Гетероплазмия

Задача
У эукариот часть наследственной информации закодирована в геномах органелл — митохондрий и пластид. Так, например, митохондрии содержат в себе молекулы митохондриальной ДНК* (мтДНК): на каждую клетку обычно приходится 50–100 000 молекул мтДНК. Если в отдельно взятой клетке содержится два или более разных вариантов мтДНК, то говорят, что она находится в состоянии гетероплазмии (рис. 1).

Гетероплазмия обычно возникает в результате мутации одной из множества молекул мтДНК. Даже в покоящихся клетках происходит круговорот молекул мтДНК: постепенно синтезируются (реплицируются) новые молекулы, а их избыток разрушается. Поэтому существует вероятность того, что мутантный вариант увеличит свою относительную частоту по сравнению с исходным вариантом и даже «обгонит» его.
Однако такое состояние гетероплазмии неустойчиво: в череде поколений частоты исходного и мутантного вариантов мтДНК меняются, так как митохондрии расходятся в две клетки случайным образом (рис. 2).

Рис. 2. Из-за случайного расхождения митохондрий многие клетки-потомки в ряду поколений теряют один из вариантов мтДНК и переходят в состояние гомоплазмии
Рано или поздно в процессе деления одна из клеток получает только один вариант мтДНК и тем самым переходит в состояние гомоплазмии. Это состояние практически необратимо. У большинства эукариот мтДНК наследуется строго по материнской линии, и получить утерянный вариант мтДНК неоткуда, а вероятность того, что произойдет точно такая же мутация в оставшемся варианте мтДНК, крайне мала.
У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Если в клетках зародышевой линии возникла гетероплазмия, то она может передаваться по наследству от матери потомкам. Однако на одно поколение животных приходится несколько десятков делений клеток зародышевой линии (у самок). Поэтому обычно частоты мтДНК от поколения к поколению сильно меняются или один из вариантов мтДНК теряется. Однако описаны случаи, когда гетероплазмия мтДНК сохранялась в череде нескольких поколений на приблизительно постоянном уровне и практически не приводила к выщеплению потомства только с одним из двух вариантов мтДНК.
Как вы думаете, что могло препятствовать переходу в состояние гомоплазмии (потере одного или второго варианта мтДНК) в этих случаях?
* Впрочем, бывают и исключения: есть множество вариантов дифференцированных клеток многоклеточных животных, лишенных мтДНК (пример — эритроциты млекопитающих). Кроме того, у некоторых видов эукариот в процессе эволюции митохондрии сильно редуцировались и мтДНК была утеряна (микроспоридии и др.).
Подсказка 1
В некоторых описанных случаях один из вариантов мтДНК был «неполноценный», то есть не содержал в себе всей необходимой информации, которая необходима для выполнения митохондриями своих функций.
Подсказка 2
Как правило, вредные мутации в митохондриальной ДНК проявляются на уровне целой клетки, только если относительная доля этих мутантных вариантов мтДНК превышает некий порог (обычно 60–80%). Если же доля мутантных вариантов мтДНК ниже этого порога, то клетка живет как ни в чем не бывало.
Решение
Первым, на первый взгляд очевидным, способом поддержания гетероплазмии в череде поколений является наличие двух взаимодополняющих мутаций в одном и в другом варианте мтДНК. Например, если в одной из молекул мтДНК произошла мутация, «испортившая» ген цитохром-оксидазы (COX1), а во второй — ген NADH-дегидрогеназы (ND1), то выживут только те клетки, в которых есть оба варианта мтДНК. Они смогут синтезировать необходимый для своего функционирования набор митохондриальных белков (рис. 3). Митохондрии в клетках активно делятся и сливаются друг с другом, а их содержимое перемешивается, поэтому возможность такого взаимного дополнения теоретически существует. Те же клетки, которые потеряли один из вариантов, — погибнут. Цитохром-оксидаза и NADH-дегидрогеназа нужны для клеточного дыхания, в их отсутствие клетки неспособны к размножению и быстро теряют жизнеспособность.
