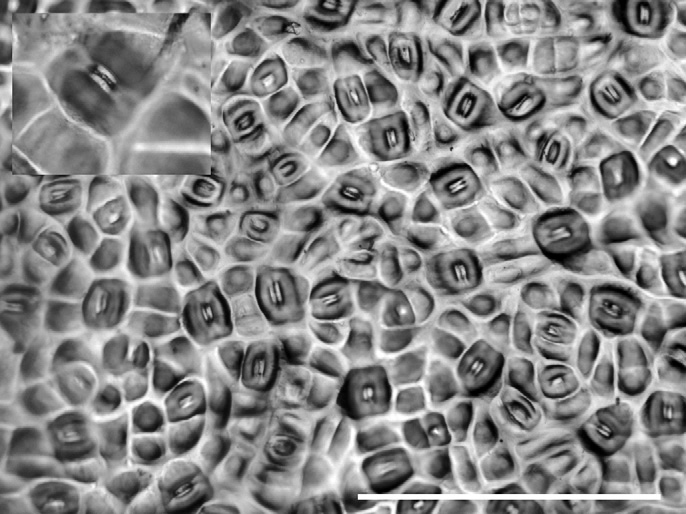
Устьица омелы

Листья и молодые стебли высших растений покрыты особой тканью — эпидермой. Почти все клетки эпидермы прозрачны. Они плотно прилегают друг к другу и выделяют на поверхность листа слой воскоподобных веществ — кутикулу. Кутикула эпидермы защищает лист от высыхания, но при этом препятствует газообмену с атмосферой.
Для фотосинтеза растению необходим углекислый газ, поэтому без газообмена не обойтись, и в эпидерме листа есть поры — устьица, состоящие из пары бобовидных клеток и щели между ними. Эти клетки могут менять свою форму, открывая или закрывая устьичную щель. Клетки толщи листа под устьицами располагаются очень рыхло, образуя микропустоты — межклетники. Воздух в межклетниках насыщен парами воды.
Открывая или закрывая устьичную щель, растение может регулировать интенсивность газообмена. При этом неизбежны потери водяных паров из влажного воздуха межклетников. Как правило, у растений больше устьиц находится с нижней стороны листа, которая меньше нагревается.
Задача
На фото перед вами отпечаток кожицы (эпидермы) листа омелы белой (Viscum album), рис. 1.
Для сравнения — эпидерма листа тюльпана (рис. 2).

Рис. 2. Устьица на поверхности листа тюльпана (срез кожицы подкрашен фиолетовым красителем). Длина масштабного отрезка 100 мкм. Фото с сайта commons.wikimedia.org
На листьях омелы необычайно много устьиц, причем с обеих сторон листа. Почему? У каких еще растений можно ожидать повышения числа устьиц по тем же причинам, что и у омелы?
Подсказка 1
Вспомните, какой образ жизни ведет омела белая.
Подсказка 2
Подумайте, для чего при образе жизни омелы может быть нужно ускоренное испарение воды листьями.
Решение
Испарение воды через устьица — важный механизм усиления тока воды по проводящей системе растения. Главный механизм создания тока воды по ксилеме (древесине) растения — это высокое осмотическое давление в корне. Движение ксилемного сока возможно даже тогда, когда испарения воды через листья нет совсем (см. картинку дня «Любители кленового сока»), но с листьями оно все-таки происходит гораздо быстрее.
Как правило, для растений характерно неравномерное распределение устьиц по сторонам листа: их больше на нижней стороне листа (представьте себе, например, хвоинку пихты: все или почти все ее устьица собраны в две светлые полосочки с нижней стороны). Верхняя сторона листа нагревается солнцем и могла бы усиленно испарять воду, но тогда растение тратило слишком много воды, поэтому бы на верхней стороне листа устьиц обычно нет или очень мало. У омелы же обе стороны листа несут частые устьица. Такое строение не располагает к экономии воды, но дополнительно усиливает испарение в солнечные дни.
Омела белая — паразит деревьев. У нее есть фотосинтезирующие листья, но нет настоящих корней, растущих в почве: омела формирует специальный корень — гаусторию, которая внедряется в проводящую систему растения-хозяина и берет оттуда воду и питательные вещества. Такие растения называются полупаразитами. Точно не известно, в какой мере омела зависит от органических веществ из проводящей системы хозяина.
