез раковины современного наутилуса. Фото с сайта cortesmuseum.com
Позже толщу воды освоили и некоторые гастроподы (например крылоногие моллюски), но они, чтобы сохранять возможность плавать, вынуждены оставаться микроскопическими, в то время как для головоногих были открыты и средний, и даже крупный размерные классы (рис. 3). Необходимость быстро перемещаться и хорошо ориентироваться в трехмерном пространстве, а также возможность вырастать до крупных размеров (известны их ископаемые раковины до 9 метров длиной, см. Гигантские наутилоидеи эндоцериды были мирными фильтраторами?, «Элементы», 02.08.2018) привели к тому, что головоногие моллюски из простых «улиток» постепенно превратились в быстрых и хорошо развитых в интеллектуальном отношении морских обитателей, во многом превзойдя не только всех беспозвоночных, но даже и рыб с амфибиями.

Рис. 3. Гигантские головоногие палеозойской эры: эндоцериды (вверху), ортоцерида (слева) и аммоноидея (справа) в сравнении с человеком. Рисунок из статьи Ch. Klug et al., 2014. Normal giants? Temporal and latitudinal shifts of Palaeozoic marine invertebrate gigantism and global change
Говоря о сифоне, нужно сразу упомянуть, что в русском языке этим термином обозначают еще один орган у моллюсков, что иногда приводит к путанице. Так, у брюхоногих и двустворчатых моллюсков сифон (англ. Siphon) — это мускулистая трубка, соединяющая мантийную полость с внешней средой. И если сифон головоногих находится внутри раковины и представляет собой элемент их гидростатического аппарата, то у двустворок и гастропод он часто очень сильно высовывается из раковины и участвует в дыхании этих животных. У головоногих моллюсков тоже есть трубка, соединяющая мантийную полость с внешней средой, но термин «сифон» специалисты к ней не применяют, а называют воронкой (англ. Hyponome). Хотя воронка и выполняет функции, сходные с функциями сифона двустворок и гастропод (а также участвует в гидрореактивном движении цефалопод), возникла она иначе, чем сифон других моллюсков, и формируется не из тканей мантии, а из того, что у предков головоногих было ногой.
Сифон головоногих моллюсков представляет собой сложно устроенный орган (рис. 4). Он состоит из внешней минерализованной оболочки и расположенных внутри нее мягких тканей: мощных кровеносных сосудов, окружающей их соединительной ткани и наружного эпителия. Внешняя минерализованная часть сифона складывается из специальных выростов в перегородках, через которые проходит сифон (они называются септальными трубками и выполнены из арагонита, как и сами перегородки) и расположенных между ними трубок — соединительных колец (см. картинку дня Атурия).

Рис. 4. Нижнемеловой наутилус Cymatoceras из Краснодарского края и строение его сифона. Цифрой 1 показаны соединительные кольца, 2 — септальные трубки. Фото с сайта ammonit.ru
Соединительные кольца состоят из нескольких слоев пористого карбоната кальция и органического вещества. Их форма очень сильно варьирует в разных группах ископаемых головоногих моллюсков — от цилиндрической, как у современных наутилусов, до почти сферической или напоминающей чечевицу (рис. 5). Также у ископаемых головоногих, судя по всему, сильно варьировало строение стенки соединительных колец, число и структура слоев в ней. Однако тонкие пористые структуры легко растворялись после смерти животного либо замещались различными минералами (иногда не по одному разу), что очень сильно затрудняет изучение их оригинального строения.

