Бактерия из пустынного озера украла фотосинтетический аппарат у другой бактерии и усовершенствовала его

В озере на западе пустыни Гоби обнаружен новый вид бактерий, способный к фотосинтезу, — Gemmatimonas phototrophica. Этот вид относится к группе Gemmatimonadetes, представителям которого фотосинтез, вообще говоря, не свойственен. Оказалось, что G. phototrophica получила свою фотосистему — комплекс белков, осуществляющий фотохимическую реакцию, — от какой-то пурпурной бактерии в результате горизонтального переноса генов. Что более удивительно, в процессе эволюции эта фотосистема приобрела другую структуру и стала работать гораздо эффективнее своего «прототипа».
При слове «фотосинтез» мы прежде всего представляем себе растения. В крайнем случае — водоросли. Но самые значимые фотосинтетики на Земле — это бактерии. Даже пластиды растений и водорослей, с помощью которых те осуществляют фотосинтез, — потомки древних цианобактерий, первыми освоивших фотосинтез с выделением кислорода. Этот процесс помог им кардинально преобразить облик земного шара.
Идея, лежащая в основе этой биологической инновации, гениально проста: объединить два типа фотосистем — молекулярных «машинок» для передачи электронам энергии квантов света — таким образом, чтобы они работали вместе, развивая большую мощность (см. врезку). Это открыло цианобактериям (а вслед за ними — водорослям и растениям) путь к получению большого количества энергии из солнечного света и простой воды (которой вокруг было в буквальном смысле залейся). Эту фотосинтетическую инновацию так до сих пор никто и не повторил.
Помимо цианобактерий, многие другие бактерии тоже «умеют» фотосинтезировать. У них фотосистемы какого-нибудь одного типа — либо феофитин-хинонового, как у пурпурных бактерий, либо Fe-S-типа, как у зеленых серных бактерий. Мощность таких биологических «солнечных батарей» существенно ниже.
Тем не менее находятся экзотические бактерии, которые в процессе эволюции усовершенствовали свои фотосистемы без таких радикальных преобразований, как у цианобактерий, и «выжимают» из них рекордные показатели.
Принципы работы фотосистем у разных групп организмов
У любого фотосинтезирующего организма свет Солнца собирается антенными (или светособирающими — англ. light-harvesting) комплексами. Они имеют различное строение и не показаны на рисунках. От светособирающих комплексов энергия возбуждения электрона (ее квант называется экситоном) передается в реакционный центр — комплекс белков. Электрон в одном из его активных центров получает квант энергии и переходит на высокий энергетический уровень, с которого постепенно «спускается», проходя по белкам электрон-транспортной цепи, как по ступенькам. При этом один из белковых комплексов — цитохромный комплекс, который работает как молекулярный насос. Он использует энергию проходящих через него электронов для перекачки протонов через мембрану: с ее внутренней стороны на наружную (у бактерий) или из стромы хлоропласта в люмен (просвет, см. Lumen) тилакоида (у цианобактерий, водорослей и растений). Этот градиент используется для синтеза АТФ.
Самая простая фотосистема — феофитин-хиноновая. В ней электроны двигаются по кругу и сами по себе никуда не расходуются — только их энергия тратится на поддержание градиента протонов на мембране. Поэтому в таком варианте фотосинтеза никакой восстановитель (источник электронов) не нужен.
Феофитин-хиноновая фотосистема. Рисунок © Георгий Куракин
Но замкнутый цикл не очень удобен — он позволяет только синтезировать АТФ, но не дает запасти восстановительных эквивалентов в виде НАД/НАДФ. Эта проблема решена у зеленых серных бактерий — в их Fe-S-фотосистеме цикл частично разомкнут, и часть электронов уходит на восстановление НАД. При этом возникает вопрос, откуда брать новые. Для этого бактерии используют сероводород. Правда, он есть не везде, да и в качестве «отходов» образуется элементарная сера — желтое твердое вещество, с которым не очень-то удобно иметь дело.
Fe-S-фотосистема. Рисунок © Георгий Куракин
Более 2,5 млрд лет назад цианобактериям удалось провернуть биохимический трюк: они смогли объединить феофитин-хиноновую и Fe-S-фотосистему в один большой комплекс, чтобы они работали вместе. Вместе с хлоропластами — «потомками» цианобактерий — этот тип фотосистем попал в клетки многих простейших, а также всех водорослей и растений. Предположительно, предки цианобактерий получили обе фотосистемы в результате горизонтального переноса генов — как и бактерия, про которую мы расскажем далее. При этом цианобактерии всё равно остаются непревзойденными мастерами эволюции фотосинтеза, так как «позаимствовали» гены фотосинтеза от двух разных фотосинтетиков, каждый из которых обладал только одной фотосистемой. Феофитин-хиноновая фотосистема дала начало фотосистеме II, а Fe-S-фотосистема — фотосистеме I. Два реакционных центра, работая вместе, «поднимают» электрон выше, чем поодиночке. За счет этого комбинированная фотосистема может использовать воду вместо сероводорода для питания своего разомкнутого цикла. При этом выделяется не сера, а газообразный кислород. Такой тип фотосинтеза называется оксигенным и сыграл огромную роль в формировании современного облика нашей планеты.