У паразитических растений работают все те же механизмы водного обмена, что и у обычных растений. Но, чтобы получить воду, они должно «перебороть» растение-хозяина: испарять сильнее и/или создать осмотическое давление в гаустории выше, чем осмотическое давление в проводящей системе растения-хозяина. Омела белая продолжает усиленно испарять воду даже в засуху — вплоть до увядания хозяина. Усиленным испарением обладают и другие паразитические растения, сохранившие листья, — например, марьянник (Melampyrum); поэтому он так быстро увядает, если его сорвать.
Послесловие
Все паразитические растения можно разделить на две большие группы: растения, паразитирующие на грибах, или микогетеротрофные растения, и растения, паразитирующие на других растениях.
Микогетеротрофные растения — это «растения-обманщики». Большинство высших растений вступает во взаимовыгодный симбиоз (см. Мутуализм) с грибами: грибы получают от растений органические вещества, растения получают от грибов воду и минеральное питание. Микогетеротрофные растения берут у грибов всё. Чаще всего они «подключаются» к чужой мутуалистической микоризной сети, но есть и несколько видов, паразитирующих на грибах-сапротрофах. Про пару недавно открытых микогетеротрофов — сциафилу (Sciaphila) и тисмию (Thismia) — см. картинки дня Сциафила без хлорофилла и Бесхлорофилльная тисмия.
Микогетеротрофность возникла в ходе эволюции более 40 раз в разных группах, от печёночников до цветковых растений. Чаще всего она возникала у растений, живущих в глубокой тени под пологом леса, и, конечно, больше всего таких микогетерофов в вечнозеленых тропических лесах. Схожий тип питания приводит к сходным изменениям во внешнем виде: у полных микогетеротрофов не синтезируется хлорофилл, листья редуцируются до бесцветных чешуй, растение переходит к преимущественно подземному образу жизни — его можно обнаружить только по периодически появляющимся надземным цветоносам с цветками. На молекулярно-генетическом уровне обнаружено, что гены пластид, связанные с фотосинтезом, теряют функциональность: обратной дороги у микогетеротрофов нет.
Известно более 400 видов полностью микогетеротрофных растений — казалось бы, это капля в море на фоне общего разнообразия растительного мира. Однако сам по себе редкий феномен микогетеротрофии, привлекая внимание исследователей, подтолкнул их к открытию разных вариантов частичной микогетеротрофии. Оказывается, что около 10% всех растений являются микогетеротрофами на некоторых стадиях своей жизни — чаще всего на стадии прорастания споры (некоторые папоротникообразные) или семени без запаса питательных веществ (большинство орхидей). Правда, семена орхидей научились проращивать и без грибов, на стерильной питательной среде с сахарами. Кроме того, обнаружено), что часть зеленых орхидей и во взрослом состоянии не полностью обеспечивает свои потребности в органических веществах фотосинтезом, а частично получает их от грибов (см. G. Gebauer et al., 2016. Partial mycoheterotrophy is more widespread among orchids than previously assumed).
Растения-паразиты других растений прикрепляются специфическими корнями — гаусториями — к корням или побегам растений-хозяев. Выделяют полупаразитов и полных паразитов. Полные паразиты, как и микогетеротрофы, — бесхлорофилльные растения с редуцированными, чешуевидными листьями. Полные паразиты нуждаются не только в воде и минеральных веществах из ксилемы, но и в органических веществах из флоэмного сока. Полупаразиты — это растения, не утратившие способность к самостоятельному фотосинтезу. Впрочем, они могут в той или иной степени зависеть от органических веществ, украденных у растений-хозяев — без специального исследования, как с марьянником, этого не определить (см. L. E. Nave et al., 2018. Radiocarbon Suggests the Hemiparasitic Annual Melampyrum Lineare Desr. May Acquire Carbon from Stressed Hosts).
Гаустории будут отличаться по деталям строения у разных растений-паразитов, в зависимости от того, внедряется ли паразит во флоэму хозяина или нет (рис. 3 и 4).

