Новый вид цианобактерий проливает свет на эволюцию кислородного фотосинтеза

Из растущего в Панаме мха выделен новый вид цианобактерий, названный Anthocerotibacter panamensis. На сегодняшний день у A. panamensis самое простое строение и самый простой фотосинтетический аппарат среди известных цианобактерий. Благодаря этому ученые смогли многое понять про историю появления кислородного фотосинтеза, преобразившего нашу планету. В частности, теперь мы знаем, что у общего предка цианобактерий еще не было тилакоидов (структур, внутри которых происходит фотосинтез), а также представляем себе минимальный набор белков, необходимых для кислородного фотосинтеза.
Монстры, преобразившие мир
Представьте, что на Земле появился новый вид бактерий, который в качестве побочного продукта выделяет ядовитый газ хлор. Из-за них в короткий период времени атмосфера наполняется хлором. Небо из голубого становится желто-зеленым. Сначала гибнут люди и другие позвоночные, позже, по мере роста концентрации токсичного газа, Земля лишается растений и насекомых. Лишь некоторые бактерии успевают приспособиться к ядовитому газу и научиться использовать его для получения энергии. Через миллионы лет они дадут начало новым формам жизни на Земле. Это выглядит как апокалиптическая сцена из фильма-катастрофы, но похожее событие на самом деле произошло на Земле около двух с половиной миллиардов лет назад. За исключением того факта, что жизнь на тот момент была представлена только бактериями и археями — эукариоты еще не успели возникнуть.
Около двух с половиной миллиардов лет назад появились цианобактерии — группа бактерий, которая научилась проводить фотосинтез, используя в качестве донора электронов обычную воду (напомню, что как при дыхании, так и при фотосинтезе поток электронов от одного белкового комплекса к другому непосредственно дает энергию для перекачки протонов и создания протонного градиента). К тому времени фотосинтез уже давно существовал, но он требовал неорганического «сырья» типа сероводорода, которое еще надо найти. Цианобактерии в этом плане оказались в явном выигрыше: воды вокруг было в буквальном смысле залейся, солнечного света и углекислого газа тоже хватало, так что необходимые им ресурсы оказались практически ничем не ограниченными. Благодаря этому их численность резко подскочила. Но у цианобактерий был один важный недостаток: в процессе такого выгодного фотосинтеза в качестве отхода выделялся токсичный кислород, получающийся при расщеплении воды.

Рис. 2. Мы часто видим цианобактерии — макроскопически они выглядят как «тина» или «ряска» в пруду. Многие цианобактерии почти не изменились за более чем 2 млрд лет своей эволюции. Так что те бактерии, которые изменили облик Земли, вполне могли выглядеть так же. Фото © Stefe с сайта flickr.com
Если бы злой рок забросил вас примерно на 2,9 миллиарда лет назад, то вы бы задохнулись. В тогдашней атмосфере почти не было кислорода. В ней преобладали углекислый газ, азот и метан — в общем, все то, что за предыдущие полтора миллиарда лет успели произвести вулканы. С химической точки зрения такая атмосфера создавала восстановительную среду, то есть в целом была склонна отдавать электроны, а не отнимать их. Как ни странно, это оказалось на руку зарождающейся жизни: вся органика очень уязвима к окислителям, и восстановительная атмосфера была поистине комфортной колыбелью для жизни. Это была оптимальная среда для первых обитателей Земли.
Массивное выделение кислорода цианобактериями произвело такой же эффект, как если бы сейчас атмосфера наполнилась хлором. В ряду электроотрицательности неметаллов кислород на втором месте, а хлор — только на четвертом. Поэтому кислород оказывал на тогдашнюю жизнь такой же негативный эффект, как хлор — на современную. Токсичность обоих газов заключается в их высочайшей окислительной способности, к которой чрезвычайно чувствительна вся земная жизнь: она построена из органических макромолекул, легко повреждающихся сильными окислителями.
Мы сейчас не замечаем пагубного влияния кислорода лишь потому, что эволюционно приспособились к нему. Точнее, к нему приспособились наши предки. Помимо того, что мы утилизируем его при дыхании, мы вооружены множеством ферментов и антиоксидантов для нейтрализации активных форм кислорода. Наши клетки не замечают этого яда лишь потому, что постоянно «сражаются» с ним и используют в своих целях. В общем, цианобактерии своим кислородом полностью преобразили биосферу и метаболизм многих живых существ, совершив настоящую «кислородную революцию».
Помимо цианобактерий, оксигенный фотосинтез могут осуществлять растения и многие группы водорослей. Но все они осуществляют его при помощи пластид, которые происходят либо от поглощенных когда-то цианобактерий, либо... от других фотосинтезирующих эукариот, имевших пластиды, произошедшие от цианобактерий. Как пишет Александр Марков в книге «Рождение сложности»: «Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные „домики“ для проживания цианобактерий. По сути дела цианобактерии не только изобрели кислородный фотосинтез, но и по сей день сохранили за собой „эксклюзивные права“ на его осуществление».
Если представить, какую роль растения и водоросли играют в современном облике нашей планеты, то станет ясно, что мы живем в мире, сформированном цианобактериями.
Цианобактерии «придумали» не только фотосинтез. Если посмотреть на цианобактерии под микроскопом, в глаза бросится еще одно их отличие: среди них много многоклеточных видов, существующих в виде нитей из многих клеток (среди эукариот аналогами такой жизненной формы являются нитчатые водоросли). Причем речь идет именно о настоящей многоклеточности: клетки в составе нити обмениваются метаболитами и сигнальными молекулами. А у наиболее «продвинутых» цианобактерий (например, представителей рода Nostoc) существует дифференцировка на разные типы клеток: одни клетки (вегетативные) фотосинтезируют, а другие (гетероцисты) занимаются фиксацией азота.
Молекулярно-филогенетические исследования показывают, что многоклеточность у цианобактерий — настолько же древнее явление, как и сам кислородный фотосинтез (B. E. Schirrmeister et al., 2013. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event). Это подтверждают и окаменелости, удивительно похожие на современные формы многоклеточных цианобактерий (см. статью Сергея Ястребова Кислородная революция и Земля-снежок). Можно уверенно утверждать, что цианобактерии точно находятся в ряду первых «изобретателей» многоклеточности. Более того, есть предположение, что какими-то молекулярными «находками» для поддержания межклеточной кооперации они поделились с нами. Есть гипотеза, что именно они изобрели каспазы — белки запрограммированной клеточной гибели, — и далее все остальные многоклеточные организмы позаимствовали эти белки у них (N. Lane, 2008. Marine microbiology: Origins of Death). Также не исключено, что у цианобактерий мы «научились» синтезировать оксилипины (см. Oxylipin) — универсальные молекулы межклеточной сигнализации (см. Зачем бактериям и простейшим липоксигеназы?, «Элементы», 31.10.2020). Впрочем, это пока только гипотеза.