Комбинированная фотосистема цианобактерий. Рисунок © Георгий Куракин
Подробнее об «изобретении» кислородного фотосинтеза цианобактериями можно прочитать в новостях: Новый вид цианобактерий проливает свет на эволюцию кислородного фотосинтеза («Элементы», 17.01.2022); Перенастроенные «молекулярные часы» показали более точное время появления кислородного фотосинтеза («Элементы», 12.10.2021); Геномы новооткрытых цианобактерий свидетельствуют о позднем появлении кислородного фотосинтеза («Элементы», 03.04.2017).



Ученые делят бактерии на филумы — большие таксономические единицы, соответствующие типам животных. Ранее считалось, что фотосинтез — это «прерогатива» только семи филумов бактерий: Cyanobacteria (уже обсуждавшиеся цианобактерии — лидеры инноваций в мире фотосинтеза), Proteobacteria (к ним относятся пурпурные бактерии с их феофитин-хиноновой фотосистемой), Chlorobi (зеленые серные бактерии с их Fe-S-фотосистемой), Chloroflexi (зеленые несерные бактерии — несерные потому, что обладают феофитин-хиноновой фотосистемой, не требующей ни сероводорода, ни другого восстановителя), Firmicutes (точнее — входящие в этот филум гелиобактерии, фотосинтетические азотфиксирующие анаэробы, которых можно найти на рисовых полях), Acidobacteria — группа бактерий, обитающих в почве.
Но в 2014 году список этих филумов пополнился. Группа ученых из Чехии и Китая провела скрининг более чем 5000 изолятов из различных водоемов на присутствие бактериохлорофилла (Y. Zeng et al., 2014. Functional type 2 photosynthetic reaction centers found in the rare bacterial phylum Gemmatimonadetes). В образце из Лебединого озера (Tian E Hu) в пустыне Гоби обнаружился ранее неизвестный вид бактерий, обладающий бактериохлорофиллом (рис. 2). Этот вид относится к филуму Gemmatimonadetes, который сам по себе мало изучен.

Рис. 2. Электронная микрофотография представителя нового вида — Gemmatimonas phototrophica. Фото с сайта diamond.ac.uk
Наличие бактериохлорофилла ясно указывало на способность этого вида к фотосинтезу. Анализ генома выявил присутствие генов, кодирующих компоненты феофитин-хиноновой фотосистемы — как у пурпурных бактерий. Более того, новооткрытая бактерия именно у них эту фотосистему и «позаимствовала» — путем горизонтального переноса генов.
Ученых удивил сам факт такого переноса целого фотосинтетического кластера, превратившего ранее не фотосинтезирующую бактерию в фотосинтетика. Этот факт в очередной раз ставит вопрос о том, насколько сложно придать бактерии способность к фотосинтезу, просто вставив в ее ДНК нужные гены. Такой трюк может иметь большое прикладное значение — например, для получения экологически чистой энергии.
Однако новый вид (ему присвоили говорящее название Gemmatimonas phototrophica) в процессе эволюции не только «стащил» эту фотосистему, но и сделал с ней еще кое-что интересное.