Рис. 5. Разнообразие сифонов даже у близкородственных наутилоидей в палеозое было очень велико. На этом фото показано разное строение сифонов у представителей рода Colorthoceras из отряда Orthocerida. У образца внизу строение сифона идентично сифону современных наутилусов, у образца вверху соединительные кольца вздутые, расширенные, в них сформированы внутрисифонные отложения, состоящие из кальцита. Фото из статьи H. Mutvei, R. H. Mapes, 2018. Carboniferous coleoids with mixed coleoid-orthocerid characteristics: a new light on cephalopod evolution
До наших дней дожили только две группы головоногих с функционирующим сифоном классического типа — наутилусы и спирулы. У последних сифон изучен крайне плохо, и о его работе мы не имеем практически никакого представления, а вот функционирование сифона наутилуса изучено детально. Для того чтобы всплыть, наутилус откачивает из камер воду, для того чтобы погрузиться, он, наоборот, закачивает воду в камеры. Для этого наутилус использует осмотическое давление. Обычно в ближайших к телу моллюска камерах фрагмокона находится некоторое количество воды. Когда наутилусу нужно всплыть, одновалентные ионы, главным образом Na+ и Cl− с помощью фермента Na-K-АТФазы, находящегося в сифональном эпителии, откачиваются из камерной жидкости в сифон и далее в кровь. Центральную часть сифона занимают крупные сосуды: четыре артерии, расположенные по краям, и одна мощная вена в середине (см. K. Tanabe et al., 2007. Soft-part anatomy of the siphuncle in Permian prolecanitid ammonoids). Под действием осмотического давления частично опресненная вода тоже поступает в сифон и через почки выводится наружу, а раковина становится легче и наутилус всплывает. Если нужно погрузиться, одновалентные ионы с помощью того же фермента перекачиваются из крови через сифональный эпителий в камерную жидкость, за ними поступает вода, камеры заполняются, плавучесть становится отрицательной (см. К. Н. Несис «Наутилус в аквариуме»). Клетки эпителия, отвечающие за этот процесс, расположены в специальных бороздках на поверхности эпителия (англ. Canaliculi).
У ископаемых головоногих моллюсков сифоны крайне редко сохраняются полностью. Чаще всего хорошо сохраняются септальные трубки, фактически являющиеся частью раковины, несколько реже палеонтологам встречаются соединительные кольца, хотя их находок тоже много. А вот находки фоссилизированных кровеносных сосудов и сифонального эпителия крайне редки — у большинства групп головоногих они неизвестны вообще, а у аммоноидей найдены лишь у двух родов (рис. 6) — пермской Akmilleria из Невады, США (K. Tanabe et al., 2007. Soft-part anatomy of the siphuncle in Permian prolecanitid ammonoids) и юрского Kachpurites из Москвы и Московской области в России (Aleksandr Mironenko, 2017. Siphuncle soft-parts in the Upper Jurassic ammonite Kachpurites fulgens). В обоих случаях мягкие ткани сифонов сохранились благодаря бактериальной фосфатизации в условиях дефицита кислорода в донном осадке и оказались замещены фторапатитом (подробнее об этом процессе у юрских аммонитов см. M. Zatoń, A. Mironenko, 2015. Exceptionally preserved Late Jurassic gastropod egg capsules).

Рис. 6. Фоссилизированные мягкие ткани сифонов аммонитов. Верху — поверхность эпителия пермской Akmilleria electraensis, фото из статьи K. Tanabe et al., 2007. Soft-part anatomy of the siphuncle in Permian prolecanitid ammonoids. Внизу — поперечное сечение тканей сифона юрского Kachpurites fulgens, фото из статьи A. A. Mironenko, 2017. Siphuncle soft-parts in the upper jurassic ammonite Kachpurites fulgens
Строение сифона как пермских, так и юрских аммонитов оказалось очень сходным со строением сифона у наутилусов: на окаменелостях прекрасно видны крупная центральная вена и четыре артерии вокруг нее, губчатая соединительная ткань и даже эпителей (рис. 7). Правда, если у наутилусов количество бороздок в эпителии составляет 240–280, то у пермской Akmilleria их насчитали всего лишь тридцать с небольшим (у юрских Kachpurites, к сожалению, сохранность сифона несколько хуже и число бороздок посчитать невозможно). В принципе, чем больше бороздок, тем больше клеток эпителия и тем быстрее работает сифон, но вполне возможно, что скорость также зависит не только от числа клеток, но и от их строения, которое на ископаемом материале пока изучить невозможно, поэтому сравнивать работу сифонов аммонитов и наутилусов пока преждевременно. Однако очевидно, что общие принципы их функционирования были одинаковыми.