Цианобактериальные инновации
Такой власти над миром цианобактериям удалось добиться лишь потому, что они научились при фотосинтезе расщеплять воду вместо сероводорода. А сделать это не так просто — вновь из-за того, что кислород является сильным окислителем, и высвободить его из воды труднее, чем серу из сероводорода (чем сильнее окислитель, тем труднее он сам окисляется). Чтобы выпустить в атмосферу такой химически активный газ, отняв у него электроны, одной фотосистемы (электрон-транспортной цепи фотосинтеза) недостаточно. История цианобактерий началась с того, что в результате горизонтального переноса генов они смогли объединить в себе фотосистемы двух «образцов», существовавшие у пурпурных бактерий и зеленых серобактерий. Вместе они развивают гораздо больший оксилительно-восстановительный потенциал, чем по отдельности, что и позволяет отрывать электроны от воды. В целом можно сказать, что объединение двух фотосистем позволило цианобактериям высвобождать в атмосферу мощный окислитель. И это же привело к катастрофе в биосфере.
Большинство известных на сегодняшний день цианобактерий имеют весьма совершенный фотосинтетический аппарат — не только с точки зрения биохимии, но и с точки зрения структуры. Цепь переноса электронов, задействованная в фотосинтезе, очень похожа на дыхательную цепь, но в митохондриях и хлоропластах (а также у аэробных бактерий и большинства цианобактерий) эти цепи располагаются в разных местах. У митохондрий (рис. 3) они встроены во внутреннюю мембрану, которая происходит от цитоплазматической мембраны когда-то поглощенного нашими далекими предками бактериального симбионта. А у бактерий дыхательная цепь находится в самой цитоплазматической мембране. Цепи переноса электронов выкачивают протоны на внешнюю сторону мембраны, наполняя ими межмембранное пространство. Получившийся градиент концентрации протонов в буквальном смысле вращает АТФ-синтазу, «чеканящую» универсальную энергетическую валюту клетки — АТФ.

Рис. 3. У митохондрий, анаэробных бактерий, хлоропластов и фотосинтезирующих бактерий похожие электрон-транспортные цепи, создающие протонный градиент на мембране. Только в первых двух случаях они «работают» на энергии окислительно-восстановительных реакций, а в двух последних случаях — на энергии солнечного света. Но их локализация причудливым (на первый взгляд) образом различается. У митохондрий электрон-транспортные цепи располагаются на внутренней мембране, а протоны закачиваются в межмембранное пространство. У аэробных бактерий они располагаются на цитоплазматической мембране (то есть собственно клеточной мембране), эволюционно соответствующей внутренней мембране митохондрий. У хлоропластов и «кроновых» цианобактерий внутри есть дополнительный компартмент (отсек), состоящий из системы дисков и пластинок — тилакоидов. Именно на их мембранах располагаются фотосистемы, перекачивающие протоны внутрь тилакоидов. Это создает внутри высокую концентрацию протонов и мощный протонный градиент. Но у глеобактерии G. violaceus (и у главного «героя» нашей статьи Anthrocerotibacter panamensis) нет никаких тилакоидов, а фотосистемы расположены на цитоплазматической мембране и работают по тому же принципу, что и дыхательные цепи митохондрий и аэробных бактерий. Похоже, так устроены все глеобактерии. Рисунок © Георгий Куракин
А в хлоропластах и у большинства цианобактерий имеется замкнутая система мембранных дисков и пластинок, отделенных от внутренней мембраны — тилакоидов. Именно на мембранах тилакоидов размещаются электрон-транспортные цепи фотосинтеза, использующие энергию солнечного света для закачивания протонов внутрь тилакоидов. Их плоская форма оптимальна для создания минимального объема внутри при максимальной поверхности, позволяющей разместить максимальное число электрон-транспортных комплексов. Такая конструкция концентрирует протоны куда более эффективно, чем внутренняя мембрана митохондрий, позволяя синтезировать АТФ куда более эффективно, чем это происходит при дыхании.
Помимо эффективного механизма концентрации протонов, современные цианобактерии и растения имеют мощные молекулярные «антенны», улавливающие свет и передающие его молекулам хлорофилла. У растений это светособирающие комплексы, а у цианобактерий — фикобилисомы (рис. 4).
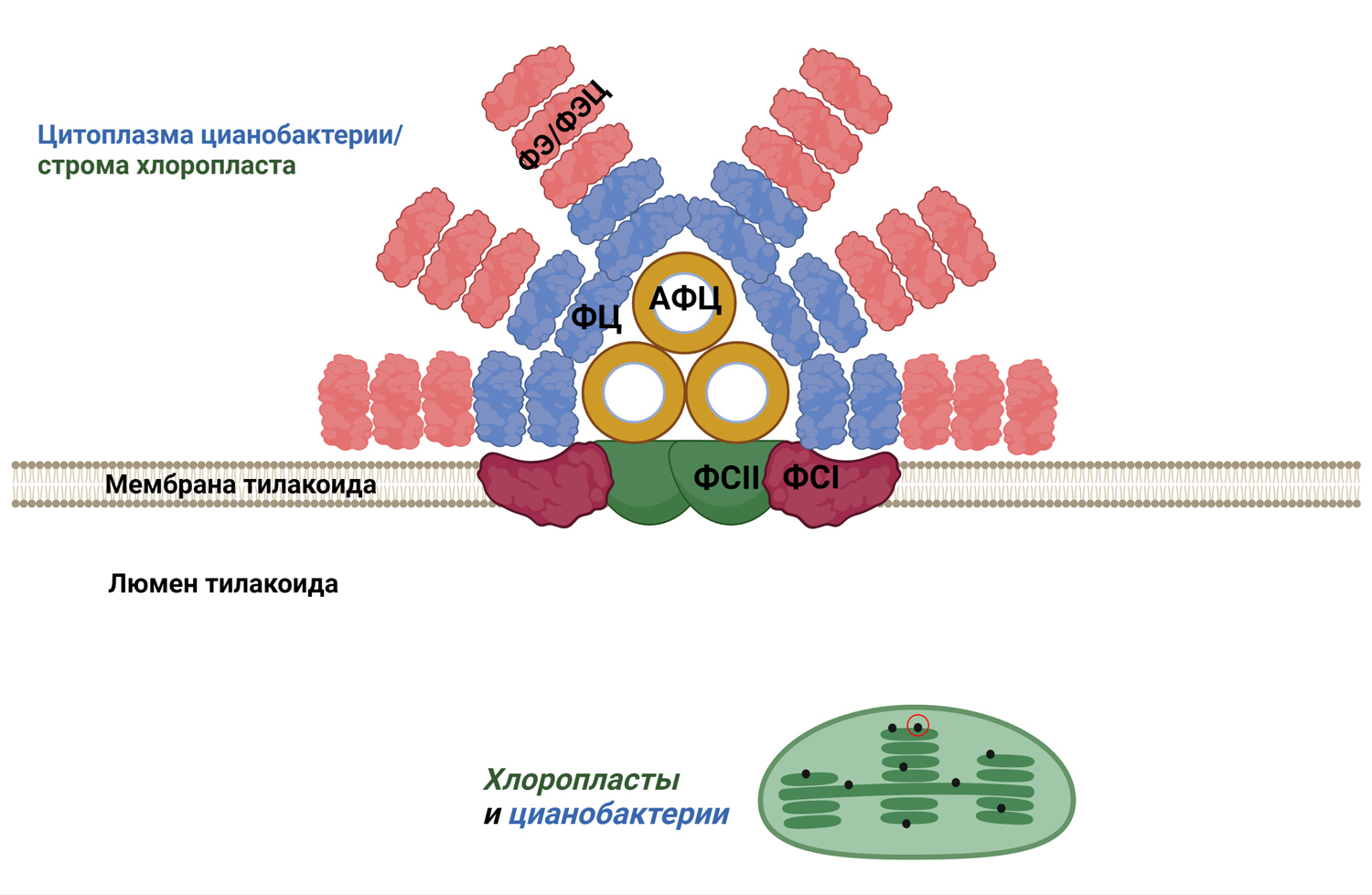
Рис. 4. У большинства цианобактерий и в хлоропластах красных водорослей над каждым комплексом из двух фотосистем, образующих электрон-транспортные цепи фотосинтеза, располагается огромный (по сравнению с ними) светособирающий «локатор» из белков — так называемая фикобилисома. Любопытно, что растения используют совсем другой светособирающий комплекс вместо фикобилисомы. АФЦ — аллофикоцианин, ФЦ — фикоцианин, ФЭ/ФЭЦ — фикоэритрин или фикоэритроцианин. Рисунок © Георгий Куракин
