«Первичность губок» опережает по очкам «первичность гребневиков»

Продолжаются дискуссии вокруг вопроса о том, какая ветвь многоклеточных животных является самой древней: губки или гребневики? На данный момент похоже, что губки «побеждают». Но это, в свою очередь, порождает новые не решенные пока вопросы, важнейший из которых касается происхождения нервной системы.
В поисках предка
Человек вышел из животного царства. В ходе своей истории он постоянно так или иначе имел дело с другими животными: собаки, лошади, киты, акулы, жирафы, страусы, жуки, дождевые черви, скорпионы, морские звезды — все они были частью мира, знакомого людям с глубокой древности. Животными при этом считали всех, кто не растения. Правда, натуралистов издавна озадачивали живые существа, сочетавшие (на первый взгляд) признаки животных и растений и относившиеся невесть куда — например, актинии или губки. Но таких существ было относительно немного — во всяком случае, заметных невооруженным глазом. На протяжении тысячелетий, от Аристотеля до Декарта, общая картина виделась более-менее стабильной и ясной.
Настоящий переворот в представлениях о структуре живой природы произошел после великой научной революции XVII века. В 1677 году в лондонском журнале Philosophical Transactions of the Royal Society (который продолжает выходить по сей день) была опубликована знаменитая работа Антони ван Левенгука «О мельчайших животных». Перед людьми начал приоткрываться огромный, разнообразный, ни на что не похожий мир микроскопических организмов. Сначала этот мир казался исследователям совершенно беспорядочным. Кое-какую ясность внесла провозглашенная в 1839 году Теодором Шванном клеточная теория, согласно которой и животные, и растения состоят из того или иного числа клеток, в целом подобных одна другой. Отсюда логически следовало, что некоторые организмы могут состоять и из одной клетки. И действительно, вышедшее в 1848 году «Руководство по сравнительной анатомии» Карла фон Зибольда и Германа Станниуса открывается утверждением, что все животные делятся на одноклеточных и многоклеточных (автором этого важнейшего обобщения был Зибольд, который написал том, посвященный беспозвоночным). Теперь стало понятно, что микроскопические живые существа — во всяком случае, подавляющее большинство их — это просто-напросто одноклеточные.
Только после работ Левенгука, Шванна и Зибольда стала возможной (а потом и необходимой) постановка проблемы происхождения многоклеточных животных. Надо заметить, что в современной биологии «животными» вообще принято называть только многоклеточных существ (Metazoa), никакие одноклеточные в эту группу не входят, так что словосочетание «многоклеточные животные» — теперь лишь дань традиции. Но в любом случае ясно, что у многоклеточных животных должны были быть одноклеточные предки. Что это за предки? Как произошел переход между уровнями организации — с одноклеточного на многоклеточный? Как выглядели переходные формы? Эти вопросы, впервые осознанные в середине XIX века, занимают биологов до сих пор.
Проще всего ответить на первый вопрос — о том, кем были предки. Ближайшие родственники животных известны: это воротничковые жгутиконосцы (рис. 2). В терминах современной эволюционной биологии многоклеточные животные и воротничковые жгутиконосцы — сестринские группы. Близкое родство воротничковых жгутиконосцев с животными предположили еще в XIX веке, сразу после открытия этих микроскопических организмов, а сейчас оно надежно подтверждается молекулярной генетикой (см. Расшифрован геном хоанофлагеллят — ближайших одноклеточных родичей всех многоклеточных животных, «Элементы», 18.02.2008). Эти группы имеют огромное количество общих генов, включая такие, которые раньше считались уникальным для многоклеточных животных «ноу-хау» (например, у воротничковых жгутиконосцев нашлись гены, кодирующие кадгерины и интегрины — белки, обычно участвующие во взаимодействии животных клеток между собой). Насчет их родства сомнений нет.

Рис. 2. Воротничковые жгутиконосцы. А — одиночный Monosiga, Б — схема токов жидкости, возникающих при пищевом биении жгутика, В — древовидная колония Codonosiga, Г — плавающая колония Sphaeroeca. Масштаб не соблюден! Использованы иллюстрации из книги: Протисты. Ч. 1. Руководство по зоологии. СПб., 2001
Многоклеточные животные, воротничковые жгутиконосцы и еще несколько групп одноклеточных или колониальных организмов образуют большую группу, которая называется Holozoa. В свою очередь, группа Holozoa входит в супергруппу Opisthokonta (кроме голозой, то есть животных и их родственников, к опистоконтам относятся грибы и их родственники). Опистоконты, или заднежгутиковые, — это одна из примерно десятка крупнейших эволюционных ветвей эукариот, организмов со сложной ядросодержащей клеткой (см. Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019). Таково место животных на общем эволюционном древе.
Итак, воротничковые жгутиконосцы — ближайшие современные родственники многоклеточных животных. Значит ли это, что они и есть их предки? Нет, не обязательно. Во-первых, надо помнить, что сколь угодно близкое родство между двумя живыми существами само по себе еще не означает, что одно из них произошло от другого (например, человек не является ни предком, ни потомком своих ближайших современных родственников — шимпанзе и бонобо). Во-вторых, у современных воротничковых жгутиконосцев есть признаки, противоречащие гипотезе, что многоклеточные животные произошли непосредственно от них. Например, сравнительная генетика совершенно однозначно показывает, что у общего предка всей супергруппы Opisthokonta был ген, который называется Brachyury (см. У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). Для многоклеточных животных этот ген очень важен: он регулирует ранние стадии их эмбрионального развития и, в частности, закладку хорды у позвоночных. Есть он и у многих неживотных опистоконт, причем не только среди голозой, но и в другой эволюционной ветви, ведущей к грибам. Более того, ген Brachyury широко распространен не только у колониальных, но и у одноклеточных опистоконт (вероятно, его продукт как-то регулирует взаимное расположение клеток при делении). Но вот у воротничковых жгутиконосцев его нет. Они его утратили. Между тем многоклеточные животные, конечно, могли произойти только от таких предков, у которых ген Brachyury еще был — иначе как бы они его унаследовали? Значит, если и они и произошли от воротничковых, то не от современных.