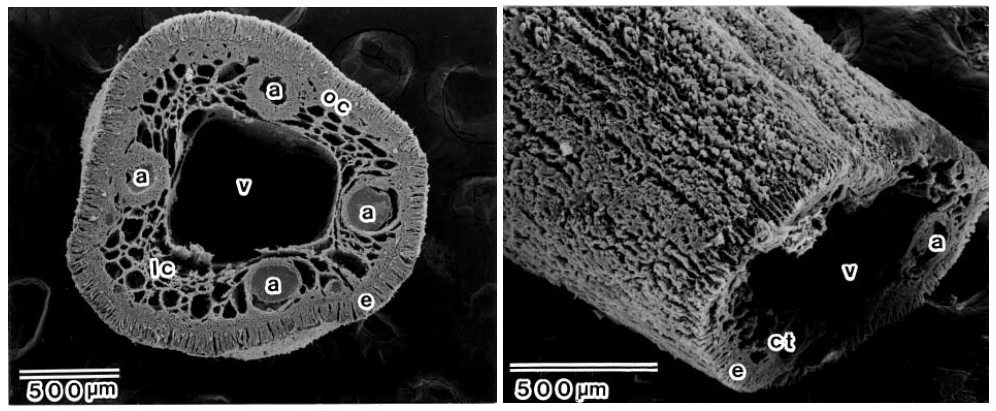
Рис. 7. Строение тканей сифона наутилуса. Видны широкая центральная вена (v), четыре артерии (a), эпителий (e) и бороздки на его поверхности (на правом снимке). Фото из статьи K. Tanabe et al., 2007. Soft-part anatomy of the siphuncle in Permian prolecanitid ammonoids
Впрочем, в этом нет ничего удивительного: и аммониты, и наутилусы унаследовали свой сифон от общих предков — ортоцерид в широком смысле (см. B. Kröger et al., 2011. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules). Однако в первой половине палеозоя существовало множество головоногих моллюсков, сифоны которых были устроены совершенно иначе. Собственно, именно строение сифона лежит в основе выделения большинства таксонов отрядного ранга у наутилоидей. Так, у представителей отрядов Actinocerida, Intejocerida, Discosorida и Oncocerida внутри сифонов формировались очень сложно устроенные минеральные отложения, состоящие из карбоната кальция, строение которых было уникальным для каждого отряда (рис. 8). Продольные ребра, звездообразные разрастания, различные пережимы и перемычки были обычны для внутренней структуры сифонов этих головоногих. Для чего они были нужны и как именно формировались — предмет долгих и пока еще очень далеких от завершения дискуссий палеонтологов.

Рис. 8. Схемы, показывающие внутреннее строение сифонов в отрядах Oncocerida (слева) и Actinocerida (справа). Рисунки из книги J. Dzik, 1984. Phylogeny of the Nautiloidea и статьи I. S. Barskov et al., 2008. Cephalopods in the marine ecosystems of the Paleozoic
Выросты перегородок (септальные трубки) в сифонах древних головоногих также имели очень разную длину: они могли быть как совсем короткими, так и очень длинными, а у некоторых представителей ордовикского отряда Endocerida их длина вдвое превышала длину камеры фрагмокона — то есть трубка, растущая из одной перегородки, заканчивалась на уровне даже не следующей перегородки, а через одну (см. Гигантские наутилоидеи эндоцериды были мирными фильтраторами?, «Элементы», 02.08.2018). У тех же эндоцерид в задней части сифона откладывались кальцитовые конусы — эндоконы. Относительно их функций специалисты более или менее едины в своем мнении: скорее всего эндоконы утяжеляли задний конец раковины, обеспечивая ее горизонтальное положение, а заодно помогали моллюскам избавляться от излишков поступавшего с пищей карбоната кальция. Вряд ли эти эндоконы и упомянутые выше сложные структуры в сифонах формировались исключительно при помощи кровеносных сосудов — скорее всего, в этом участвовал специальный эпителий, аналогичный эпителию мантии, отвечающему за формирование раковины.
Еще необычнее выглядят сифоны некоторых прямораковинных наутилоидей, которых объединяют в отряд Palliocerida (или по-старому Pallioceratida). У этих цефалопод в камерах фрагмоконов встречаются минеральные отложения очень сложной формы, а в стенках соединительных колец у них имеются отверстия, через которые эти отложения соединяются с отложениями в полости сифона. Некоторые палеонтологи считают, что и отложения в камерах, и отверстия в сифонах прижизненные, и что эпителий из сифонов разрастался в камеры, где и формировал эти отложения (см. I. Kolebaba, 1999. Sipho-cameral structures in some Silurian cephalopods from the Barrandian area (Bohemia)). Эту точку зрения неоднократно критиковали и общепризнанной ее назвать сложно, но уж очень сложно устроены отложения в камерах и очень регулярно расположены отверстия в сифонах для того, чтобы можно было считать их посмертными (рис. 9).