Белки фикобилисомы содержат в качестве небелковой части фикобилины (похожие на наш билирубин), которые непосредственно «собирают» кванты света. Аналогичные вещества используют растения — только у них светособирающий комплекс устроен по-другому.
Чтобы эффективно использовать свет, цианобактериям пришлось обзавестись и другой инновацией. Как известно, у многих многоклеточных организмов — как растений, так и животных — независимо развились сенсоры дня и ночи и регуляция ритмов активности в зависимости от времени суток, называемых циркадными (циркадианными) ритмами.
У растений за это отвечают криптохромы — особые белки, позволяющие растению чувствовать свет. Хоть растения и не способны видеть, но свет от тьмы они «отличают». У животных, в том числе у человека, криптохромы тоже задействованы в регуляции циркадных ритмов (они находятся в ядрах гипоталамуса, регулирующих цикл сна и бодрствования).
Большинству бактерий такие механизмы без надобности, но цианобактериям они жизненно необходимы: их биохимия целиком и полностью зависит от солнечного света. Поэтому у них есть внутренние «биологические часы», подсказывающие, когда какие реакции проводить. Они состоят из трех белков (не родственных криптохромам), вступающих в циклические биохимические реакции и тем самым образующих своеобразный «молекулярный маятник» (хотя в хронобиологии подобные системы называются осцилляторами). Что интересно, эту систему можно вживить в любую другую бактерию — и она тоже будет осуществлять те или иные биохимические функции «по часам» (см. В бактерию без циркадных ритмов встроили «часы» от цианобактерии, «Элементы», 15.06.2015).
Наличие «биологических часов» тоже позволяет большинству известных цианобактерий эффективно использовать солнечный свет. В общем, у цианобактерий есть все, чтобы в «промышленных» масштабах разлагать воду и производить сильный окислитель — кислород. Возникает вопрос: а как они пришли к такому состоянию? Как способность к фотосинтезу развивалась в ходе эволюции?
Древо жизни цианобактерий
Ближайших «родственников» цианобактерий мы можем обнаружить... у себя в кишечнике, а также в пресных водоемах. Они называются мелайнабактериями (Melainabacteria), что в буквальном переводе с греческого означает «черные бактерии» (в противоположность цианобактериям — «лазурным бактериям»). Это название весьма говорящее: никто из мелайнабактерий не умеет осуществлять фотосинтез. Оксигенный фотосинтез, очевидно, появился после разделения цианобактерий и мелайнабактерий. Ясности в вопросе, как развился оксигенный фотосинтез, это не прибавляет.
Но сами цианобактерии — не однородная группа. На ее филогенетическом дереве выделяется базальная (то есть отходящая раньше всего) ветвь — так называемые глеобактерии (Gloeobacteria, рис. 5). Все остальные цианобактерии выделяются в группу Phycobacteria, но чаще всего в англоязычной литературе встречается наименование «кроновые цианобактерии» (crown cyanobacteria) — из-за того, что именно они образуют «крону» филогенетического дерева цианобактерий.

Рис. 5. Эволюционное дерево цианобактерий в упрощенном виде. Звездочкой отмечен «главный герой» этой статьи — Anthocerotibacter panamensis. Рисунок © Георгий Куракин, использованы изображения с сайта acssi.org и из статей R. N. Prasad et al., 2013. Isolation, Identification and Germplasm Preservation of Different Native Spirulina Species from Western Mexico, J. H. W. Saw et al., 2013. Cultivation and Complete Genome Sequencing of Gloeobacter kilaueensis sp. nov., from a Lava Cave in Kīlauea Caldera, Hawai'i, B. T. Hovde et al., 2019. Vampirovibrio chlorellavorus draft genome sequence, annotation, and preliminary characterization of pathogenicity determinants
В изучении эволюции цианобактерий глеобактерии представляют собой такую же ценность, как ланцетники при изучении эволюции хордовых (см. Функциональный анализ генома ланцетника проливает новый свет на эволюцию позвоночных, «Элементы», 26.11.2018). Это наиболее рано обособившиеся представители, по которым легче всего попытаться реконструировать эволюционные изменения, приведшие к появлению целой новой группы. Глеобактерии на данный момент — практически единственное доступное нам звено, переходное от нефотосинтезирующих мелайнабактерий к мастерам высокоэффективного кислородного фотосинтеза. И практически единственный наш шанс ответить на вопрос: как появились эти монстры, испортившие земную атмосферу 2,5 млрд лет назад?