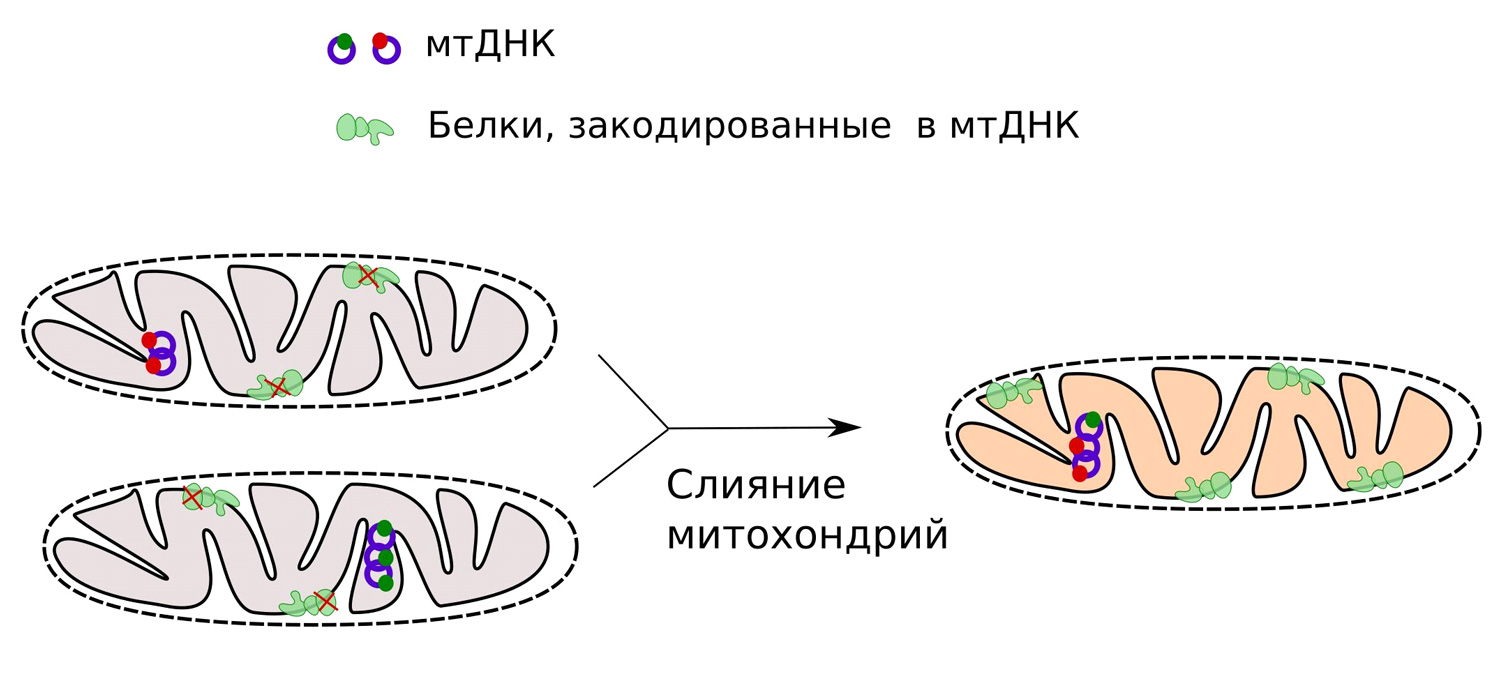
Рис. 3. Слияние митохондрий с мутациями в разных генах (обозначены как синяя и зеленая точки на молекулах мтДНК) теоретически может дать возможность синтезировать полный набор белков, необходимых для выполнения митохондрией ее важнейшей функции — окислительного фосфорилирования
Однако реальных примеров такой взаимной комплементации (автору задачи) неизвестно. Более того, в исследованиях, где клетки с мутациями в разных генах мтДНК искусственно заставляли сливаться, получались цибриды с нефункциональными митохондриями (D. Takai et al., 1999. Transcomplementation between Different Types of Respiration-deficient Mitochondria with Different Pathogenic Mutant Mitochondrial DNAs). Цибридами (или цитоплазматическими гибридами) называют клетки, полученные при слиянии двух клеток, если ядро одной из двух исходных клеток было утеряно, а цитоплазма осталась общей. По всей видимости, на практике неполноценные молекулы мтДНК с вредными мутациями в разных генах далеко не всегда могут дополнять друг друга.
Есть еще одна причина, по которой описанный выше вариант маловероятен. Как два дополняющих друг друга варианта мтДНК могли бы оказаться в одной и той же клетке зародышевой линии? Гаметы, которые являются клетками зародышевой линии, сливаются и образуют зиготы. Если бы гаметы содержали два разных мутантных варианта мтДНК, то оба они оказались бы в зиготе. Однако если каждый из вариантов мтДНК неполноценен (не содержит в себе полной информации, необходимой для работы митохондрий), то скорее всего гаметы, содержащие такую мтДНК, были бы неполноценными. Кроме того, для многих видов показано, что отцовская митохондриальная ДНК, попавшая в зиготу из мужской гаметы, активно уничтожается — переваривается вместе с отцовскими митохондриями в аутофагосомах.