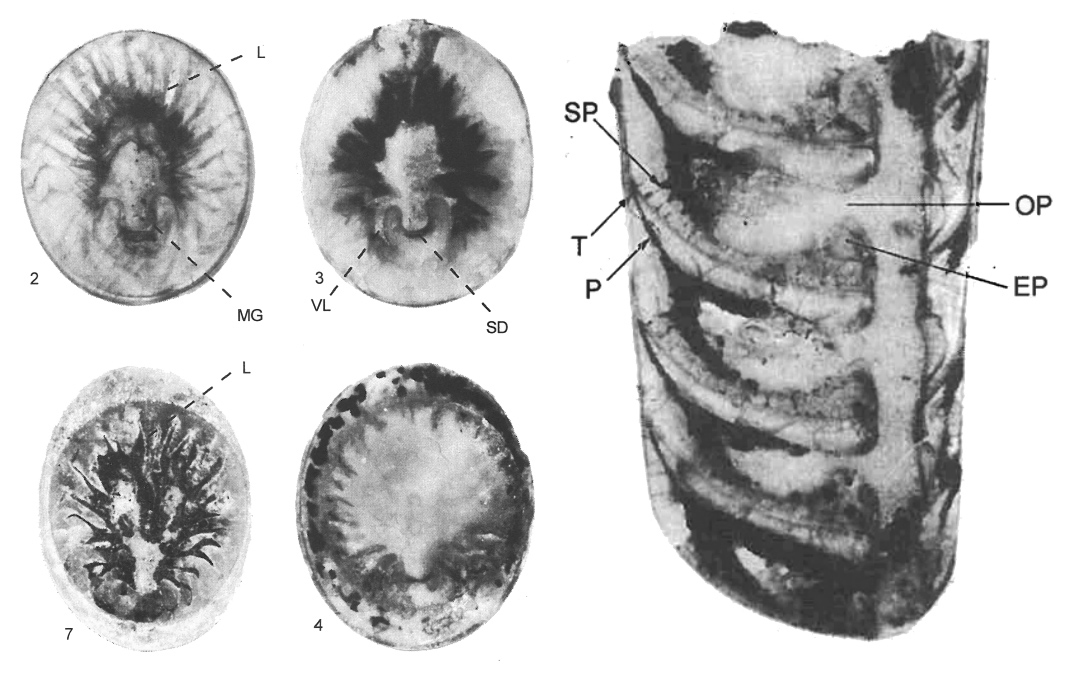
Рис. 9. Поперечные срезы камер фрагмокона (слева) и продольный срез, на котором видны отверстия, соединяющие полости камер с сифоном (справа) в раковинах Palliocerida. Фото из статьи I. Kolebaba, 1999. Sipho-cameral structures in some Silurian cephalopods from the Barrandian area (Bohemia)
О том, как именно функционировали все эти странные и удивительные сифоны, мы можем только догадываться. Ясно только, что они тоже обеспечивали плавучесть раковин своих владельцев, но как именно они работали и были ли они более эффективны, чем сифоны наутилусов и аммонитов, или менее, мы не знаем. Все обладатели таких сифонов вымерли по большей части к середине палеозоя. Далеко не факт, что их вымирание было как-то связано с особенностями строения их сифонов, могло быть множество других причин для их исчезновения. Но, например, в отряде Oncocerida строение сифона неуклонно упрощалось со временем и до второй половины палеозоя дожили лишь формы с очень простым сифоном, почти неотличимым от сифона наутилусов. Значит, что-то полезное в этой изящной простоте было.
Как именно возник сифон у головоногих моллюсков, достоверно не известно, существует множество разных гипотез (см. I. S. Barskov et al., 2008. Cephalopods in the marine ecosystems of the Paleozoic). Так как моноплакофоры с перегородками-септами в раковинах известны уже из низов кембрия, а головоногие появляются только во второй половине этого периода, большинство палеонтологов согласны с тем, что сифон появился у моллюсков, уже умевших строить перегородки внутри раковины. Его возникновение могло быть связано со сбоем в формировании перегородок — задний конец тела моллюска вместо того, чтобы сдвигаться вперед при строительстве каждой новой перегородки, почему-то оказывался намертво прикреплен к апикальной части раковины, и перегородки строились вокруг него. Однако интересно, что уже в кембрии выделились две ветви головоногих — одна с узкими и простыми сифонами, другая — с широкими и сложно устроенными. Представители обеих ветвей сумели пережить великое вымирание в самом конце кембрия, уничтожившее почти 95% существовавших тогда родов головоногих, и в ордовике дали начало всему тому разнообразию, о котором говорилось выше (см. X. Fang et al., 2018. Palaeogeographic distribution and diversity of cephalopods during the Cambrian–Ordovician transition).