Проблема в том, что доступных данных по глеобактериям до настоящего времени было мало — всего два вида были выращены в культуре и описаны экспериментально: Gloeobacter violaceus и Gloeobacter kilaueensis. G. violaceus стал основным модельным организмом для изучения эволюции оксигенного фотосинтеза. На поверку он оказался устроен гораздо проще «кроновых» цианобактерий. Во-первых, он одноклеточный и не имеет циркадных ритмов. Во-вторых, у него отсутствуют тилакоиды. Если фотосистемы «кроновых» цианобактерий и фотоавтотрофных эукариот «закачивают» протоны в люмен (просвет) тилакоидов, то у G. violaceus они просто выкачиваются на внешнюю сторону цитоплазматической мембраны, в межмембранное пространство (см. рис. 3). В итоге способом создания протонного градиента G. violaceus больше всего напоминает не «кроновые» цианобактерии и хлоропласты, а наши митохондрии и их альфа-протеобактериальных предшественников — у них протонный градиент генерируется так же.
Таким образом, общий план строения клетки G. violaceus очевидно проще, чем у «кроновых» цианобактерий. Помимо этого, у G. violaceus фикобилисомы устроены проще, чем у других описанных цианобактерий (об этом позже) и отсутствуют некоторые компоненты фотосистем. Что, казалось бы, интуитивно ясно — можно ожидать, что рано «ответвившийся» представитель цианобактерий будет более примитивным.
Есть у него и еще одно биохимическое отличие, связанное с биосинтезом каротиноидов. Каротиноиды — это те самые желтые, красные и оранжевые пигменты, которые расширяют спектральный диапазон поглощаемого света и помогают более эффективно использовать его энергию при фотосинтезе. Но каротиноиды умеют синтезировать не только растения и цианобактерии. Они также синтезируются многими нефотосинтезирующими бактериями и грибами. Но при этом в природе существует два способа синтезировать каротиноиды — два варианта биохимического пути их синтеза (рис. 6). Различия между этими путями касаются одного «участка» — введения в молекулу фитоена (phytoene) дополнительных двойных связей, что превращает его в ликопин. В «растительном» пути, используемом растениями и большинством цианобактерий, это превращение осуществляется в три стадии тремя разными ферментами-десатуразами. А в «бактериальном» пути, используемом грибами и нефотосинтезирующими бактериями, это превращение протекает в одну стадию и катализируется одной десатуразой.
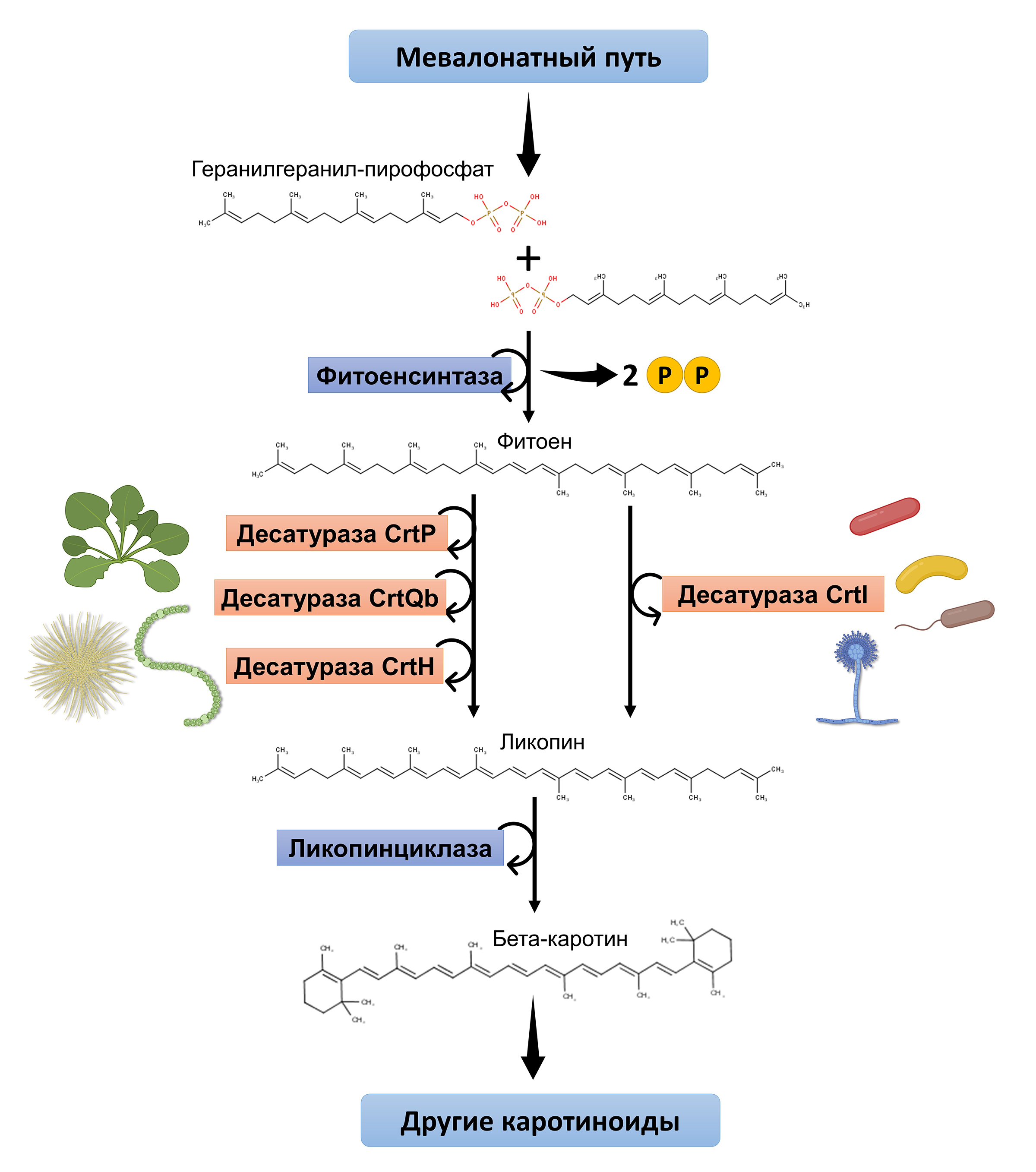
Рис. 6. Биосинтез каротиноидов начинается по мевалонатному пути — у человека этот биосинтетический путь задействован в синтезе холестерина. Но в биосинтезе каротиноидов вместо стероида из изопреновых звеньев (как из кубиков или вагончиков игрушечного поезда) собирается длинная молекула фитоена. Но в ней еще слишком много предельных (одинарных) связей «углерод-углерод», которые должны превратиться в непредельные (двойные). Именно эта стадия и различается в двух существующих в живой природе путях: в «растительном» пути за нее «отвечают» три фермента, а в «бактериальном» — всего один. Дальше пути опять «объединяются»: концы молекулы замыкаются в цикл с образованием уже знакомого скелета каротиноида. Рисунок © Георгий Куракин, использованы материалы из статьи S. Steiger et al., 2005. Carotenoid biosynthesis in Gloeobacter violaceus PCC4721 involves a single crtI-type phytoene desaturase instead of typical cyanobacterial enzymes
Как оказалось, G. violaceus использует «бактериальный» тип биосинтеза каротиноидов, соответствующий нефотосинтезирующим бактериям. Возникает интуитивный соблазн тоже отнести эту особенность к проявлениям «примитивизма».