Случаи стабильной гетероплазмии описаны для различных групп эукариот. Стабильная гетероплазмия показана для пекарских дрожжей (одноклеточные грибы) и растений. У нематод Caenorhabditis elegans и C. briggsae также известны линии со стабильной гетероплазмией. Так, например, описана линия C. elegans, в которых гетероплазмия поддерживалась на протяжении по крайней мере ста поколений: в нематодах обнаруживали один полноразмерный вариант мтДНК, второй вариант мтДНК содержал делецию одиннадцати генов (W. Y. Tsang and B. D. Lemire, 2002. Stable heteroplasmy but differential inheritance of a large mitochondrial DNA deletion in nematodes). Недавно устойчивая гетероплазмия была обнаружена в популяции обыкновенных скворцов, интродуцированных на территорию Австралии. Передача гетероплазмии из поколения в поколение описана и в случае человека.
Как предполагается, случаи стабильной гетероплазмии связаны с тем, что селекция митохондриальной ДНК происходит сразу на нескольких уровнях (рис. 4).

Рис 4. Естественный отбор мтДНК может проходить на уровне индивидуумов в популяции, клеток в организме, митохондрий в клетке и мтДНК в митохондриях. При этом некоторые варианты мтДНК могут иметь увеличенный шанс распространения в клетке, но вредить на уровне целых клеток или индивидуумов. Рисунок из статьи: L. A. Rollins et al., 2016. Selection on Mitochondrial Variants Occurs between and within Individuals in an Expanding Invasion
С одной стороны, разные варианты митохондриальных геномов «конкурируют» на уровне организма: плохие варианты (мутации в белок-кодирующих генах) обычно приводят к миопатиям или неврологическим проблемам и быстро отсеиваются естественным отбором. С другой стороны, разные варианты мтДНК конкурируют и внутри клетки — так, например, короткие митохондриальные ДНК реплицируются быстрее, чем длинные, и постепенно вытесняют их (рис. 5). В результате может возникнуть баланс: один из вариантов мтДНК вытесняет другой внутри клетки, но вредит на уровне целых клеток (или целых организмов).

Рис. 5. Разнонаправленная селекция мтДНК на уровне отдельных молекул и на уровне целых клеток приводит к стабилизации гетероплазмии в череде поколений. Мутантная мтДНК (а, обозначена красным цветом) быстрее реплицируется в клетках (например, за счет того, что в ней произошла делеция и нужно меньше времени, чтобы завершить репликацию). Поэтому в череде поколений она постепенно увеличивает свою относительную частоту по сравнению с полноразмерной мтДНК (b, обозначено синим цветов). Однако если мтДНК b становится слишком мало, то такая клетка прекращает деление или погибает. Поэтому клетки, в которых больше мтДНК b, делятся быстрее, чем клетки, в которых мтДНК b мало.
Послесловие

Рис. 6. Митохондрии фибробласта кожи человека образуют сеть. Фотография сделана с помощью флуоресцентного микроскопа, для визуализации митохондрий к клеткам добавляли родамин 123 (см. Rhodamine 123) — флуоресцентное соединение, которое накапливается в митохондриях. Фото из статьи: J. Hayashi et al., 1994. Human mitochondria and mitochondrial genome function as a single dynamic cellular unit
В условиях нашей задачи было сказано, что в клетках эукариот содержится 50–100 000 молекул мтДНК. Но как эти мтДНК распределены по митохондриям? Сколько молекул мтДНК содержится в каждой отдельной митохондрии и сколько отдельных митохондрий обычно можно обнаружить в клетке? На эти вопросы нет прямого (количественного) ответа. Это связано с тем, что митохондрии в клетках постоянно делятся и сливаются друг с другом, а во многих случаях вообще образуют единую сеть (рис. 6). Такую сеть иногда называют митохондриальным ретикулумом. Однако даже в тех клетках, в которых митохондрии представлены разрозненными органеллами, в течение времени их содержимое перемешивается. Поэтому мтДНК в клетке имеет смысл рассматривать только вместе, как совокупность, вне зависимости от того, как они распределены по отдельным органеллам (см. J. Hayashi et al., 1994. Human mitochondria and mitochondrial genome function as a single dynamic cellular unit).