Послесловие
Несмотря на то что головоногие моллюски в принципе возникли благодаря появлению сифона, две процветающие в современных морях ветви цефалопод (кальмары и осьминоги) полностью утратили его в ходе своей эволюции. Сифон у них исчез вместе с фрагмоконом, а раковина превратилась в тонкую органическую пластинку — гладиус, задняя часть которого распалась у некоторых осьминогов на две тонкие «веточки», именуемые стилетами. Наиболее вероятным объяснением отказа от фрагмокона и, как следствие, от сифона является развитие мускульной мантии, обеспечивающей гидрореактивное движение головоногих (рис. 10; см. В. А. Бизиков, 2008. Эволюция раковины головоногих моллюсков).

Рис. 10. Гладиусы современных кальмаров (вверху), вампиротеутиса (внизу слева) и осьминогов (внизу в центре и справа). Все эти внутренние раковины функционируют только как опорный скелет, фрагмокон и сифон у них отсутствуют. Рисунок из статьи В. А. Бизиков, 2008. Эволюция раковины головоногих моллюсков
Большинство современных головоногих моллюсков плавают, с силой выталкивая воду из мантийной полости через мускулистую трубку — воронку (как раз ту, которая похожа на сифон двустворок и гастропод, но им не является). Конечно, есть и множество других способов локомоции — с помощью плавников или растянутой между щупальцами перепонки-умбреллы, но гидрореактивное движение — самые быстрое, оно позволяет сбежать от хищника или догнать добычу.
Однако, выталкивать воду из мантийной полости можно по-разному. Наутилусы для этого втягивают голову в раковину, и она работает как поршень, выталкивая воду. Раньше этот способ экстраполировали на всех вымерших наружнораковинных головоногих (см. В. А. Бизиков, 2008. Эволюция раковины головоногих моллюсков). Однако для такого плавания нужны очень мощные мускулы-ретракторы головы, места прикрепления которых к раковине прекрасно видны даже в ископаемом состоянии. Но ни у кого из ископаемых цефалопод, кроме прямых предков современных наутилусов, таких мест прикрепления мускулатуры нет, а значит, и плавали они как-то иначе. Возможно, что основным двигателем древних наутилоидей была не мантийная полость, а воронка — именно ее сокращения выталкивали воду (см. W. B. Saunders, P. D. Ward, 1994. Nautilus is not a model for the function and behavior of ammonoids). У современных колеоидей воронка тоже способна сжиматься, но эти пульсации играют вспомогательную роль по сравнению со сжатием мантийной полости. Объем мантийной полости в любом случае больше объема даже самой толстой воронки. Поэтому у белемнитов и некоторых других мезозойских колеоидей стенки жилой камеры снизу и с боков исчезли, их заместила мускульная мантия, позволявшая выжимать воду из мантийной полости (рис. 11).

Рис. 11. Схематичное строение белемнита. Красным показана раковина (фрагмокон с сифоном) и покрывающий ее проостракум, выступающий вперед на спинной стороне, синим — кальцитовый ростр. Видно, что в передней и брюшной части раковина отсутствует, там расположена мускульная мантия. Рисунок из статьи R. Hoffmann, K. Stevens, 2019. The palaeobiology of belemnites — foundation for the interpretation of rostrum geochemistry
После возникновения мускульной мантии минерализованный фрагмокон внезапно стал помехой на пути дальнейшего развития этого механизма плавания: он и занимал часть внутреннего объема тела, за счет которого можно было бы увеличить мантийную полость, и был хрупким, что не позволяло мощной мускулатуре развиваться в непосредственной близости от него. Уменьшение его размеров и в конце концов полный отказ от него стал выгоден с точки зрения развития нового способа быстрого плавания. Точной уверенности в этом пока нет, но похоже, что фрагмокон и сифон неоднократно независимо исчезали в разных эволюционных линиях колеоидей в мезозое.