Еще один повод для размышления дает физиологически обусловленная синхронность делений клеток (в этом случае общее число клеточных ядер в организме какое-то время остается степенью двойки: 2, 4, 8, 16...). Такая синхронность наблюдается при дроблении оплодотворенного яйца большинства многоклеточных животных и — с другой стороны — при образовании колонии у некоторых голозой, не относящихся к воротничковым жгутиконосцам (см. Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). У самих воротничковых ее нет. Все это, вероятно, означает, что жизненный цикл общего предка воротничковых жгутиконосцев и многоклеточных животных был гораздо более сложным, чем жизненный цикл современных воротничковых (который вторично упрощен). Непосредственный предок многоклеточных животных до нашего времени не дожил. Что и неудивительно: ведь с тех пор прошло, вероятно, около 600 миллионов лет.
Гастрея vs фагоцителла
Проблема происхождения многоклеточных животных — одна из самых старых проблем эволюционной биологии. За полтора века на эту тему были высказаны десятки гипотез, в том числе детально разработанных, которые принято повышать в статусе и называть теориями (см. А. В. Иванов, 1968. Происхождение многоклеточных животных). Многие из них сейчас устарели настолько, что представляют интерес только с точки зрения истории науки. Другие, может быть, частично и верны, но по разным причинам непопулярны. Главными соперниками с 1880-х годов и до сих пор выступают две теории (именно их в первую очередь вспомнит любой зоолог). Это теория гастреи Эрнста Геккеля и теория фагоцителлы Ильи Ильича Мечникова.
Теория Геккеля основана на том, что многоклеточным животным свойственны эмбриональные стадии бластулы и гаструлы. В варианте, который принято считать наиболее типичным, бластула представляет собой однослойный шар из жгутиковых клеток. Потом одно полушарие втягивается внутрь, и однослойный шар трансформируется в двуслойный мешок — гаструлу. Возникающая при этом внутренняя полость гаструлы (архентерон) становится кишечной полостью взрослого животного. Геккель предположил, что эти эмбриональные стадии повторяют (рекапитулируют) ход эволюционных событий. Вначале была шарообразная колония жгутиконосцев, похожая на бластулу, но представляющая собой взрослый организм — Геккель назвал ее бластеей. Переднее полушарие бластеи (преимущественно двигательное) так и осталось снаружи, а заднее (преимущественно пищеварительное) втянулось внутрь, образовав камеру со входом. Эта камера и стала первичной кишкой, или архентероном. Получившийся двуслойный организм Геккель назвал гастреей. Предполагается, что гастрея была подобна гаструле, но являлась не промежуточной стадией развития, а конечной (взрослой).
Геккелевская гаструла, устроенная поразительно единообразно, образует своего рода «узел сходства» в индивидуальном развитии великого множества многоклеточных животных, от коралловых полипов до ланцетника включительно. Неудивительно, что Геккель посчитал широкую распространенность этой эмбриональной стадии следом общего для всех животных эволюционного прошлого. Образование гаструлы путем упорядоченного впячивания целого клеточного слоя называется инвагинацией. Типичную инвагинационную гаструлу можно увидеть у представителей самых разных типов животных, как бы сильно ни отличались между собой их взрослые формы (рис. 3).

Рис. 3. Гаструлы разных животных: 1 — известковая губка, 2 — актиния, 3 — ресничный червь, 4 — асцидия, 5 — улитка прудовик, 6 — морская звезда, 7 — ракообразное, 8 — ланцетник. a — первичная кишка (архентерон), o — первичный рот (бластопор), e — внешний слой клеток (эктодерма), i — внутренний слой клеток (энтодерма). У губок нет рта, и соответствие их бластопора таковому других животных спорно. В остальном представленное здесь сравнение выдерживает критику по сей день: у актинии, плоского червя, улитки, ракообразного, морской звезды и двух хордовых (асцидии и ланцетника) гаструла однотипная. По Эрнсту Геккелю из статьи: L. Salvini-Plawen, 1998. Morphologie: Haeckels gastraea-theorie und ihre folgen
Для Геккеля было важно, что инвагинационная гаструла преобладает у таких примитивных животных, как коралловые полипы (к которым относятся актинии). С другой стороны, этот способ развития не является всеобщим. Например, у гидроидных полипов (к которым относится пресноводная гидра) гаструляция чаще всего идет не путем инвагинации, а путем иммиграции: клетки будущей кишки по одной уходят в глубину зародыша. Нет инвагинационной гаструлы и у бескишечных ресничных червей, очень простых и примитивных представителей гигантской ветви двусторонне-симметричных животных (билатерий). Эти факты и привели Мечникова к мысли, что у потомков бластеи клетки первым делом начали уползать внутрь поодиночке, чтобы заниматься внутриклеточным пищеварением. В результате возникла фагоцителла — организм, состоящий из слоя движущих жгутиковых клеток снаружи и рыхлой массы переваривающих клеток (фагоцитов) внутри. Инвагинационная гаструла, по Мечникову, появилась гораздо позже — после того, как масса фагоцитов превратилась в кишечник, имеющий собственную полость (рис. 4).