У пурпурных бактерий белки реакционного центра (reaction centre, RC) образуют большой надмолекулярный комплекс, который окружен цилиндром из других белков, формирующих светособирающий комплекс 1 (light-harvesting complex 1, или LH1). Получается комплекс RC-LH1, имеющий форму колеса. Но LH1 — это лишь промежуточное звено передачи энергии. Непосредственно свет захватывается светособирающим комплексом 2 (LH2), который также является «колесиком» из белков, но меньшего диаметра. Цилиндры LH2 не соединены жестко с комплексом RC-LH1 и просто находятся рядом в мембране бактериальной клетки. При этом в реальных условиях они все равно находятся очень близко к LH1, что обеспечивает передачу энергии возбуждения электрона от LH2 к LH1 и далее к реакционному центру (рис. 3).
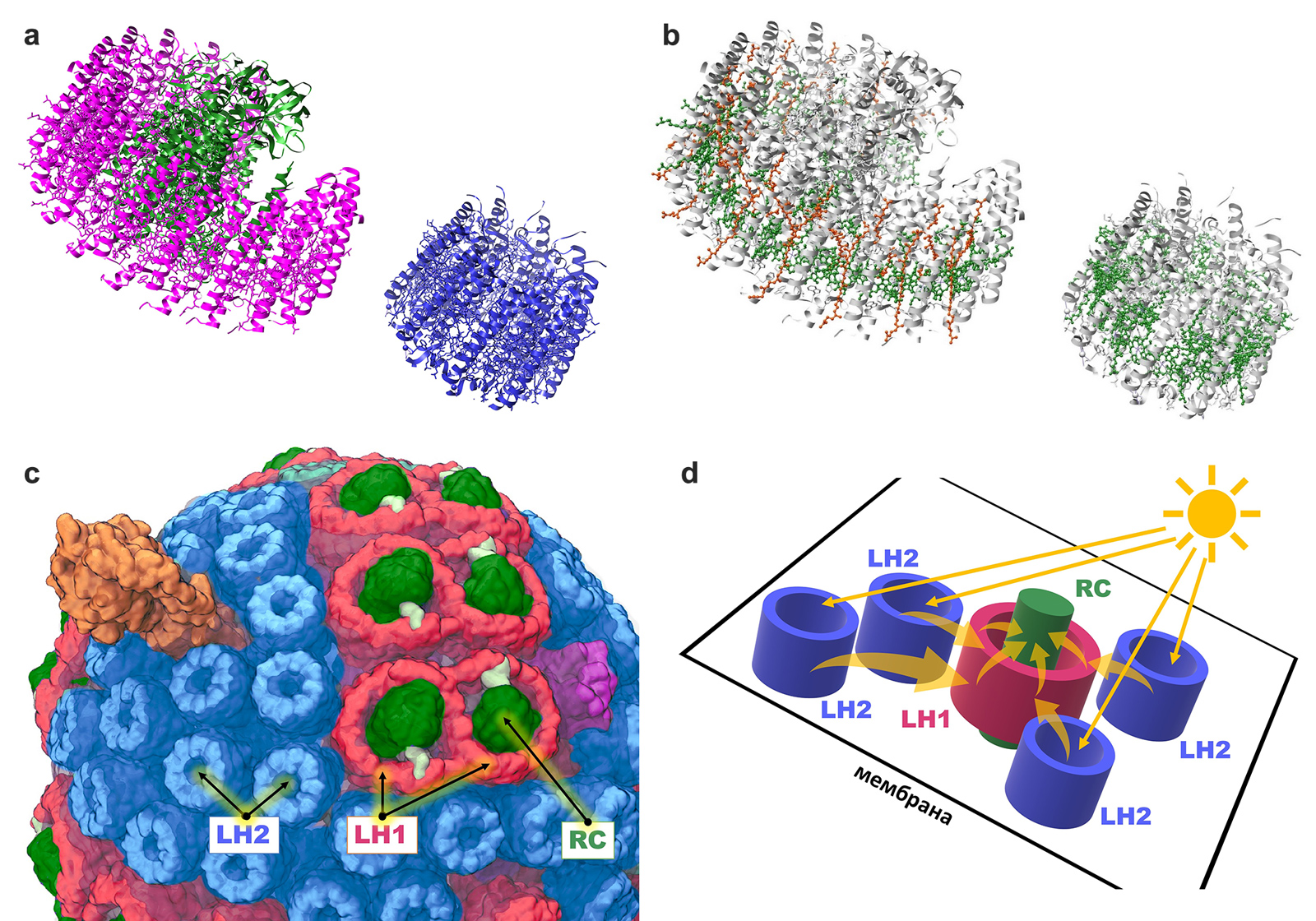
Рис. 3. a — структуры комплексов RC-LH1 и LH2 феофитин-хиноновой фотосистемы пурпурной бактерии Rhodobacter sphaeroides. Так как они не образуют прочного комплекса, их нельзя выделить вместе при кристаллографии. Чтобы показать их рядом, автору пришлось искусственно переместить один из них в программе UCSF Chimera. Реакционный центр, напоминающий толстую ось колеса, обозначен зеленым цветом. Он окружен пурпурным цилиндром — комплексом RC-LH1. Рядом расположен синий цилиндрический белковый комплекс меньшего диаметра — LH2. «Сверху» — внутренняя сторона мембраны, «снизу» — внешняя. b — Внутри каждого белкового цилиндра проходит своеобразное кольцо, образованное ансамблем выстроившихся «в круг» пигментов, непосредственно улавливающих свет — бактериохлорофилла (зеленого) и каротиноидов (оранжевых). Именно они улавливают свет и передают его дальше, являясь «рабочей частью» антенного комплекса. «Сверху» — внутренняя сторона мембраны, «снизу» — внешняя. Рисунки © Георгий Куракин, изображения созданы на основе структур PDB 7PIL и 7PBW в программе UCSF Chimera. c — несмотря на отсутствие тесной межмолекулярной связи, в мембране клеток пурпурных бактерий RC-LH1 и LH2 расположены рядом, что позволяет экситонам «перескакивать» с LH2 на RC-LH1. Рисунок из статьи M. Sener et al., 2014. Overall energy conversion efficiency of a photosynthetic vesicle. d — схематичное изображение взаиморасположения RC-LH1 и LH2 в мембране клетки пурпурных бактерий и переноса энергии между ними. Прямые стрелки обозначают свет (фотоны), изогнутые стрелки — перенос энергии (экситоны). Рисунок © Георгий Куракин
Структура светособирающего компекса бактерии Gemmatimonas phototrophica была расшифрована только недавно — в феврале 2022 года. Она оказалась не менее удивительной, чем сам факт горизонтального переноса необходимых для фотосинтеза генов. Ее структура претерпела значительные «усовершенствования» по сравнению с предковым состоянием.
Реакционный центр этой фотосистемы устроен почти так же, как его «прототип» у пурпурных бактерий. И он так же окружен комплексом LH1, как ось колеса — ободом. Но «маленького» комплекса LH2 у Gemmatimonas phototrophica нет — его функцию выполняет больший по размеру комплекс LHh, который обхватывает комплекс LH1 еще одним внешним кольцом (рис. 4).