Но выводы о «примитивном» или «изначальном» состоянии в данном случае являлись бы очень спекулятивными и поспешными. В самом деле, какие-то из перечисленных особенностей могли развиться у G. violaсeus уже после его обособления от других цианобактерий, — то есть он мог претерпеть что-то вроде «эволюционной дегенерации» (если говорить совсем грубо). Тем самым не исключено, что общий предок всех цианобактерий уже имел и тилакоиды, и биосинтез каротиноидов «растительного» типа, а глеобактер просто их утратил. Двух видов было абсолютно недостаточно, чтобы делать далеко идущие эволюционные выводы. Требовалось выделить и охарактеризовать какие-нибудь еще из глеобактерий — и желательно не из рода Gloeobacter.
Откуда не ждали
Новый вид был открыт неожиданно. Открыла его группа под руководством Фэй-Вэя Ли (Fai-Wei Li), сотрудника Института Бойса Томпсона (входящего в Корнеллский университет), которая занимается выделением цианобактериальных симбионтов из антоцеротовых мхов. У любого из этих мхов есть какой-нибудь бактериальный симбионт из числа многоклеточных цианобактерий, осуществляющий для мха фиксацию атмосферного азота (так же, как клубеньковые бактерии снабжают фиксированным азотом растения семейства Бобовые). Этих-то симбионтов и искали исследователи. И очень удивились, когда из мха Leiosporoceros dussii (рис. 7) вместо нитчатой цианобактерии выделилась одноклеточная глеобактерия. Это была редкая удача, но тем не менее она была неожиданной.

Рис. 7. Антоцеротовый мох Leiosporoceros dussii, из которого выделили новый вид цианобактерий: слева — фото с сайта stri.si.edu, справа — в художественном воплощении иллюстратора Анастасии Самоукиной
Это было тем более неожиданно, что новый вид, названный Anthocerotibacter panamensis (рис. 1 и рис. 8; буквально это название означает «бактерия антоцеротового мха из Панамы») никак не годился на роль азотфиксирующего: у него нет гена, необходимого для фермента, отвечающего за фиксацию азота. Такого симбионта мху держать незачем, так что непонятно, как он оказался в его талломе. Авторы выдвинули два предположения: или Anthocerotibacter panamensis случайно занял «пустующее» место постоянного симбионта (который, скорее всего, существует, просто не был обнаружен в этом образце), или только он выдержал процедуру поверхностной стерилизации, применяемую авторами для удаления бактерий, попавших из почвы.

Рис. 8. Новооткрытый вид Anthocerotibacter panamensis под микроскопом и без окрашивания. Хорошо видно, что одноклеточная цианобактерия палочковидной формы. Изображения предоставлены Fay-Wei Li
Еще одним неожиданным моментом стало то, что Anthocerotibacter panamensis оказался ближайшим родственником не выделенных пока глеобактерий, обитающих в Арктике и филогенетически входящих в так называемую «полярную кладу». Ведь сам-то A. panamensis происходит из тропиков (а конкретно — из Панамы, о чем и говорит его название). Возможно, что «полярная клада» вовсе не такая уж полярная, а возможно, что новый вид действительно просто экзотический.
Говоря «не выделенных пока глеобактерий», я имел в виду, что их пока никто не видел под микроскопом и не вырастил в культуре. А откуда мы тогда знаем, что бактерии «полярной клады» существуют? Это известно из метагеномного исследования (C. L. Grettenberger et al., 2020. A phylogenetically novel cyanobacterium most closely related to Gloeobacter). В ходе таких исследований секвенируется вся ДНК, найденная в каком-нибудь образце почвы или воды. Секвенирование и последующий биоинформатический анализ могут с некоторой степенью надежности воссоздать геномы еще не выделенных в культуре бактерий, оказавшихся в образце.
Новый вид оказался во многом похож на глеобактера. Он также одноклеточный, не имеет циркадных ритмов и тилакоидов. Это надежно указывает на то, что многоклеточность, «биологические часы» и тилакоиды появились у общего предка «кроновых» цианобактерий, а общий предок всех цианобактерий их еще не имел. Очевидно, он при фотосинтезе использовал ту же менее эффективную схему, когда протонный градиент создается на мембране клетки, а способ повысить напряжение за счет тилакоидов был более поздним приобретением. «Теперь мы можем быть почти уверены, что тилакоиды развились у фикобактерий» — говорит Фэй-Вэй Ли. Как и следовало ожидать, в плане отсутствия тилакоидов глеобактерии представляют собой сохранившийся реликт, самое примитивное состояние. Но так произошло не со всеми признаками.
С биосинтезом каротиноидов все получилось совсем наоборот, чем можно было бы предполагать. Оказалось, что у A. panamensis каротиноиды синтезируются по «растительному» пути, как и у наиболее «продвинутых» цианобактерий. Это позволяет предположить, что этот путь сформировался уже у общего предка всех цианобактерий — и лишь у некоторых глеобактерий, включая G. violaceus, он потом заменился на «бактериальный». Такой пример вторичного «упрощения», если так можно выразиться.