Рис. 4. Основные теории происхождения многоклеточных животных, совмещенные на одном рисунке. В тексте обсуждаются только теории гастреи и фагоцителлы. Стрелки обозначают гипотетические эволюционные переходы. Кишечнополостные — устаревшее общее название гребневиков и стрекающих. По Адольфу Ремане, с изменениями, из книги: А. В. Иванов, 1968. Происхождение многоклеточных животных
Веским доводом за теорию фагоцителлы и против теории гастреи было убеждение многих зоологов, что гидроидные полипы древнее и примитивнее коралловых. Однако это убеждение никогда не было всеобщим, а появившаяся в 1990-х годах молекулярная систематика животных сразу показала, что оно неверно (D. Bridge et al., 1992. Class-level relationships in the phylum Cnidaria: evidence from mitochondrial genome structure). Именно кораллы — самые древние и примитивные из современных стрекающих, а гидроиды — группа относительно молодая (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). В этом отношении Геккель был прав.
Сколько бы ни спорили между собой сторонники теорий гастреи и фагоцителлы, общего между этими теориями в любом случае гораздо больше, чем различий. Обе теории (по крайней мере в их классических версиях) предполагают, что исходной формой многоклеточных животных была шарообразная жгутиковая бластея, весь жизненный цикл которой проходил в толще морской воды. Такой жизненный цикл называется голопелагическим.
Плавающие шарообразные колонии, похожие на геккелевскую бластею, действительно существуют. Они независимо возникли у далеких друг от друга эукариот, относящихся к разным супергруппам: у золотистых водорослей (см. Synura), у зеленых водорослей (см. Volvox) и у воротничковых жгутиконосцев (см. Sphaeroeca). Последние, как мы знаем, и есть ближайшие родственники животных. Причем сфероэка, бластееподобный представитель воротничковых, была открыта немецким биологом Робертом Лаутерборном больше чем через двадцать лет после того, как Геккель описал гипотетическую бластею. Казалось бы, вот пример удачного предсказания! Но тут есть оговорки. Дело в том, что подавляющее большинство воротничковых жгутиконосцев питается не в плавающем, а в сидячем состоянии: к этому способу жизни они приспособлены гораздо лучше. Для них наиболее типичны прикрепленные колонии, имеющие форму лент, пластинок, звездочек или ветвящихся деревьев. Активно плавающая колония у них редкое исключение.
Кроме того, с теорией гастреи Геккеля плохо согласуются губки — прикрепленные существа, которых многие исследователи считали (и считают до сих пор) самыми примитивными из всех многоклеточных животных. Геккель несколько преувеличил сходство ранних стадий развития губок, с одной стороны, и остальных многоклеточных животных, с другой (всех животных, кроме губок, иногда называют настоящими многоклеточными животными, Eumetazoa). У губок безусловно есть истинная бластула, причем у многих видов свободно плавающая — иными словами, бластула у них может быть личинкой. Но вот наличие у них истинной гаструлы спорно. Встречается обоснованное мнение, что ее просто нет (см., например: L. Salvini-Plawen, 1998. Morphologie: Haeckels gastraea-theorie und ihre folgen). А отсюда следует, что губки, вероятнее всего, отделились от других животных на эволюционной стадии бластеи (L. Salvini-Plawen, 1978. On the origin and evolution of the lower Metazoa). Стадию гастреи они не проходили.
Могли ли настоящие многоклеточные животные, снабженные мускулатурой и нервной системой, произойти от неподвижных губок? И если могли, то как?
Вершки и корешки
Мы знаем, что многоклеточные животные и воротничковые жгутиконосцы относятся к эволюционной ветви Holozoa, входящей, в свою очередь, в состав супергруппы Opisthokonta. Название Opisthokonta — заднежгутиковые — связано с уникальным признаком этой группы: жгутиковые клетки опистоконт, в отличие от всех остальных эукариот, всегда движутся жгутиком назад. Откуда этот признак взялся? Скорее всего, дело в том, что одноклеточные предки заднежгутиковых питались в сидячем состоянии, и жгутик у них бился таким образом, чтобы создавать ток воды, поднимающий пищевые частицы с поверхности дна (В. В. Малахов, 2011. Современные представления о происхождении многоклеточных животных). При переходе к плаванию такой способ работы жгутика как раз и означает, что клетка будет двигаться жгутиком назад. Именно так плавают сперматозоиды животных и жгутиковые споры хитридиевых грибов (напомним, что грибы тоже опистоконты, они гораздо ближе к животным, чем, скажем, растения или инфузории). Древние опистоконты легко прикреплялись к грунту, потому что были в основном обитателями твердых поверхностей — дна или почвы, а не открытого водного пространства, как многие эукариоты из других супергрупп (например, одноклеточные зеленые водоросли, см. Хламидомонада).
Со жгутиком опистоконт связана еще одна проблема. Дело в том, что любой эукариотный жгутик закреплен в теле клетки с помощью так называемого корешкового аппарата, который устроен у разных эукариот совершенно по-разному (см. С. А. Карпов, 2001. Строение клетки протистов). Есть два главных типа корешковых аппаратов: ленты микротрубочек и исчерченные корешки, состоящие из микрофиламентов (микрофиламенты тоньше микротрубочек, но зато их пучки, в отличие от пучков микротрубочек, могут сокращаться). Электронно-микроскопические исследования, в ходе которых были открыты эти структуры, сразу показали, что тип корешкового аппарата часто служит важным систематическим признаком (см., например: С. А. Карпов, 1990. Система протистов).