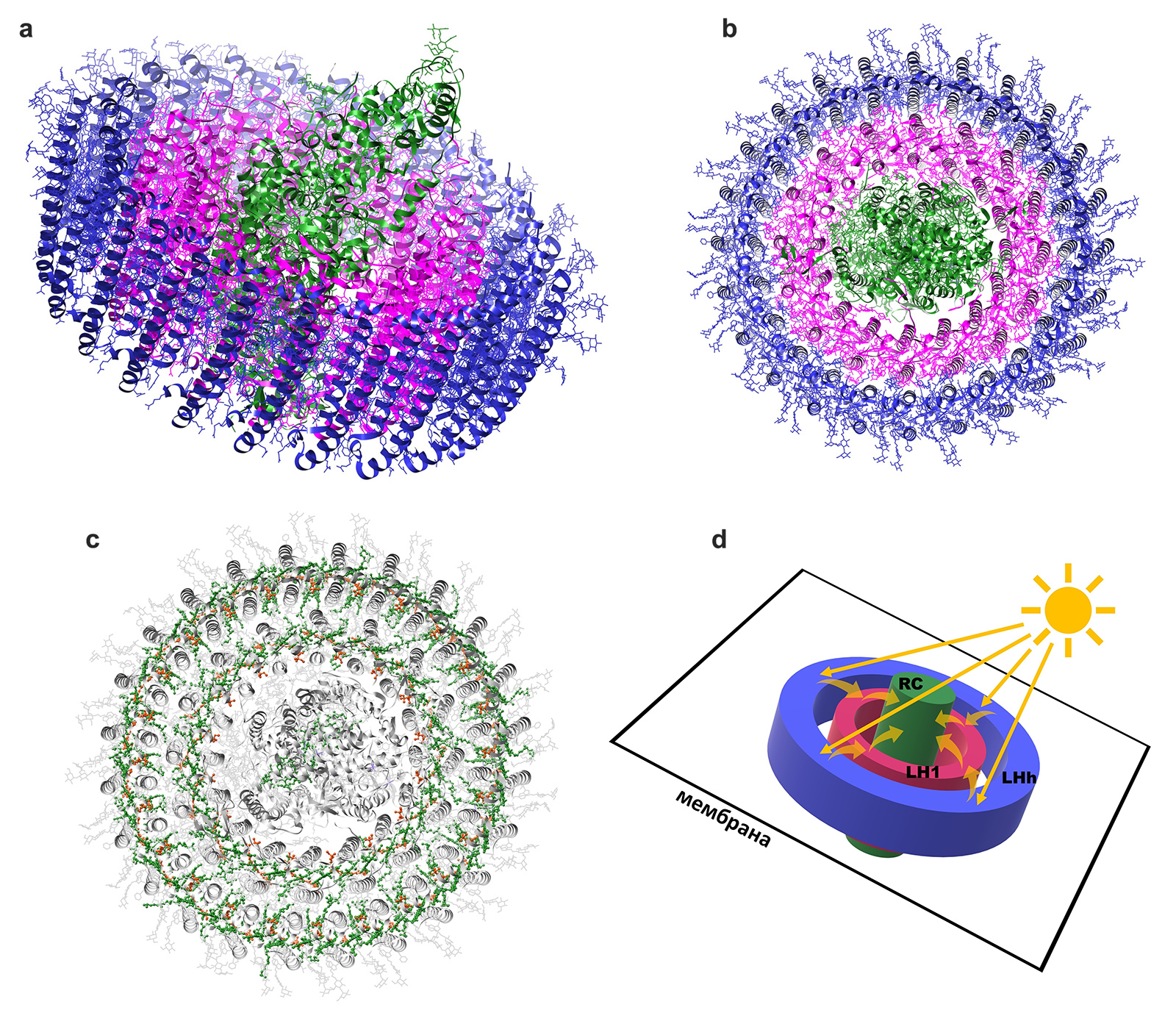
Рис. 4. a — антенные комплексы фотосистемы бактерии Gemmatimonas phototrophica образуют единый белковый агрегат вместе с реакционным центром, который также имеет форму колеса, лежащего в плоскости мембраны клетки. Цвета соответствуют цветам на рис. 3. Синим цветом закрашен комплекс LHh, функционально заменяющий LH2 и образующий дополнительное внешнее кольцо. b — то же самое, вид сверху. Изображения созданы в программе UCSF Chimera на основе структуры PDB 7O0V. c — как и в антенных комплексах пурпурных бактерий, каждый из светопоглощающих комплексов заключает в себе ансамбль пигментов — бактериохлорофилла (зеленого) и каротиноидов (оранжевых), которые служат ему рабочей частью. d — схематичное изображение строения фотосистемы бактерии Gemmatimonas phototrophica и передвижения электронов внутри нее. Рисунки © Георгий Куракин
Такая структура делает фотосистему бактерии Gemmatimonas phototrophica рекордно большой (в молекулярных масштабах) — но вместе с тем потрясающе эффективной. Благодаря тесной связи обоих антенных комплексов энергия передается между ними и на реакционный центр гораздо быстрее: постоянная времени передачи экситона между «внешним» и «внутренним» светособирающими комплексами у нового комплекса составляет всего 2 пикосекунды против 3–5 пикосекунд у пурпурных бактерий. Проще говоря, более компактная структура фотосистемы позволяет новому виду лучше улавливать энергию солнечного света. Так что он сумел значительно улучшить ту «модель» фотосистемы, которую получил путем горизонтального переноса генов. Самое интересное, что для этого потребовалось приобретение только одного дополнительного гена — вот где потрясающая экономия и эффективность!
СМИ уже успели многократно окрестить фотосистему бактерии Gemmatimonas phototrophica «уникальной». Это верно лишь отчасти: по механизму действия это в целом такая же феофитин-хиноновая фотосистема, как и у пурпурных бактерий. Светособирающие комплексы менялись в процессе эволюции многократно в разных эволюционных линиях. Однако, коллеги-журналисты правы в том, что такая структура антенных комплексов пока больше не описана ни у одного вида — и она крайне интересна своей экономичностью и эффективностью.