Рис. 12. Расположение внутренних раковин (показаны синим) в телах современных головоногих. Верхний ряд: Spirula и Sepia, их раковины сохраняют функцию обеспечения нейтральной плавучести. Нижний ряд — кальмар (Chtenopteryx) и осьминог (Octopus), их раковины являются только опорными элементами скелета. Рисунки В. А. Бизикова из статьи V. A. Bizikov, R. B. Toll, 2016. The Gladius and its Vestiges in Extant Coleoidea
Конечно, после отказа от фрагмокона кальмарам и осьминогам пришлось решать проблему с обеспечением нейтральной плавучести. Тут в разных эволюционных линиях возникли разные решения. Глубоководные кальмары и осьминоги пошли по пути обводнения тканей и использования плавников, которые, постоянно находясь в движении, поддерживают их на плаву. Мелководные кальмары научились запасать в своих мышечных тканях пузырьки аммиака, который легче воды и позволяет им всегда сохранять нейтральную плавучесть. Осьминоги (отряда Incirrata), живущие среди скал и камней на относительно небольших глубинах, вообще стали тяжелее воды — в случае необходимости они могут плавать на своем гидрореактивном двигателе, но предпочитают неспешно перемещаться по дну. А их потомки из семейства Argonautoidea, вернувшиеся в толщу воды, были вынуждены изобретать аппарат нейтральной плавучести заново — у рода Argonauta им стала раковина с пузырьком воздуха в верхней части, а представители близкого к ним рода Ocythoe и вовсе разработали аналог плавательного пузыря рыб (см. картинку дня Осьминог в раковине).
Однако не все колеоидеи утратили фрагмокон и сифон. Полностью функциональный гидростатический аппарат сохранили спирулы (см. картинку дня Удивительная спирула), ушедшие в мезопелагиаль, туда, где меньше конкуренция и можно плавать не слишком быстро (рис. 13, слева). Но необычнее всего поступили каракатицы (отряд Sepiida). Многие знакомы с их внутренними раковинами — овальными белыми пластинками, которые называют сепионом или «костью каракатицы» (англ. Cuttlebone). Их часто продают в зоомагазинах в качестве источника кальция для домашних птиц и рептилий. Эта белая пластинка представляет собой сильно видоизмененный фрагмокон, у которого совершенно редуцировалась вентральная (нижняя) сторона (рис. 13, справа). При этом редуцировалась и нижняя сторона сифона, а сам сифон из замкнутой трубки превратился в широкую плоскую область в задней части раковины, которая выделяется хорошо заметными линиями нарастания (см., например, A. G. Checa et al., 2015. The cuttlefish Sepia officinalis (Sepiidae, Cephalopoda) constructs cuttlebone from a liquid-crystal precursor).

Рис. 13. Слева — сифон в раковине современной Spirula (показан красным). Справа — раковина каракатицы Sepia с сифонной областью (sz). Рисунок спирулы из статьи R. Hoffmann, K. M. Warnke, 2014. Spirula — das unbekannte Wesen aus der Tiefsee, фото сепии из статьи A. G. Checa et al., 2015. The cuttlefish Sepia officinalis (Sepiidae, Cephalopoda) constructs cuttlebone from a liquid-crystal precursor
Подобная радикальная перестройка фрагмокона позволила существенно увеличить площадь сифона и скорость его работы. Этим каракатицы убили сразу не двух, а трех зайцев: сохранили работающий аппарат нейтральной плавучести, сделали его компактным и прочным и переместили на спинную сторону, что позволило развить и мускульную мантию, и большую мантийную полость, а к тому же за счет широкого плоского сифона получили возможность почти мгновенно менять плавучесть раковины и даже точно настраивать балансировку, быстро утяжеляя или облегчая задний конец раковины (см. В. А. Бизиков, 2008. Эволюция раковины головоногих моллюсков).
Поэтому пока еще точку в истории сифона головоногих ставить рано. Хотя обладатели «классического» сифона — наутилусы и спирулы — не слишком многочисленны и разнообразны в современных морях, каракатицы со своей «продвинутой» версией сифона процветают. И кто знает, какие еще эволюционные модификации сифонов могут появиться у головоногих в будущем?






Рис. 1. Tannuella elinorae из нижнекембрийских отложений, Австралия. У этих моллюсков были перегородки в раковинах, но сифона не было. Фото из статьи G. A. Brock, J. Paterson, 2004. A new species of Tannuella (Helcionellida, Mollusca) from the Early Cambrian of South Australia