Рис. 3. Гаустории омелы белой. Из книги Юлиуса фон Сакса (Julius von Sachs) «Лекции по физиологии растений», 2-е издание, Лейпциг, 1887. Изображение с сайта ru.wikipedia.org

Рис. 4. Гаустория повилики (она еще не достигла проводящего пучка стебля). Фото © Moran Farhi с сайта www-plb.ucdavis.edu
Мы обсудили, как омела усиливает ток воды в своей проводящей системе за счет усиления испарения. Дополнительно усилить испарение можно, если устьица не будут реагировать на изменение влажности воздуха и будут открыты всегда — и действительно, у некоторых полупаразитов устьица могут просто не закрываться ни днем, ни ночью. Растения-хозяева таких полупаразитов при нехватке воды в почве или при сильном заражении паразитом будут увядать. Омела белая относится к таким растениям. Устьица этого вида малочувствительны к абсцизовой кислоте — гормону водного стресса, к тому же исследования показывают, что омела каким-то образом уменьшает концентрацию абсцизовой кислоты в ксилемном соке, который она забирает у хозяина.
Кстати, у мхов устьица есть только на коробочках. А у печеночных мхов устьиц нет вообще никогда — устьицами иногда называют поры на их слоевище, имеющие совсем другое строение. Спорофит мха (диплоидный организм, образующий споры) вырастает из оплодотворенной яйцеклетки прямо на гаметофите (гаплоидный организм, образующий гаметы). Существовали предположения, что устьица спорофиту нужны для скорейшего высыхания коробочки, но сейчас ученые склоняются к тому, что испарение воды усиливает ток растворенных веществ от гаметофита к спорофиту. Можно условно сказать, что спорофит мхов является полупаразитом на своем же собственном родителе!
У полных паразитов листья редуцированы, и испарение через устьица в любом случае будет неэффективным из-за очень небольшой площади листа. Для них оказывается возможен сброс воды в жидком виде через специальные структуры — гидатоды. Гидатоды могут быть устроены по-разному: это либо поры в эпидерме листа (похожие на устьица, но без механизма открывания-закрывания), к которым подходит сосудистый пучок, или волоски эпидермы. Такой механизм реализован как у некоторых полупаразитических растений, так и у полных паразитов — заразихи, Петрова креста. У последнего капельки воды выделяются через чешуевидные листья на подземном побеге — корневище.
Получается, что полные паразиты с редуцированными листьями и низким уровнем испарения не приводят к таким заметным потерям воды. Основной ущерб они наносят, забирая у хозяина органические вещества, аминокислоты и сахара из флоэмного сока. Если сравнить биомассу системы «хозяин + паразит» с биомассой здорового растения-хозяина, может оказаться, что биомасса системы «хозяин + паразит» значительно меньше. Предполагают, что в этом случае рост растения-хозяина сильно ограничен из-за того, что паразит забирает те или иные лимитирующие вещества.
Потери органических веществ опосредованно влияют на способность корня поглощать воду. Корень — гетеротрофный орган, который полностью зависит от поставок органических веществ надземной частью растения. При этом корень тратит большие количества энергии, активно «закачивая» ионы солей в клетки. Таким способом в клетках корня повышается осмотическое давление и создается ток воды из почвы в корень. Если паразитическое растение забирает из проводящей системы хозяина органические вещества, предназначенные для корня, то корень хозяина может испытывать нехватку энергии для поглощения воды и в конечном счете погибнуть от истощения.
Интенсивность паразитизма связана с продолжительностью жизненного цикла паразита. Многолетние паразиты, такие, как Петров крест, или полупаразиты, такие, как омела, отличаются медленным ростом и редко доводят хозяина до гибели. Однолетние паразиты и полупаразиты (многие заразихи, стриги (Striga), повилики) в прямом смысле высасывают жизнь из своих хозяев.
И у полупаразитов, и у полных паразитов поглощение раствора из проводящей системы хозяина связано с повышением осмотического давления в гаустории. Поток воды из проводящей системы хозяина перенаправляется в проводящую систему паразита благодаря тому, что в его теле, прежде всего в корнях-гаусториях, содержатся вещества, повышающие осмотическое давление. Осмотическое давление повышают любые растворенные вещества. Это могут быть те же самые сахара, аминокислоты, минеральные вещества, которые паразит забирает из проводящей системы растения-хозяина.
Литература:
1. P. Escher, A. D. Peuke, P. Bannister, S. Fink, W. Hartung, F. Jiang, H. Rennenberg. Transpiration, CO2 assimilation, WUE, and stomatal aperture in leaves of Viscum album (L.): Effect of abscisic acid (ABA) in the xylem sap of its host (Populus x euamericana) // Plant Physiology and Biochemistry. 2008. V. 46. Issue 1. P. 64–70.