Источники:
1) Pu Qian, Alastair T. Gardiner, Ivana Šímová, Katerina Naydenova, Tristan I. Croll, Philip J. Jackson, Nupur, Miroslav Kloz, Petra Čubáková, Marek Kuzma, Yonghui Zeng, Pablo Castro-Hartmann, Bart van Knippenberg, Kenneth N. Goldie, David Kaftan, Pavel Hrouzek, Jan Hájek, Jon Agirre, C. Alistair Siebert, David Bína, Kasim Sader, Henning Stahlberg, Roman Sobotka, Christopher J. Russo, Tomáš Polívka, C. Neil Hunter, Michal Koblížek. 2.4-Å structure of the double-ring Gemmatimonas phototrophica photosystem // Science advances. 2022. DOI:10.1126/sciadv.abk3139.
2) Y. Zeng, F. Feng, H. Medová, J. Dean, & M. Koblížek. Functional type 2 photosynthetic reaction centers found in the rare bacterial phylum Gemmatimonadetes // Proceedings of the National Academy of Sciences. 2014. DOI: 10.1073/pnas.1400295111.
Георгий Куракин
-
Такие эпизоды (если эти бактерии действительно конкурировали за качество этого фотосинтезирующего аппарата на общем субстрате, и «похититель» до этого не обладал похожим аппаратом) косвенно могут указывать на возможность существования «абстрактных регуляторных генетических сетей» - неких групп генов, взаимодействие между которыми выражают некие математические зависимости (например, своими активностями могут выражать арифметическую прогрессию, или соответствовать геометрическим задачам - активность одного гена будет зависеть от того, насколько точно пропорции активностей трёх других генов выражают стороны треугольника, вписанного в окружность радиусом, соответствующим активности четвёртого гена - пример кривой, с потолка). Эволюция точности таких сетей проходила бы под давлением отбора одного фенотипа, а потом могла бы сцепиться с «похищенными» генами нового фенотипа, заменив его «оригинальную» регуляторную сеть. Т.е., решив одну задачу, генофонд обретает способность решать сразу класс аналогичных задач (возможно лучше, чем аналогичная сеть, отбираемая под менее жёстким давлением отбора на другом фенотипе).
Не уверен, существует ли в одноклеточной природе такое проявление эволюционной динамики, а если существует, то насколько широко, и насколько примитивной может быть генетический аппарат клетки, чтобы уже демонстрировать подобные механизмы абстракции, но в многоклеточных конструкциях (например, в мозгу, или даже в бактериальном мате) путём опосредования взаимодействий генов через механику межклеточных сигнальных молекул такая «эволюция абстракций» (накопление генофондом «универсальных решателей») кажется неизбежной. -
Георгий, спасибо за статью. В общем-то, становится всё более очевидным, что, по-видимому, почти все современные филы бактерий (кроме, разве что, цианобактерий), обладающие аппаратом хлорофильного фотосинтеза, получили его в результате горизонтального переноса.
-
Автор рассказывает, что и цианобактерии получили радость через ГПГ.
Предположительно, предки цианобактерий получили обе фотосистемы в результате горизонтального переноса генов
-
Автор рассказывает, что и цианобактерии получили радость через ГПГ.
Да, я знаком с мнением автора, но в последние пару десятков лет появилось много работ, приводящих к обратному выводу, см., например: https://www.pnas.org/doi/full/10.1073/pnas.0605709103 Так что, в этом вопросе мы с автором расходимся.-
Ссылка познавательная, спасибо!
Хорошо, примем, что предки цианобактерий являются родоначальником всех видов хлорофильного фотосинтеза для всей биоты. Однако сложно представить возникновение двух фотосистем без участия горизонтального переноса.
Конечно, оба реакционных центра произошли от общего предка. Но вряд ли это произошло внутри одной линии клеток. Отбор препятствует расточительному наличию двух одинаковых групп генов в одной бактерии. (Хотя у многоклеточных такое в порядке вещей). Более вероятно, что две фотосистемы некоторое время эволюционировали по отдельности, затем фотосистема II попала в цианобактерию посредством горизонтального переноса.
Это не противоречит выводам статьи по вашей ссылке. На Fig. 1 всё описанное происходило внутри овала Procyanobacteria. А линия, в которой возникла фотосистема II, до нас не дожила или же науке неизвестна.-
По-видимому, реакционный центр гелиобактерий наиболее похож на первый архаичный реакционный центр общего предка всех цианобактерий. А в нём переносчиками электронов могут выступать как хиноны, так и ферредоксины. В зависимости от внешних условий, могло оказаться выгодным использовать то один, то другой носитель (см. например, пояснения в обсуждаемой статье). Да и управлять процессом проще, если они разделены. А дублирование генов, после которого они могут независимо эволюционировать по разным траекториям под давлением разных факторов отбора, в биоте событие весьма рутинное. Далее быстро выяснилось, что при работе в тандеме две фотосистемы могут использовать в качестве источника электронов практически неисчерпаемый ресурс - воду, и это покрывает все издержки, включая необходимость выстранивания защиты от кислорода, особенно, его активных форм. См. так же рисунок 1.b и пояснительный текст к нему вот в этой статье: https://onlinelibrary.wiley.com/doi/10.1111/gbi.12322
-
Согласен с этими аргументами, кроме:
дублирование генов, после которого они могут независимо эволюционировать по разным траекториям под давлением разных факторов отбора, в биоте событие весьма рутинное
Да, рутинное, но у многоклеточных эукариот. А для бактерий можете ли привести какой‑нибудь другой пример дублирования целого комплекса генов? Полагаю, для бактерий это существенно менее вероятно. А более вероятно, что два комплекса генов эволюционируют по отдельности в разных линиях клеток, а затем посредством ГПГ встречаются в одной клетке.