Фотосинтезирует, но плохо
Но самые интересные находки касались фотосинтеза. У нового вида фотосинтетический аппарат оказался самым примитивным из известных на сегодняшний день. Прежде всего, у него самые простые фикобилисомы. Вспомним, что у «кроновых» цианобактерий и некоторых фотосинтезирующих эукариот фикобилисомы имеют вид антенны-локатора или вентилятора (рис. 9, сверху). Основание антенны образовано белком аллофикоцианином (АФЦ), а «лепестки» построены из фикоцианина (ФЦ) и фикоэритрина (ФЭ). Иногда место последнего занимает фикоэритроцианин (ФЭЦ). Но у G. violaceus фикобилисома проще и имеет вид стопки из белков, или, если изображать упрощенно, столбика (рис. 9, в середине). Таким образом, если сравнивать фикобилисомы с техническими сооружениями, то у «кроновых» цианобактерий имеется тарелка локатора, а у глеобактера — что-то вроде телевизионных антенн, которые можно встретить на крышах домов. Но состоит антенна из тех же белков: АФЦ, ФЦ и ФЭ.
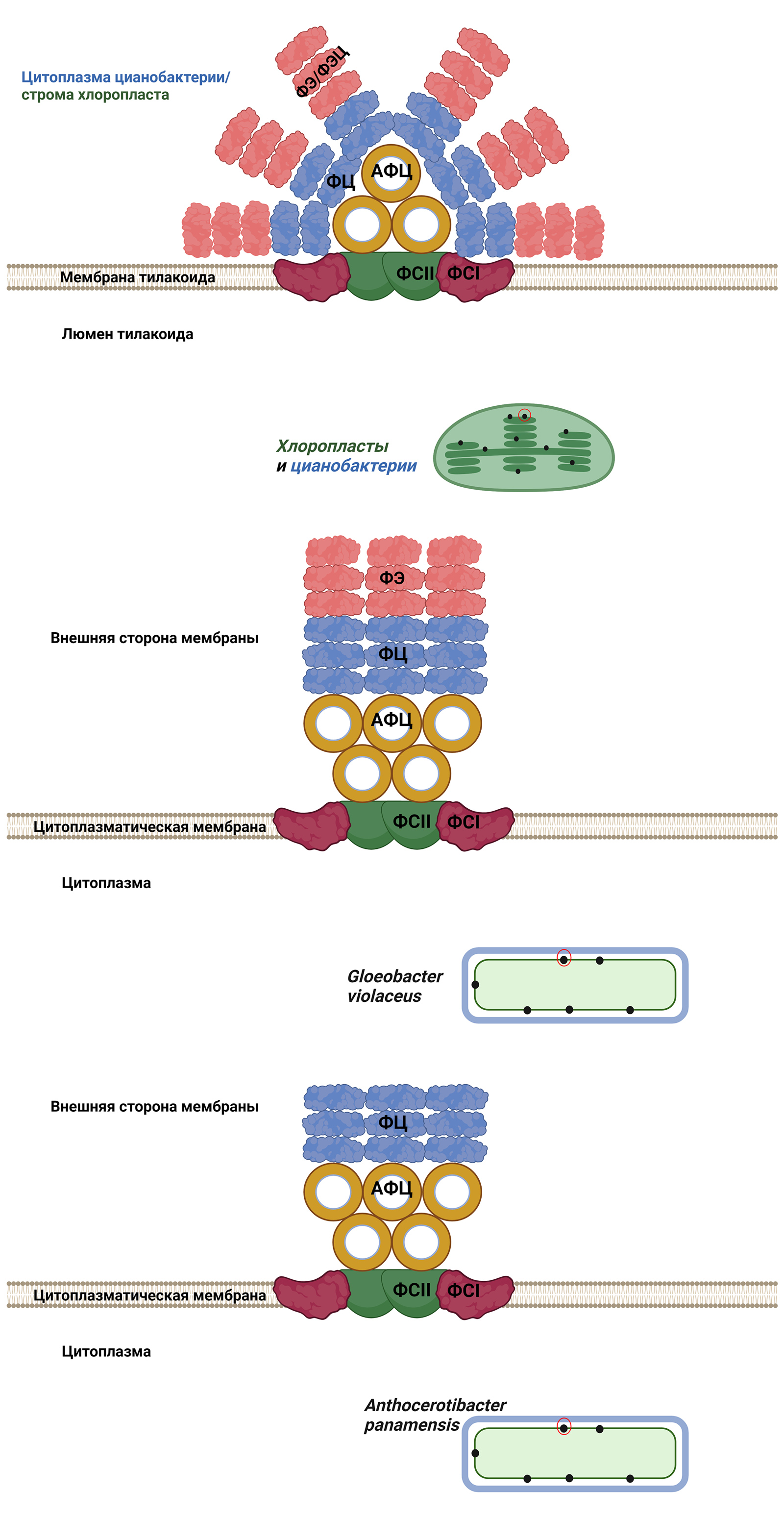
Рис. 9. Вверху — уже знакомая читателю фикобилисома «кроновых» цианобактерий и красных водорослей (дана для сравнения). Фикобилисома глеобактера состоит из тех же белков, но в этом случае они образуют антенну-«столбик» вместо антенны-«тарелки». Предположительное строение фикобилисомы антоцеротибактера. В ней отсутствует фикоэритрин, что делает ее упрощенной даже по сравнению с фикобилисомой глеобактера. Рисунок © Георгий Куракин
Структура фикобилисомы A. panamensis пока экспериментально не определена, так что говорить о ней (и пытаться ее нарисовать!) можно лишь предположительно. Одно мы знаем точно — она проще, чем даже у глеобактера: в ней отсутствует фикоэритрин. Таким образом, фикобилисома антроцеротибактера пока что претендует на звание самой простой и примитивной по белковому составу. Внизу на рис. 9 я нарисовал, как это могло бы выглядеть, если форма фикобилисомы антоцеротибактера такая же, как у глеобактера. Продолжая наши аналогии, получившуюся конструкцию можно сравнить с комнатной телевизионной антенной, — настолько она упрощена. Но, повторюсь, формы фикобилисомы глеобактера мы пока не знаем. И это лишь делает более интригующим вопрос: как выглядит самая простая фикобилисома на свете?
Впрочем, у нового вида не только фикобилисомы устроены максимально просто. Сами фотосистемы — «рабочий механизм» фотосинтеза — у него тоже упрощены (рис. 10). В фотосистеме I отсутствует треть всех белков, из которых строится та же фотосистема у «кроновых» цианобактерий. Правда, у глеобактера фотосистема I устроена, по всей видимости, так же. А вот по части фотосистемы II антоцеротибактер пока удерживает безусловный рекорд простоты: у него отсутствует пять из 23 белков (у G. violaceus нет четырех белков, у G. kilaueensis — трех). Подобная примитивность заставляет биохимиков еще раз задуматься над вопросом, каков минимальный «набор» белков, необходимых для оксигенного фотосинтеза? Это представляет большой интерес для синтетической биологии.