Проблема же заключается вот в чем. У воротничковых жгутиконосцев корешковый аппарат состоит из лент микротрубочек, радиально расходящихся от основания единственного жгутика (поскольку клетка у воротничковых радиально-симметричная). У жгутиковых клеток губок — хоаноцитов, которые во всех отношениях очень похожи на воротничковых жгутиконосцев, корешковый аппарат тоже состоит из лент микротрубочек. А вот у всех остальных многоклеточных животных, от кораллов и плоских червей до человека, корешковые аппараты жгутиков и ресничек — это исчерченные корешки.
Кроме того, и у воротничковых жгутиконосцев, и у хоаноцитов губок жгутик несет многочисленные тонкие придатки — мастигонемы (особенность, для нефотосинтезирующих эукариот очень редкая). У настоящих многоклеточных животных мастигонем не бывает никогда.
Тонкие особенности устройства живых клеток, для изучения которых нужен электронный микроскоп, в биологии принято называть ультраструктурой. Итак, между воротничковыми жгутиконосцами и хоаноцитами губок есть ультраструктурное сходство, которое свидетельствует о родстве воротничковых жгутиконосцев и губок друг с другом, но отличает и тех и других от настоящих многоклеточных животных. В 1980-х годах это послужило основанием для гипотезы, что от воротничковых жгутиконосцев произошли только губки, в то время как происхождение всех остальных многоклеточных животных — совершенно иное (Л. Н. Серавин, 1986. Природа и происхождение губок). Губок с воротничковыми выделяли в особое царство Parazoa (см. О. Г. Кусакин, А. Л. Дроздов, 1994. Филема органического мира. Ч. 1). От кого в таком случае произошли настоящие многоклеточные животные, оставалось загадкой. На роль их вероятных предков предлагались то хитридиевые грибы, то жгутиковые зеленые водоросли. На самом деле, как мы сейчас понимаем, хитридиевые грибы относятся к другой ветви опистоконт, а зеленые водоросли — вообще к другой супергруппе эукариот.
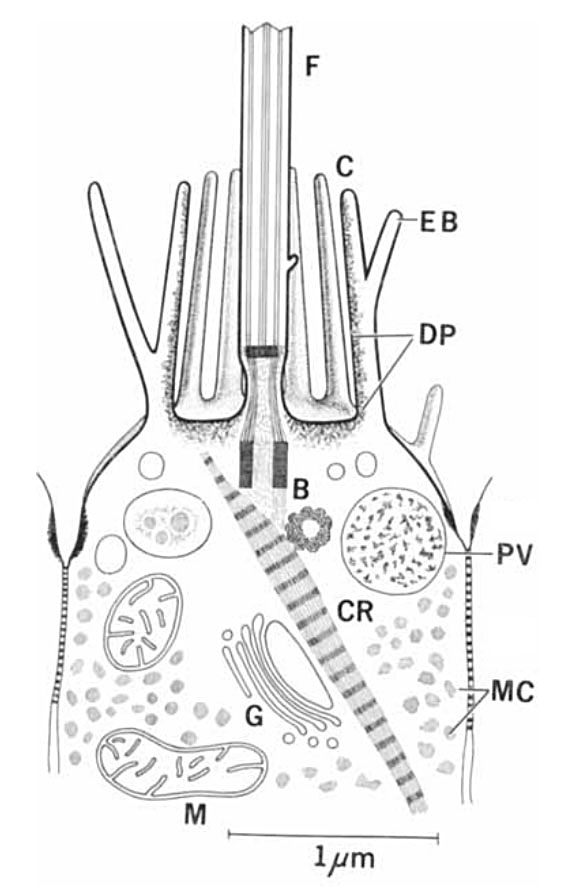
Рис. 5. Жгутиковая клетка личинки морской звезды. Тут прекрасно видно микроворсинки воротничка и исчерченный корешок. F — жгутик, C — воротничок, EB — внешняя ветвь микроворсинки, DP — электронноплотная зона, B — центриоли, CR — исчерченный корешок, PV — пищеварительная вакуоль, MC — мелкие включения в цитоплазме, G — аппарат Гольджи, M — митохондрия. Иллюстрация из статьи: A. Norrevang, K. G. Wingstrand, 1970. On the occurrence and structure of choanocyte-like cells in some echinoderms
У настоящих многоклеточных животных тоже встречаются воротничковые клетки, напоминающие о воротничковых жгутиконосцах. Воротничок, которому воротничковые жгутиконосцы обязаны своим названием — это прозрачная воронка, окружающая жгутик и состоящая из множества отдельных микроворсинок, длинных тонких выростов с микрофиламентами внутри (см. рис. 1, А). У хоаноцитов губок он устроен очень похоже. Но эта структура не уникальна. Жгутиковые клетки с настоящим воротничком из микроворсинок описаны, например, у личинок некоторых полипов, причем как коралловых, так и гидроидных (K. M. Lyons, 1973. Evolutionary implications of collar cell ectoderm in a coral planula). Специальные исследования показали, что воротничковые клетки широко распространены у иглокожих (не только в покровах, но и в выстилке воднососудистой системы; см. рис. 5 и статью: A. Norrevang, K. G. Wingstrand, 1970. On the occurrence and structure of choanocyte-like cells in some echinoderms). Типичные воротничковые клетки найдены и в покровах полухордовых (A. Norrevang, 1964. Choanocytes in the skin of Harrimania kupfferi (Enteropneusta)), и в предротовой воронке ланцетника (T. Stach, 1996. On the preoral pit of the larval amphioxus (Branchiostoma lanceolatum)).