Наличие множества молекул мтДНК в клетках приводит к «трагедии общин» (см. D. Haig, 2016. Intracellular evolution of mitochondrial DNA (mtDNA) and the tragedy of the cytoplasmic commons). Получается, что каждая отдельно взятая мтДНК практически ни на что не влияет. Появление одной мтДНК с вредной мутацией никак не отразится на эффективности работы митохондрий. В большинстве случаев клетка «не чувствует» до 60% дефектных мтДНК (рис. 7). Под дефектными мтДНК мы подразумеваем такие мтДНК, в которых изменения нуклеотидной последовательности не позволяют синтезировать весь необходимый набор белков, необходимых для клеточного дыхания. Превышение этого «порога патогенности» довольно быстро приводит к нарушениям в работе клеток, а если таких клеток оказалось много — к патологиям многоклеточного организма.
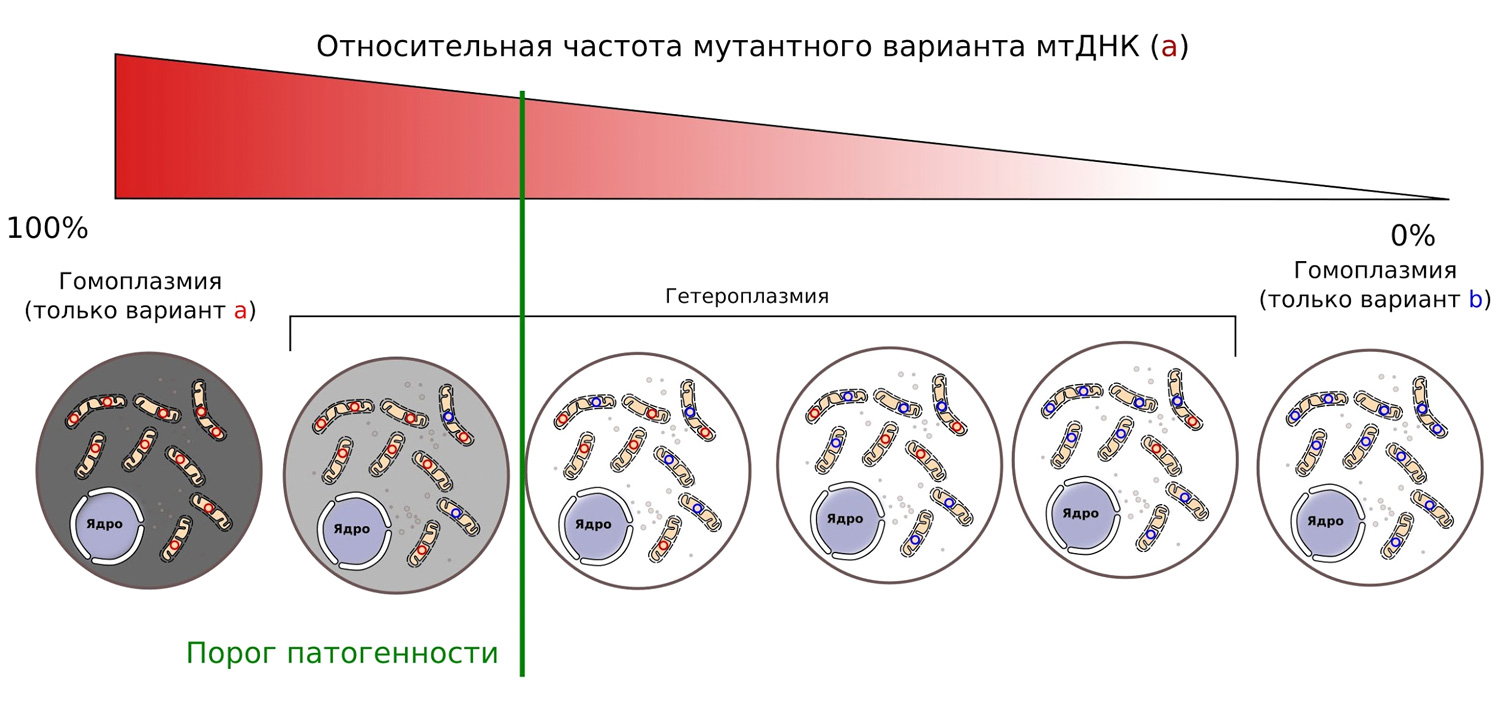
Рис. 7. Увеличение относительной частоты мтДНК в мутации начинает проявляться только при превышении определенного порога (своего для каждого типа мутаций)
Эти два условия — 1) «невидимость» для клетки мтДНК с вредными мутациями до тех пор, пока их доля относительно нормальной мтДНК не превысит определенный порог, и 2) случайная сегрегация (расхождение) мтДНК в процессе деления клеток — порождают для многоклеточных существ проблему. Даже незначительная доля мтДНК с вредной мутацией (ниже порога патогенности) в зиготе может привести к нарушениям у взрослого многоклеточного существа, которое разовьется из этой зиготы. В процессе развития многоклеточного организма количество клеток многократно увеличится. Так, например, человек состоит из приблизительно 3·1013 клеток (R. Sender et al., 2016. Revised Estimates for the Number of Human and Bacteria Cells in the Body). Одни клетки получают чуть меньше, а другие чуть больше этих «неблагополучных» молекул мтДНК. Рано или поздно могут появиться клетки, в которых доля молекул мтДНК с вредными мутациями приблизительно равна порогу токсичности. При делении таких клеток с большой вероятностью будут появляться клетки, в которых доля мутантных мтДНК уже превышает порог. Такие клетки не смогут выполнять свои функции, и если таких клеток окажется много, то ткань или орган многоклеточного организма не будет справляться со своей работой.