Рис. 10. Мы пока не можем напрямую увидеть структуры фотосистем антроцеротибактера, но можем взять структуры фотосистем «кроновых» цианобактерий и отметить (красным) те компоненты, которых нет у нового вида цианобактерий. Компоненты, присутствующие у него, отмечены зеленым. «Кубики» — это железо-серные кластеры, оранжевые шарики — марганцевый водоокисляющий кластер. Слева — фотосистема I, рисунок создан на основе фотосистемы I цианобактерии Synechococcus elongatus (PDB ID: 1jb0); справа — фотосистема II, рисунок на основе фотосистемы II цианобактерии Thermosynechococcus elongatus (PDB ID: 2axt). Рисунок © Георгий Куракин
Не менее интересным для исследователей оказался вопрос: а насколько эффективный фотосинтез способна производить бактерия с таким примитивным фотосинтетическим аппаратом? Для этого ученые протестировали скорость роста нового вида в отсутствие органических питательных веществ и при наличии одного лишь света, а также определили скорость выработки кислорода. Оказалось, что, «питаясь» одним светом, новая цианобактерия растет очень медленно: почти трое суток ей требуется на одно деление, даже если свет яркий. При более тусклом свете она делится и вовсе раз в неделю. Это гораздо медленнее, чем у «кроновых» цианобактерий (их представитель Synechocystis sp. PCC 6803 делится каждые 8 часов) и сопоставимо с одним из рекордсменов медленного роста совсем из другой «оперы» — микобактерией туберкулеза. Это показывает, насколько неэффективен фотосинтетической аппарат антоцеротибактера. Но при этом скорость выделения кислорода была лишь примерно вдвое ниже, чем у «кроновой» цианобактерии Synechocystis sp. PCC 6803. То есть, если бы антроцеротибактер 2,5 млрд лет назад «работал» в одиночку, он все же имел бы шансы произвести кислородную революцию и преобразить мир. Только, может быть, на полмиллиарда лет позже.
Рисунки созданы с помощью biorender.com.
Источник: Nasim Rahmatpour, Duncan A. Hauser, Jessica M. Nelson, Pa Yu Chen, Juan Carlos Villarreal A., Ming-Yang Ho, Fay-Wei Li. A novel thylakoid-less isolate fills a billion-year gap in the evolution of Cyanobacteria // Current Biology. 2021. DOI: 10.1016/j.cub.2021.04.042.
Георгий Куракин
-
Белки фикобилисомы содержат в качестве небелковой части каротиноиды,
Кажется, тут ошибка - они содержат совсем другие пигменты, фикобилины, похожие на порфирин с разомкнутым кольцом.-
Любопытно, что фикобилины при этом синтезируются из гема b. Вначале замыкается порфириновое кольцо, туда вставляется атом железа (получаем гем b), после чего это железо вынимается обратно, а кольцо разрезается. Чудны дела твои, слепой часовщик! ))
-
Фикобилины растений – это химические аналоги нашего билирубина (о чём я и написал в поправке – не про "разомкнутый порфирин", как @HellMaus рекомендует, а именно про билирубин).
В этом плане любопытно, что биосинтез фикобилинов у растений повторяет "биосинтез" билирубина у человека – у человека билирубин синтезируется из распадающегося гема. То есть мне как медику по образованию этот биосинтетический путь не то что понятен, а до боли знаком – он протекает как у человека. Видимо, это очень консервативный путь, который. Я согласен c @olegov, что он просто сохранился как реликт. Вопрос – реликт чего? Но эту уже тема отдельного исследования!


каков минимальный «набор» белков, необходимых для оксигенного фотосинтеза?наверное тут стоило сравнить с белками зеленых серный и пурпуробактерий?
А если идти обратно то отключать белки по одному и смотреть что выйдет.
Были же такие работы наверное?
Статья интересная спасибо!
-
"наверное тут стоило сравнить с белками зеленых серный и пурпуробактерий"
Кстати, мысль хорошая, можно было бы! Мне в голову не пришло, так как я следовал логике обсуждаемой статьи. А можно было бы и дополнить её немного, учитывая мою привычку писать новости-разборы.
"А если идти обратно то отключать белки по одному и смотреть что выйдет.
Были же такие работы наверное?"
Если отключить у того же ностока даже все те белки, которых нет у глеобактера, носток может вообще не выжить. Всё-таки за миллиарды лет эволюции консервативные субъединицы ФС1 и ФС2 "притёрлись" к "новым" субъединицам, и вряд ли уже смогут работать без них, как у глеобактера. Вряд ли получится такой эксперимент... мне такие примеры неизвестны.
"Статья интересная спасибо!!
Вам спасибо за отзыв!-
А можно было бы и дополнить её немного, учитывая мою привычку писать новости-разборы.
это всегда интереснее чем просто перевод, тем более что авторы сознательно ограничивают данные по статье, даже если владеют данным материалом, в силу особенностей публикаций.
Хотя я думаю что участие сульфида железа в ФС1 тоже совсем неспроста, сульфид этот скорее всего реликт абиогенезного периода, поэтому и отказать от него как и от гликолиза, циклов Кребса и т.п. очень трудно.Вряд ли получится такой эксперимент
согласен, здравое замечание, эксперимент тогда должен был быть модифицирован не прсото удалением белка а заменой его низкоэффективным катализатором белком предшественником, наверняка с этим в итоге начнут работы, направление перспективное.-
Хотя я думаю что участие сульфида железа в ФС1 тоже совсем неспроста, сульфид этот скорее всего реликт абиогенезного периода, поэтому и отказать от него как и от гликолиза, циклов Кребса и т.п. очень трудно.
==================
Я какое-то время назад тоже так думал, но последние данные, как это ни странно, указывают на то, хиноны в качестве переносчиков электронов и протонов при фотосинтезе появились раньше ферредоксинов.-
хиноны в качестве переносчиков электронов
хиноны в природе часто работают вместе с Fe, усиливая друг друга, могу предположить что данные по хинонам должны биться с железом. С одной стороны FeS вот он готовый бери и используй, а хинон еще синтезировать надо и окислитель он неплохой. Хотелось бы понимать для чего и когда они возникли?
-
-
-
Но если бы все было так просто в эволюции фотосинтеза, как думают авторы статьи...Достаточно посмотреть несколько работ (например, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6451369/ , https://www.nature.com/articles/s41467-021-24396-y или вот эту https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1551899/), чтобы стало ясно, как все запутано и как простая логика авторов может привести к выводам, весьма далеким от реальности.
-
Искандар Миртибатович, спасибо большое за комментарий.