Более того, преемственность воротничковых клеток дотянулась и до позвоночных. Именно от воротничковых клеток, судя по всему, произошли сенсорные клетки внутреннего уха, каждая из которых несет жгутик и группу микроворсинок (правда, жгутик там расположен не посреди микроворсинок, а эксцентрично). Недавно было показано, что существует большая группа общих генов, которые активны в сенсорных клетках внутреннего уха позвоночных и в хоаноцитах губок (J. F. Pena et al., 2016. Conserved expression of vertebrate microvillar gene homologs in choanocytes of freshwater sponges). Функции белков, кодируемых этими генами, связаны или со сборкой микрофиламентов, или с контактами микроворсинок между собой. Пример такого белка — виллин, который назван в честь микроворсинок (по-латыни microvilli) и регулирует сборку пучков микрофиламентов, дающих микроворсинкам опору. Виллин есть у настоящих многоклеточных животных, у губок, у воротничковых жгутиконосцев и у амебоидных Holozoa (см. Capsaspora), но не найден ни у каких других эукариот.
Однако во всех жгутиковых клетках настоящих многоклеточных животных корешковый аппарат жгутика представлен исчерченным корешком, а не лентами микротрубочек. В этом отношении жгутиковые клетки настоящих многоклеточных животных (неважно, воротничковые или нет) резко отличаются и от воротничковых жгутиконосцев, и от хоаноцитов губок. Не разобравшись в причинах этих отличий, делать выводы о происхождении всех многоклеточных животных было нельзя.
Неотенические личинки губок?
К началу XXI века тщательные электронно-микроскопические исследования прояснили ситуацию. Оказалось, что у бластулообразных личинок губок жгутиковые клетки как раз могут обладать типичными исчерченными корешками, которые исчезают и сменяются лентами микротрубочек только при метаморфозе (см. M. Maldonado, 2004. Choanoflagellates, choanocytes, and animal multicellularity). Это установлено как минимум для нескольких видов губок, относящихся к разным классам. У жгутиков личинок губок есть и другие черты сходства со жгутиками настоящих многоклеточных животных: кое-какие детали расположения микротрубочек внутри жгутика, отсутствие воротничка и мастигонем (рис. 6).

Рис. 6. Жгутиковые клетки воротничкового жгутиконосца (слева), типичной взрослой губки (в центре) и личинки известковой губки (справа). v — мастигонемы, mi — микроворсинки, m — микротрубочки, bb — центриоль, bf — базальная подошва, ac — добавочная центриоль, r — дополнительный корешок, mt — митохондрии, g — аппарат Гольджи, sr — исчерченный корешок. Иллюстрации из статьи: M. Maldonado, 2004. Choanoflagellates, choanocytes, and animal multicellularity
Кроме того, жгутиковые клетки личинок губок нередко бывают подостланы базальной мембраной, которая характерна для эпителиев настоящих многоклеточных животных, а среди взрослых губок встречается только у представителей небольшого класса гомосклерид. У личинок тех же гомосклеридных губок описаны и десмосомы — межклеточные контакты весьма продвинутого типа, которые обычно обеспечивают в эпителиях настоящих многоклеточных животных механическое сцепление клеток друг с другом.
Строго говоря, эти факты — не такая уж и новость. Часть их была известна уже в шестидесятых-семидесятых годах XX века. Австрийский зоолог Луитфрид Сальвини-Плавен считал, что смена корешкового аппарата у губок связана с характером нагрузки: жгутикам плавающей личинки, в отличие от жгутиков неподвижной взрослой губки, надо обеспечивать поступательное движение животного в толще воды, потому они и снабжены корешками из микрофиламентов, способными активно сокращаться (см. L. Salvini-Plawen, 1978. On the origin and evolution of the lower Metazoa).
Получается, что никакой пропасти между губками и настоящими многоклеточными животными на самом деле нет.
Теперь подведем некоторый итог. У жгутиковых клеток настоящих многоклеточных животных для «заякоривания» жгутика в теле клетки служит исчерченный корешок, воротничка из микроворсинок вокруг жгутика чаще все же нет (несмотря на вышеперечисленные примеры, обязательным приложением к жгутику он тут не является), мастигонем не бывает, а поверхностный слой клеток почти всегда представляет собой полноценный эпителий с десмосомами и базальной мембраной. И мы видим, что по сумме структурных признаков настоящие многоклеточные животные гораздо более похожи на личинок губок, чем на взрослых губок.
Вряд ли это случайность. В роли предка настоящих многоклеточных животных трудно представить взрослую губку, но достаточно легко — бластулообразную личинку губки. Напрашивается идея, что именно личинка губки и была этим предком. Конечно, речь идет о личинке, отказавшейся от метаморфоза и научившейся самостоятельно размножаться — явление, хорошо знакомое зоологам еще с XIX века и называемое неотенией (см., например: B. A. Pierce, H. M. Smith, 1979. Neoteny or paedogenesis?). Датский зоолог Клаус Нильсен так и назвал одну свою обзорную статью: «Шесть главных шагов в эволюции животных: являемся ли мы производными личинок губок?» (C. Nielsen, 2008. Six major steps in animal evolution: are we derived sponge larvae?) — и ответ он дает положительный: да, являемся.
Неотеническая личинка губки и есть геккелевская бластея. Во всяком случае, это возможно. Что касается дальнейшей эволюции Eumetazoa, то она вполне могла идти примерно так, как описывает геккелевская теория гастреи — но только с дополнением, касающимся происхождения исходной формы, бластеи. Если эта гипотеза верна, значит, мы все — прямые потомки неотенических личинок губок (рис. 7).

Рис. 7. Элементы происхождения многоклеточных животных по Нильсену. А — гипотетический предок губок, Б — типичная губка, В — гомосклеридная губка, личинка которой наиболее похожа на геккелевскую бластею, Г — гипотетическая губка с гаструлоподобной личинкой, уже освоившей самостоятельное размножение (показаны половые клетки). Когда неотеническая личинка окончательно прекратит оседать на дно и проходить метаморфоз, губка перестанет быть губкой (это могло произойти и на стадии бластулы, а не гаструлы). Красная линия обозначает базальную мембрану. Иллюстрация из статьи: C. Nielsen, 2008. Six major steps in animal evolution: are we derived sponge larvae?