Такое участие ГПГ никак не противоречит ни остальным вашим аргументам, ни ссылаемым статьям. И напротив, ни из чего не следует, будто реакционные центры обоих типов I и II всегда существовали в линии предков цианобактерий и никогда не разделялись по двум разным линиям (линии прямых предков и линии сестринской к предковой).-
У гелиобактерий весь реакционный цетр представлен одним геном, так что никакого комплекса генов дублировать не нужно. А примеров дублированиея одного конкретного гена у прокарит более, чем достаточно. Далее обе копии уже могли эволюционировать и обрастать дополнительными субъединицами, подстраиваясь под совместную работу.
Реакционный центр у предков цианобактерий вначале был один, потом от него произошли предки РЦ первого и второго типов. Вариант независимой эволюции в двух линиях с их последующей встречей не исключён, но, с моей точки зрения, маловероятен. Гораздо проще наладить соаместную работу двух одинаковых РЦ, один из которых постепенно начинает специализироваться на хинонах, а другой на ферредоксинах, чем ту же адаптацию для уже существенно разошедшихся копий, которые к тому же уже успели обрасти своими оригинальными сабъюнитами, светособирающими комплексами и т.д.
-
-
-
-
-
-


Вот ещё одна интересная статья: https://www.quantamagazine.org/dna-jumps-between-animal-spec
Другое дело, что это чрезвычайно редкие события по сравнению с тем, как это происходит у бактерий (для которых это обычное дело). Технически между эукариотами гену "перепрыгнуть" трудно. По всей видимости, эту вероятность может повышать и делать ненулевой ситуация с очень сильным давлением отбора – как в случае токсинов растений и антифризов рыб.
Ну и следует упомянуть, что у эукариот есть друга крутая особенность – половое размножение, которое заменяет им горизонтальный перенос генов. Если бактерии про него знают, то точно не посмеиваются, а просто обалдевают, как у нас тут всё сложно...
Скажите пожалуйста, позволяет ли она создавать такие комплексы белков? (А не только повторять имеющиеся уже в реальности)
Ведь интересно же определить/рассчитать, возможно ли увеличить кол-во этих концентрических колец, , затолкав описанную систему в еще больший бублик, а потом и в ещё - вдруг возможны стабильные конструкции не только из одного и двух бубликов, но из 3х, 4х, ...128ми? - увеличив эффективность системы многажды еще и ещё?
И , "подкидывая" фотонами электрон столько раз, можно же "допрыгать" до черт-те каких возможностей!!
Что там вода - с такими агрегатами бактерии камни смогут грызть! и в кислород обращать!))
А мы - чудовищные солнечные батарейки ляпить сможем как минимум
-
>> возможно ли увеличить кол-во этих концентрических колец, , затолкав описанную систему в еще больший бублик, а потом и в ещё ... И , "подкидывая" фотонами электрон столько раз, можно же "допрыгать" до черт-те каких возможностей!!
концентрическая конструкция не увеличивает потенциал, а ускоряет передачу экситона. То есть, с ними бактерии по-прежнему будут камни не грызть, а лизать, но быстрее.-
Позвольте. уважаемый Новый Участник! Тут же написано!
А, Вы. вероятно. не прочли статью...
Вот же, однозначно констатируется:Два реакционных центра, работая вместе, «поднимают» электрон выше, чем поодиночке. За счет этого комбинированная фотосистема может использовать воду вместо сероводорода для питания своего разомкнутого цикла.
Именно этим - тем, что электрон "поднят выше", автор объясняет нам обретение возможности работать с водой. До тех пор, пока он (электрон) "ниже" - то есть лишь один раз накачан фотоном - "пищей" служит сероводород-
>> А, Вы. вероятно. не прочли статью...
хмм... Вы не первый, кто предполагает, что я пишу комментарии, не читая статьи. Что-то с моим ником, наверное, не так.
Тем не менее, статьи я читаю. Два реакционных центра - у цианобактерии. А концентрические бублики - у Gemmatimonas phototrophica, один единственный реакционный центр во внутреннем бублике и светоулавливающая система (а не реакционный центр №2) во внешнем.-
Что-то с моим ником, наверное, не так.
Встречают‑то по одёжке, что уж тут поделаешь. Взять и сменить псевдоним, не?-
да как-то уже лет пять как нoвый учacтник, а до этого года три был нoвый учаcтник. Уже период встречи должен бы закончиться :)
-
Вот, например, я. Почитываю и комментирую здесь все эти годы. Но совершенно не смог бы сказать, встречал ли я раньше ваш псевдоним. И случись с вами дискутировать, тоже, вероятно, отнёсся бы предвзято.
Как по мне, ваш псевдоним несёт в себе этакий мазохизм. Ваше право, конечно. Но врачи считают мазохизм патологией.-
ну а я стараюсь не дискутировать с конкретным Романом Пеховым или Юрием Фёдоровым, а отвечать на комментарии. Обмениваться идеями без учета личности их носителя.
>>Но совершенно не смог бы сказать, встречал ли я раньше ваш псевдоним.
значит мой ник соответствует моей цели
>>Как по мне, ваш псевдоним несёт в себе этакий мазохизм. Ваше право, конечно. Но врачи считают мазохизм патологией.