Варианты мтДНК, которые реплицируются внутри клетки быстрее обычной мтДНК, но несут в себе вредные мутации (см. рис. 5 в решении), тем более опасны для многоклеточного организма. По мере деления клеток относительная доля таких мутаций будет постепенно возрастать, и порог патогенности будет достигнут раньше. Такие варианты мтДНК называют «эгоистичными мтДНК». Они имеют преимущество в репликации, но не несут никакой пользы (а чаще даже несут вред) для клетки или для всего организма. Предполагается, что уничтожение отцовской митохондриальной ДНК в зиготе, о котором мы упоминали выше, необходимо как раз для того, чтобы защищаться от «заражения» зародышевой линии такими вариантами мтДНК. Те немногие виды, у которых мтДНК наследуется от обоих родителей, сталкиваются с большими проблемами, если в них появляется эгоистичная мтДНК. Примером эукариот, которые получают мтДНК от обоих родителей, являются пекарские дрожжи. В них описаны мтДНК, которые вообще не несут никакой полезной информации, но в подавляющем большинстве случаев (~99 из 100) полностью замещают собой полноразмерную мтДНК, содержащую необходимую наследственную информацию (см. I. E. Karavaeva et al., 2017. Mitochondrial depolarization in yeast zygotes inhibits clonal expansion of selfish mtDNA).
Существует большое число заболеваний человека, связанных с мутациями в митохондриальной ДНК. Многие из этих заболеваний наследственные. Поскольку у человека митохондриальная ДНК наследуется по материнской линии, то и заболевания, связанные с мутациями в мтДНК, также передаются от матери — детям. В большинстве случаев эти ассоциированные с заболеваниями варианты мтДНК находятся в состоянии гетероплазмии: одновременно с мутантной мтДНК в клетках содержится и мтДНК без вредных мутаций. Чаще всего такие митохондриальные заболевания затрагивают те ткани, в которых наиболее важна роль митохондрий: мышцы и нервные ткани. Одним из наиболее известных и распространенных митохондриальных заболеваний является нейродегенеративное заболевание — синдром MELAS. Наиболее распространенная форма MELAS связана с мутацией в одном из генов мтДНК — транспортной РНК лейцина. Этот ген кодирует транспортную РНК, которая необходима для синтеза закодированных в митохондриальной ДНК белков. Замена одного из нуклеотидов в этой РНК приводит к нарушению ее функции и большим проблемам в работе митохондрий.
Принципы сосуществования разных мтДНК внутри клетки (гетероплазмии) до сих пор не до конца ясны. Остается довольно много открытых вопросов. Не до конца понятна природа порога патогенности, далеко не всегда прослежена цепочка причинно-следственных связей между мутациями в мтДНК и нарушениями функций тканей и органов, также неизвестно, какие мутации дают молекулам мтДНК преимущества в скорости репликации, а какие нет. Однако уже сейчас ясно, что гетероплазмия очень широко распространена в природе и оказывает большое влияние на жизнь и эволюцию живых организмов.






Рис. 1. Схематическое изображение эукариотической клетки в состоянии гетероплазмии (слева) и гомоплазмии (справа)