К сожалению, надежно определимые элементы минерального скелета губок (они называются спикулами) появляются в палеонтологической летописи только в кембрийском периоде, то есть примерно тогда же, когда и все основные группы настоящих многоклеточных животных (см. А. Ю. Журавлев, 2014. Ранняя эволюция Metazoa — взгляд палеонтолога). Поэтому происхождение Eumetazoa от кого-то из современных классов губок маловероятно. Доводы в пользу того, что губки появились намного раньше, остаются большей частью косвенными (например, химические биомаркеры, которые, по некоторым предположениям, могут принадлежать и водорослям). Правда, надо учитывать, что у некоторых губок твердого скелета вообще нет (см. Oscarellidae). А если древнейшие губки были бесскелетными, то шансов сохраниться они имели немного.
В 2015 году международная группа палеонтологов описала открытые в известном китайском местонахождения Доушаньтуо остатки древнего организма, который получил название Eocyathispongia qiania (Z. Yin et al., 2015. Sponge grade body fossil with cellular resolution dating 60 Myr before the Cambrian). Это было сидячее существо очень небольшого размера — около 2 миллиметров, и оно сохранилось достаточно хорошо, чтобы под электронным микроскопом можно было рассмотреть минерализованные остатки отдельных клеток. Общая форма тела, множество дырочек (видимо, служивших для входа воды), внутренние камеры, трубчатое устье, характерные очертания покровных клеток-пинакоцитов — все эти детали позволили палеонтологам предположить, что эоциатиспонгия была самой настоящей губкой. Правда, маленькой и бесскелетной. А жила она 600 миллионов лет назад, почти за 60 миллионов лет до начала кембрийского периода и до знаменитого кембрийского взрыва (см. С. А. Ястребов, 2016. Кембрийский взрыв). Первые бесспорные остатки настоящих многоклеточных животных появляются в летописи не более чем за 13–15 миллионов лет до начала кембрия (см. Кимберелла). Так что если эоциатиспонгия интерпретирована верно — значит, губки все-таки опередили настоящих многоклеточных животных примерно на 45 миллионов лет.
Проблема гребневиков
Ситуация едва успела проясниться, как появилась новая гипотеза, основательно спутавшая карты классическим зоологам: самая древняя эволюционная ветвь многоклеточных животных — не губки, а гребневики. Эта проблема обсуждалась на страницах «Элементов» уже несколько раз, поэтому сейчас будет достаточно напомнить основные моменты. Гребневики — это полупрозрачные морские хищники, очень отдаленно похожие на медуз; подавляющее большинство их проводит всю свою жизнь, активно плавая в толще воды, но несколько представителей перешли к ползанию по дну (скорее всего, эволюционно недавно). Гипотеза «первичности гребневиков» впервые была высказана молекулярными биологами в 2013 году (см. Геном гребневиков говорит в пользу двукратного возникновения нервной системы у животных, «Элементы», 19.12.2013). Отсюда сразу же вытекала серьезная проблема, связанная с происхождением нервной системы. Дело в том, что у губок нервной системы нет и, судя по всему, никогда не было. Она есть, с одной стороны, у гребневиков, а с другой — у стрекающих и у двусторонне-симметричных, или билатерий (эти две группы образуют единую ветвь). Если верна гипотеза «первичности гребневиков», значит, ветви гребневиков и стрекающих с билатериями находятся на эволюционном древе слишком далеко друг от друга, чтобы их общий предок мог иметь нервную систему. Она должна была появиться независимо.
Вскоре гипотеза «первичности гребневиков» получила новые подтверждения, причем как со стороны исследования родственных связей, так и со стороны генов, функции которых связаны с работой нервной системы и мышц (см. Гипотеза о двукратном появлении нервной системы получила новые подтверждения, «Элементы», 26.05.2014). По сравнению со стрекающими и билатериями гребневики имеют совершенно иной набор нейромедиаторов, рецепторов, синаптических белков и некоторых других молекул, обладающих сигнальными функциями. Гены семейства Hox, играющие огромную роль в дифференцировке отделов тела (включая нервную систему) и у билатерий, и у стрекающих, у гребневиков отсутствуют вообще. Архитектура нервной системы — причем на всех возможных уровнях — у них альтернативная. В этом отношении гребневики еще больше похожи на инопланетян, чем иглокожие, облик которых дал инопланетным разумным существам Питер Уоттс в знаменитом романе «Ложная слепота».
Все это выглядит достаточно убедительно. Однако еще одна серьезная проблема заключается вот в чем: после принятия гипотезы «первичности гребневиков» становится очень трудно представить, как мог бы выглядеть общий предок всех животных (если, конечно, не считать его одноклеточным). Стройный сценарий происхождения всех настоящих многоклеточных животных от губок разрушается — ведь губки теперь оказываются более молодой группой, чем подвижные, хищные, имеющие нервную систему гребневики, — а взамен ничего толком не предлагается.