надеюсь, врачи не считают ничего плохого насчет ставящих диагноз по нику. Потому что у меня, например, ники, напоминающие настоящие имена и фамилии, вызывают недоверие
-
-
-
-
-
Что бы ни было в Вашем нике - в процитированном автор говорит о том, что пара бубликов подкидывает электрон выше. Вы же утверждаете, что этого не происходит. Вот и сделал вывод, что не читали.
Как иначе оправдать несоответствие?
НаучИте.)-
учу
>> Предположительно, предки цианобактерий получили обе фотосистемы в результате горизонтального переноса генов — как и бактерия, про которую мы расскажем далее. При этом цианобактерии всё равно остаются непревзойденными мастерами эволюции фотосинтеза, так как «позаимствовали» гены фотосинтеза от двух разных фотосинтетиков, каждый из которых обладал только одной фотосистемой. --->> Феофитин-хиноновая фотосистема дала начало фотосистеме II, а Fe-S-фотосистема — фотосистеме I.<<--- Два реакционных центра, работая вместе, «поднимают» электрон выше, чем поодиночке.
электрон подкидывает выше сочетание двух фотосистем.
>> Анализ генома выявил присутствие генов, кодирующих -->> компоненты феофитин-хиноновой фотосистемы <<-— как у пурпурных бактерий.
у Gemmatimonas phototrophica только одна фотосистема.
Внешний концентрический бублик не является новой фотосистемой, он приемная антенна феофитин-хиноновой фотосистемы. Просто у пурпурных эти приемники плавали отдельными кольцами, а у этой бактерии наделись концентрично
-
-
-
А меня эта программа заинтересовала, которую автор так настойчиво упоминает, и в которой можно рисовать такие штучки:Настойчивое упоминание – не моя прихоть и не желание порекламировать программу, а требование лицензионного соглашения на пользование этой программой.
Скажите пожалуйста, позволяет ли она создавать такие комплексы белков? (А не только повторять имеющиеся уже в реальности)
UCSF Chimera – это визуализатор. Она позволяет только графически демонстрировать уже рассчитанные структуры и выполнять их несложное редактирование. Ваша задача – в другом: нужно рассчитать, как белки группировались бы друг с другом. Такая задача называется белок-белковым докингом (protein-protein docking), и она не входит в функционал этой программы. Но для этого есть много других программ, некоторые решения даже доступны онлайн через браузер. Более того, в целом задача белок-белкового докинга менее сложна, чем задача лиганд-белкового докинга: белки можно спокойно сделать гибкими и выполнять полностью гибкий докинг, учитывая все изгибы молекул. С лигандами-малыми молекулами так не очень выйдет – они требуют параметризации, а это отдельная головная боль. В общем, решений много, поищите.
Единственная трудность – речь идёт о мультибелковом комплексе, и вот лично я затрудняюсь вспомнить какое-нибудь удобное решение для белок-белкового докинга, а уж тем более гибкого, которое могло бы моделировать такие комплексы. Но, возможно, можно что-нибудь запрограммировать на базе имеющихся программ, добавив собственные скрипты на том же Питоне.
Теперь про разгрызание камней. Тут разочарую – абсолютно прав пользователь "новый участник":
концентрическая конструкция не увеличивает потенциал, а ускоряет передачу экситона. То есть, с ними бактерии по-прежнему будут камни не грызть, а лизать, но быстрее.Читая статью, не путайте реакционные центры и компоненты антенного комплекса. Концентрические бублики относятся к антенному комплексу, поэтому сами по себе они электрон никуда не поднимают в итоге. Разделение зарядов ("подъём" электрона) происходит в реакционном центре фотосистемы. Именно от его устройства зависит, насколько высоко электрон поднимется и на какой восстановитель этого хватит (в то время как антенный комплекс обеспечивает улавливание света, позволяя реакции протекать быстрее).
в процитированном автор говорит о том, что пара бубликов подкидывает электрон вышеНе приписывайте мне того, чего я не говорил и не писал.
Я говорил, что два реакционных центра подкидывают электрон выше. Но:
1) это вообще не про главного героя этой статьи, а про цианобактерий и растения;
2) у них ДВЕ фотосистемы, каждая из которых имеет свои антенные комплексы и свой реакционный центр. Вот как раз два центра и подкидывают;
3) на рисунках в этой статье реакционные центры – это не бублики, а втулка зелёная посреди бублика. У растений и цианобактерий они устроены и расположены немного иначе.
В общем, Юрий, Вы просто неправильно поняли статью.
Сколько бубликов ни приделывай, камни грызть не получится.
Chloroflexi называются НЕсерными бактериями, потому что в отличие от хлоробий они не запасают серу внутри клеток, а не потому, что вообще ее не используют.
-
Хлорофлексус использует сероводород (правда, плохо и без удовольствия - лучше водород) для автотрофного роста потому, что фиксация СО2 без источника электронов невозможна. Синтез NADPH происходит через обратный перенос электронов от хинонов, хиноны получают электроны от сероводорода. Фотосинтез участвует в этом процессе только опосредованно, создавая протонный градиент для обратного транспорта. Перенос электронов в фотосистеме остается при этом циклическим.