Всё зависит от того, как понимать термин "хлоропласты". Ранее (в основном в описательной биологии и ботанике) господствовала традиция называть хлоропластами только фотосинтезирующие органеллы зелёных водорослей и растений. А у красных водорослей их называли родопластами. Сейчас термин "родопласты" либо почти не употребляется, либо считается частным случаем "хлоропластов". И идёт тенденция называть "хлоропластами" все хлорофилл-содержащие пластиды.
Вот как пример журнальная статья:
https://nph.onlinelibrary.wiley.com/doi/10.1111/j.1469-8137.1996.tb01923.x
"Because the thylakoids in red algal chloroplasts are not stacked, freeze-fractured thylakoid membranes exhibit only two types of fracture faces (EF and PF)".
И даже там, где используется общий термин "пластиды", родопласты считаются частным случаем хлорпластов с биохимической точки зрения. Вот, например, глава из книги:
https://link.springer.com/chapter/10.1007/978-1-4020-4061-0_1
"Technically chloroplasts, because they do contain chlorophyll a, rhodoplasts are red to purple, depending on the water depth, which influences the light environment and phycocyanin or phycoerythrin pigment composition (MacColl and Gaurd-Friar, 1987; Cunningham et al., 1989)".
Поэтому и здесь я называю "хлоропластами" пластиды всех фотосинтезирующих эукариот. На рис. 3, 4 и 9 речь идёт о хлоропластах "вообще", обо всей их эволюционной линии, происходящей от эндосимбиоза с цианобактерий (казуистический случай Paulinella sp.) мы тут не рассматриваем. Разумеется, я при этом держал в голове, что в той эволюционной линии хлоропластов, которая разделилась на растения и зелёные водоросли, фикобилисомы были утрачены и заменены другим типом светособирающего комплекса. И поэтому в подписи под рис. 4 указано, что речь идёт о хлоропластах красных водорослей, а растений фикобилисом нет.
Необходимое уточнение я сделал, разница только в терминологии. Но сейчас термин "хлоропласты" применяют ко всем таксономическим группам, у кого они есть. Даже про паулинеллу, по-моему, можно так сказать, хотя вот у неё самые экзотичные хлоропласты из всех существующих на нашей планете.
Думаю, с моими пояснениями в тексте понятно, про кого я говорил, и что у растений фикобилисом нет.
-
открытие Anthocerotibacter не проливает свет и мало что добавляет (скорее всего ничего) нам для понимания эволюции кислородного фотосинтеза по сравнению с Gloeobacter
Я бы так не сказал. Всё-таки, когда у нас был описан всего один род из группы глеобактерий, мы не могли сказать, не являются ли его относительно примитивные признаки вторичным упрощением? До того момента не было теоретически исключено, что предок всех цианобактерий напоминал ностока: был многоклеточным, имел тилакоиды, циркадные ритмы и сложные фотосистемы. Тогда бы нам пришлось гораздо больше ломать голову, как он приобрёл все эти признаки, если у мелайнабактерий всего этого нет. Но Anthocerotibacter продемонстрировал, что общий предок цианобактерий, скорее всего, напоминал его самого. Вторичное "упрощение" имело место только в плане биосинтеза каротиноидов – а вот тилакоидов не было, и фотосистемы были простые. Кроме того, Anthocerotibacter пока может послужить нам оценкой "снизу", НАСКОЛЬКО они могли быть простыми. Не исключено, что в будущем мы найдём глеобактерию с ещё более простыми фотосистемами, но пока мы можем сказать, что первые ФСI и ФСII после объединения в линии будущих цианобактерий могли быть устроены так же минималистично.
Это уже зацепка, так как мы косвенными методами можем представить себе переходную форму в эволюции фотосинтеза. И теперь есть основания полагать, что именно такой формой общий предок цианобактерий и являлся.
Правда, от себя могу добавить, что эта переходная форма находится всё-таки довольно близко к современным цианобактериям. Сами можете видеть на рис. 10, что у Anthocerotibacter нет в основном периферийных компонентов фотосистем. Весь центр у него цел. В общем, если представить себе путь от мелайнабактерий до "продвинутых" цианобактерий вроде ностока как дорогу из Петербурга в Москву, то Anthocerotibacter – это примерно город Клин. И остаётся интрига, как на этой дороге выглядело хотя бы "Бологое"? Было ли что-то более примитивное?а так статья хорошая
Спасибо большое, Искандар Миртибатович!
использующие энергию солнечного света для закачивания протоков внутрь тилакоидовПротонов жи. :)
У любого из этих мхов есть какой-нибудь бактериальный симбионт из числа многоклеточных цианобактерий, осуществляющий для мха фиксацию атмосферного азотаНу.. не только. Гораздо важнее - источник "дармовой" органики.
а способ повысить напряжениеПотенциал жи. :)
чем у «кроновых» цианобактерий (их представитель делится каждые 8 часов)Какбэ вариации там в довольно широких пределах... :з
Это показывает, насколько неэффективен фотосинтетической аппарат антоцеротибактера. Но при этом скорость выделения кислорода была лишь примерно вдвое ниже, чем у «кроновой» цианобактерии.Взаимоисключающие параграфы, как они есть. :З
А ларчик может просто открываться: это же мутуалистический партнёр; он заточен на генерацию нямки (см. выше); усиленный рост тут ему НЕ НУЖЕН. :)
-
Протонов жи. :)
Спасибо, исправим!Ну.. не только. Гораздо важнее - источник "дармовой" органики.
Зачем мху дармовая органика, когда он сам её себе наделает путём фотосинтеза? А вот фиксировать азот он не умеет. У бобовых есть ризобии, а вот антоцеротовый мох вместо ризобий использует цианобактерий.Потенциал жи. :)
Знаю, я специально сказал бытовым языком, чтобы читателю было понятнее. Потом, с точки зрения физики речь идёт о разности потенциалов по обе стороны мембраны, а разность потенциалов – это напряжение.Какбэ вариации там в довольно широких пределах... :з
Я имел в виду конкретного представителя – Synechocystis sp. PCC 6803, а добавить его имя забыл. Исправим, спасибо!А ларчик может просто открываться: это же мутуалистический партнёр; он заточен на генерацию нямки (см. выше); усиленный рост тут ему НЕ НУЖЕН. :)
Возможно, кстати! Я просил картинки непосредственно у Fai-Wei Li. Пожалуй, напишу-ка я ему и изложу эту Вашу гипотезу... Как Вас представить? Имя, аффилиация?
Последние новости















Рис. 1. Стилизованная микрофотография недавно открытой цианобактерии Anthocerotibacter panamensis. Изображение предоставлено автором обсуждаемой статьи Fay-Wei Li