В 2015 году группа известных зоологов, в которую входил тот же Клаус Нильсен, опубликовала большую статью с анализом этой проблемы (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Эти авторы подчеркнули, что вопрос о положении гребневиков на эволюционном древе и вопрос о происхождении нервной системы суть в общем случае разные вопросы, которые лучше бы разделять, чтобы не получилась путаница. Что касается положения гребневиков на древе, то тут есть многочисленные техническое сложности, связанные с анализом генных последовательностей этой группы животных — а она в любом случае очень древняя, и в ее истории были бурные эпизоды вроде почти полного вымирания в момент мел-палеогенового кризиса, после которого, однако, разнообразие гребневиков быстро восстановилось; неудивительно, что запутаннные генетические следы таких эволюционных событий сбивают биоинформатиков с толку. Самостоятельно решать вопрос о родственных связях гребневиков Нильсен и его коллеги не пытаются. Происхождение нервной системы — другое дело. Нейробиологические данные (сами по себе не связанные с темой родства) убедительно показывают, что нервная система гребневиков устроена крайне своеобразно, если сопоставлять ее с нервной системой стрекающих и билатерий. Вопрос в том, с какого уровня они все стартовали.
Между тем молекулярные филогенетики, занимающиеся именно проблемой родственных связей между разными группами животных, продолжали свои исследования. В том же 2015 году появилась скрупулезно выполненная работа, в которой сравнивалась надежность гипотез «первичности гребневиков» и «первичности губок» с точки зрения топологии эволюционного древа (Сравнительная геномика вынуждает пересмотреть место гребневиков на эволюционном древе животных, «Элементы», 18.12.2015). Было показано, что решение этой задачи неустойчиво: оно зависит от выбранной вычислительной методики и от того, какие группы организмов, помимо животных, включены в анализ (например, в одном случае для сравнения используются данные по грибам, в другом по воротничковым жгутиконосцам — и выводы получаются разными). В общем, результат здесь в любом случае на грани достоверности. Если же аккуратно сопоставить все возможные методики и сочетания выбранных групп, то чаша весов, похоже, склоняется всё-таки в сторону «первичности губок».
Мнение Нильсена
Несколько недель назад вышел очередной обзор всего этого комплекса проблем, на сей раз подписанный Клаусом Нильсеном как единственным автором. Знаменитый датский зоолог суммирует накопившиеся факты и предлагает эволюционный сценарий, который кажется ему самым вероятным.
Прежде всего, за последние годы вышло еще несколько молекулярно-филогенетических работ, где на основе постоянно растущих наборов данных вновь и вновь отстаивается гипотеза «первичности губок» (см., например, рис. 8 и статью: P. Simion et al., 2017. A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals). Гипотеза «первичности гребневиков» там объявляется вычислительным артефактом, научной ошибкой, и делается вывод, что гребневики — только вторая по порядку отхождения ветвь эволюционного древа животных (после губок).

Рис. 8. Эволюционное древо многоклеточных животных, соответствующее гипотезе «первичности губок». Иллюстрация из статьи: P. Simion et al., 2017. A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals
Конечно, это не означает немедленного прекращения дискуссий по данному вопросу. Не исключено, что окончательное «укоренение» ветвей губок и гребневиков вообще находится за пределом разрешающей способности молекулярной филогенетики как метода (W. Pett et al., 2019. The role of homology and orthology in the phylogenomic analysis of metazoan gene content). Такое мнение тоже надо принять во внимание. Похожую проблему можно наблюдать, например, в лингвистике (см. Лингвистическая эволюция сходна с биологической, «Элементы», 18.10.2007). Языковеды прекрасно справляются с установлением родственных связей языков внутри семей (скажем, индоевропейских языков друг с другом), а вот поиск родственных связей между разными языковыми семьями — так называемая макрокомпаративистика — остается хотя и интересной, но крайне дискуссионной темой (см. Г. С. Старостин, 2016. К истокам языкового разнообразия). Наряду с энтузиастами макрокомпаративистики есть исследователи, считающие, что сколько-нибудь надежные результаты в этой области невозможны принципиально, потому что для языков, разошедшихся более 8–10 тысяч лет назад, следы родства становятся неотличимы от случайных совпадений. Анализ генетических текстов, конечно, позволяет проникать на несравненно большую глубину (вплоть до миллиардов лет), но его разрешающая способность тоже не бесконечна: рано или поздно следы родства забиваются «шумом», и с этим ничего сделать уже нельзя.
В любом случае гипотезу «первичности гребневиков» сейчас никак нельзя признать победившей, и к гипотезе «первичности губок» приходится относиться серьезно. Как рабочая версия она до сих пор пригодна.
Что же теперь можно сказать о происхождении нервной системы? Характерно, что авторы вышеупомянутых статей о «первичности губок» — молекулярные филогенетики, занимающиеся проблемами родства — решать этот вопрос не берутся (во всяком случае, пока), а оставляют открытыми несколько возможностей.
Всего есть пять эволюционных ветвей многоклеточных животных: губки, гребневики, пластинчатые, стрекающие и билатерии. Самая маленькая ветвь — пластинчатые: всего два известных представителя, трихоплакс и описанная в 2018 году хойлунгия, отличающаяся от трихоплакса исключительно генетически. Самая огромная ветвь — билатерии: около полутора миллионов описанных видов (две трети из которых — насекомые). У гребневиков, стрекающих и билатерий нервная система есть, у губок и у пластинчатых ее нет. Это сухие факты, а вот дальше начинаются интерпретации.
Существует гипотеза, что нервная система когда-то была у всех животных, но губки и пластинчатые ее по каким-то причинам утратили (см., например: J. F. Ryan, M. Chiodin, 2015. Where is my mind? How sponges and placozoans may have lost neural cell types). Нильсен прямо называет эту гипотезу плодом воображения. Нервная система — штука весьма универсальная («адаптация общего значения» в терминах Ивана Ивановича Шмальгаузена), она может пригодиться почти что при любом образе жизни; несколько случаев ее потери у животных действительно известно, но все они связаны с переходом к паразитизму, а ни для губок, ни для пластинчатых нет оснований такое предполагать. Сильным аргументом в пользу большой древности нервной системы часто считают наличие у губок и у пластинчатых ряда генов, которые ассоциированы с нервной системой у ее обладателей (продукты этих генов обеспечивают срабатывание ионных каналов, выброс из клетки нейромедиатора и тому подобное). Однако сами по себе эти генетические факты мало что доказывают. Ген всего лишь определяет структуру белка, а физиологические эффекты этого белка в ходе эволюции могут меняться, и иногда очень сильно.