-
Не понял, в чём поинт. У того же хлороби синтез NADPH тоже происходит через обратный перенос электронов от цитохромов, а цитохромы, в свою очередь, получают электроны от сероводорода. В чём, собственно, принципиальная разница? По сути, все системы фотосинтеза, кроме PSII цианобактерий, просто создают протонный градиент, и лишь у неё отбор электронов от воды происходит непосредственно в самом реакционном центре.
-
Хлоробиум NADPH напрямую без обратного переноса элекронов синтетизирует. То есть поднимает электроны от водорода или сульфида до уровня NADPH за счет энергии света. При таком раскладе транспорт электронов не циклический. Аноксигенные фототрофы со второй фотосистемой к этому не способны.
-
Ну при таком подходе любой поток электронов, создаваемый при помощи реакционного центра, отдающего электроны на хиноны, можно считать циклическим. Но ведь хлороби и гелиобактерии могут отдавать электроны не только на ферредоксин, но и на хиноны, так что, выходит, чёткого разделения нет.
-
Ну, не знаю. ИМХО, разница есть. У зеленых серных бактерий электроны загоняются на NADPH (и ферредоксины, если я правильно понимаю) исключительно силой света. У зеленых несерных восстановление NADP происходит с помощью протонного градиента, не важно, как он возник. Но и у тех, и у других, думаю, электроны в основном по кругу бегают.
-
Ну в этом смысле да, разница есть, но не уверен, что она принципиальна. И понятно, что 100% циклический оборот электронов невозможен, они всё равно будут постепенно где-то теряться, и их поток должен пополняться из других источников, например, отбираться у каких-то органических молекул. Так что, вопрос лишь в степени экономичности цепочки фотосинтеза, соответственно, деление по этому признаку достаточно условно.
-
-
-
-
-
С уважением, автор :-)
-
Хотелось заметить что схематическое изложение структуры реакционного центра который состоит из 13 вполне очевидных молекул. (2 хлорофилла, 4 цитохрома и т.п.) украсило бы статью, потому что "красивые завитушки" на Рис.3 ничего не говорят о то как ловится фотон как передается по цепи электрон и т.д.
-
Поддерживаю! Рис. 3 не вяжется с описанием фотосистемы II в Википедии, и английской и русской. А русская статья аж оценена как избранная.
ru.wikipedia.org/wiki/Фотосистема_II
Где в этой статье упомянуто что‑нибудь подобное комплексу LH2 ? Такого там нет?
С другой стороны ясно, что не стоит перегружать обсуждаемую заметку разъяснением матчасти. Как же быть? Быть может, дописать в Википедию?-
Тут возникает некоторая путаница из за смешения понятий реакционные центры первого и ворого типов и фотосистемы первого и второго типов. Это разные понятия. Название фотосистема (1-го или 2-го типа) обычно применяется к цианобактериям и пластидам. А реакционные центры 1-го и 2-го типов используется для любых фотосинтетиков на основе хлорофилла. Насколько вижу, автор в пояснениях к Рис. 3, иллюстрирующих систему фотосинтеза на основе реакционного центра второго типа, имеющегося у пурпурных бактрий, нигде не использует термина "фотосистема", так что, я лично противоречия здесь не вижу.
-
Ясно, хорошо.
Но по‑прежнему довольно сложный реакционный центр описан на рис. 3 поверхностно. И не имеет подробного описания нигде, в том числе в:
en.wikipedia.org/wiki/Photosynthetic_reaction_centre#In_bacteria
Это контрастирует с подробной википедийной статьёй «Фотосистема II».-
Но по‑прежнему довольно сложный реакционный центр описан на рис. 3 поверхностно. И не имеет подробного описания нигде, в том числе в:
Само собой, контрастирует, потому что у материалов разные задачи. Википедия – это энциклопедия, а здесь у нас научно-популярная новость. В её рамки не входит подробный экскурс – наоборот, он максимально краткий. Предполагается, что за подробностями читатель обратится к соответствующим источникам, при необходимости автор и редакция на них ссылаются. Я вижу, что Вы обратились к статье в Википедии и почитали её. Моя задача как популяризатора науки выполнены :-) А делать из новости энциклопедическую статью нет смысла.
en.wikipedia.org/wiki/Photosynthetic_reaction_centre#In_bacteria
Это контрастирует с подробной википедийной статьёй «Фотосистема II».-
при необходимости автор и редакция на них ссылаются
Но ведь нет такой ссылки для рис. 3. Вероятно, коллега olegov поднял эту тему именно по такой причине — попытался найти подробности про фотосистему пурпурных бактерий и не нашёл их.
Не хочу сказать, что подробное описание требуется в новости. Просто обращаю внимание, что подробного описания в Википедии нет. Возможно, его нигде нет.
-
-
-
-
-
Где в этой статье упомянуто что‑нибудь подобное комплексу LH2 ?
"Фотосистема I" и "фотосистема II" мы говорим только про цианобактерий и фотосинтезирующих эукариот. И у них антенные комплексы устроены совершенно по-другому, нежели у пурпурных бактерий. Поэтому и искать комплекс LH2 в упомянутой википедийной статье нет смысла. Это всё равно что искать упоминание рогов и копыт в википедийной статье про морских червей-сипункулид.
-
Последние новости













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Рис. 1. Когда-то давным-давно пурпурная бактерия «поделилась» своей фотосистемой с бактерией другого вида при помощи горизонтального переноса генов. Новый обладатель настолько удачно ее усовершенствовал, что его вариант фотосистемы стал гораздо эффективнее своего прототипа. Рисунок © Георгий Куракин