2. D. Haig. Filial mistletoes: the functional morphology of moss sporophytes // Annals of Botany. 2013. V. 111. Issue 3. P. 337–345.
3. L. Irving & D. Cameron. You are What You Eat: Interactions Between Root Parasitic Plants and Their Hosts // Advances in Botanical Research. 2009. V. 50. P. 87–138. DOI: 10.1016/S0065-2296(08)00803-3.
4. Mycoheterotrophy: The Biology of Plants Living on Fungi. Ed.: Vincent Merckx. New York: Springer, 297–342.
5. M. C. Press, J. D. Graves, G. R. Stewart. Physiology of the interaction of angiosperm parasites and their higher plant hosts // Plant, Cell & Environment. 1990. V. 13. Issue 2. P. 91–104.
-
Внимательно посчитал устьица на фото тюльпана и омелы.
На кждый квадрат 100х100 мкм приходится чуть меньше одного устьица и там и там.
(А именно, отношение устьиц к площади у омелы 11:16 (нижний правый квадрат 4х4), у тюльпана 10:12(вся фотка, 4х3). За единицу площади взял квадрат 100х100мкм. Пусть даже 12:16 - это только 3/4. У тюльпана 5/6. Привожу к общему знаменателю: 9/12 и 10/12. Этот элементарный подсчет показывает, что у тюльпана устьица даже чаще.
Где необычайное их скопление ? Где основание , откуда взято условие задачи??
Что я неправильно вижу??-
Добрый день. Возможно, вы не учли устьица омелы, которые находятся не в фокусе и у них не видна устьичная щель -- это фото реплики эпидермиса, она неровная. Попробуйте взять небольшой квадрат, в котором все устьица в фокусе, получите разницу с тюльпаном в несколько раз.
-
В том участке, где я считал, все сомнительные, нечеткие места я защитывал в устьица.
Вся приведенная фотография выглядит равномерной в смысле плотности частоты их (устьиц) расположения. Мало того, вверху слева есть увеличенный участок, где еще увеличено устьице, и этот поясняющий кучочек облегчает трактовку мутностей и нечеткостей фото.
Только допуская, что приведенная фотография омелы в принципе не подходит в качестве иллюстрации высокой частоты расположения устьиц (например, (а) это фото не той части поверхности, где много устьиц, или (б) отрезок линейки на самом деле не 400мкм) можно согласиться с мыслью, что у омелы устьиц больше, чем у тюльпана.
Иначе- посчитав фото верным- имеем доказательство несоответствие условия задачи реальности.
Учитывая, что к-во устьиц у омелы и тюльпана в реальности сильно отличаются
Делаю вывод, что эта фотография -ошибочная. Тут опечатка (либо фото нужно другое, либо подпись, трактующая размеры масштабного отрезка)-
Я тоже считала устьица на этой фотографии и сравнивала количество устьиц на единицу площади. У меня на квадрате 400 на 400 в нижнем правом углу выходит совсем не 11 устьиц, как у вас, а гораздо больше.
-
Фантастика:
Два человека смотрят на одну и ту ж фотку и считают на ней предметы. И разница подсчетов выходит большая!!
Вы насчитываете там (в квадрате со стороной в этот отрезочек белого цвета) устьиц в несколько раз больше дюжины??
Тогда простите, я что-то не то делаю.
Возможно, просто арифметика - это не мое.
Беру все свои слова обратно. И больше считать тут ничего не буду.
А послесловие с моей точки зрения снова интереснее задачи, это не в первый раз уже в этой рубрике такое случается)-
Я сейчас взяла прямоугольник такого же размера (примерно 500 на 300 мкм), как вся фото эпидермиса тюльпана (там все же ближе к 500 мкм по ширине, нежели к 400), и все посчитала еще раз :) У меня в нем получается 20 несомненных устьиц (на фото эпидермиса тюльпана их 10). Так что да, примерно в 2 раза больше. К сожалению, не могу сюда прикрепить размеченную фотографию с подсчетами, но если Вы укажете свой e-mail, могу отправить ее туда.
Да, жанр как раз подразумевает интересное послесловие :)
-
-
-
-
-








Рис. 1. Устьица на поверхности листа омелы. Длина масштабного отрезка 400 мкм