Например, есть такое понятие, как протосинаптические белки (P. Burkhardt, S. G. Sprecher, 2017. Evolutionary origin of synapses and neurons — bridging the gap). Синапс — это специфический контакт между нервными клетками, через который передается сигнал (химический или электрический). Клетки, образующие синапс, оснащены в зоне контакта сложными молекулярными конструкциями, в создании которых участвуют десятки белков, кодируемых соответствующими генами. Многие из этих синаптических белков есть не только у животных с нервной системой, но и у губок. На первый взгляд это хорошее основание для гипотезы, что у губок раньше были нервные клетки. Но значительная доля синаптических белков обнаружена (благодаря исследованиям геномов и транскриптомов) не только у губок, но и у воротничковых жгутиконосцев. И у еще более далеких от животных амебоидных Holozoa они тоже встречаются. Нельзя же предполагать, что вообще у всех Holozoa когда-то была нервная система (то есть предположить-то можно, но это будет очевидный пример сведения к абсурду). Вот и пришлось переименовать синаптические белки в протосинаптические: ясно ведь, что у одноклеточных организмов их функции — иные, чем у обладателей нервных систем. Хотя сами белки — те же самые, многие из них сохраняются на всем протяжении эволюционного древа от одноклеточных амеб до позвоночных.
Что могут делать протосинаптические белки у одноклеточных организмов? Например, помогать концентрировать ионные каналы в определенных областях клеточной мембраны (белки GKAP), регулировать сборку микрофиламентов в клеточных выростах (белки Shank) или обеспечивать слияние движущихся внутри клетки мембранных пузырьков (белки SNARE). А уже после возникновения многоклеточности все эти функции белков пригодились для создания архитектуры синапсов. Следует добавить, что для многих протосинаптических белков (которые точно есть и у животных с нервной системой, и у одноклеточных Holozoa) исходные функции просто еще неизвестны — возможно, эти детали прояснятся в ближайшее время.
Таким образом, наличие нервной системы у предков губок и пластинчатых ничем серьезно не подтверждается. Что касается гребневиков, то они отличаются от стрекающих с билатериями и по набору нейромедиаторных систем, и по молекулярной архитектуре синапсов (она «собрана» на основе отличающегося набора белков), поэтому гипотеза о самостоятельном возникновении их нервной системы выглядит хорошо обоснованной независимо от положения гребневиков на филогенетическом древе. Смена гипотезы «первичности гребневиков» на гипотезу «первичности губок» в этом отношении ничего принципиально не меняет. Разбираться нужно именно с молекулярными компонентами нервных клеток, во всех деталях, чтобы в качестве промежуточного итога можно было реконструировать состояние, свойственное общему предку гребневиков, стрекающих и билатерий, и посмотреть — насколько сильно оно отличается от состояния губок. Тогда и можно будет попытаться окончательно выяснить, сколько раз происходил эволюционный прорыв на уровень «настоящих многоклеточных животных» (с нервной системой). На данный момент гипотеза, что он произошел дважды, остается достаточно вероятной.
Общим предком гребневиков, пластинчатых, стрекающих и билатерий (то есть всех животных, кроме губок) Нильсен считает уже знакомую нам геккелевскую гастрею — голопелагический организм, потомки которого довольно быстро в эволюционном масштабе времени (но независимо в двух разных линиях) приобрели нервную систему и мышцы. Гребневики так до сих пор и остались голопелагическими: у подавляющего большинства из них весь жизненный цикл проходит в плавающем состоянии. А вот предки стрекающих и билатерий в некоторый момент опустились на дно, сохранив, однако, плавающую личинку. Свойственная многим стрекающим жизненная форма медузы, а также многочисленные активно плавающие билатерии вплоть до морских стрелок, рыб и китов — это уже результаты вторичного возвращения в толщу воды.
Совершенно особый статус имеет ветвь пластинчатых. Ее положение на эволюционном древе до сих пор окончательно не установлено: на большинстве полученных деревьев пластинчатые выглядят сестринской группой по отношению к ветви, объединяющий стрекающих и билатерий, но есть и такие реконструкции, где пластинчатые находятся ближе к стрекающим, чем к билатериям (C. E. Laumer et al., 2018. Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias). Насколько велико значение этих «разночтений» — пока неясно. У пластинчатых нет ни рта, ни мышц, ни нервных клеток, но есть уникальная система межклеточной сигнализации, основанная на использовании разнообразных пептидов, то есть коротких цепочек аминокислот (F. Varoqueaux et al., 2018. High cell diversity and complex peptidergic signaling underlie placozoan behavior). Не исключено, что родственниками пластинчатых — только гораздо более крупными — были некоторые представители загадочной вендской фауны, родство которых с многоклеточными животными сейчас считается доказанным (см. Подтверждена принадлежность дикинсонии к животному царству, «Элементы», 24.09.2018). Нильсен считает пластинчатых потомками гастреи, которая легла на дно и распласталась по нему. При этом слой клеток, выстилавший первичный кишечник (архентерон), обратился в подошву, на которой животное теперь ползает. Тут можно только добавить, что очень похожие взгляды на природу пластинчатых высказывались и раньше (см. В. В. Алешин, Н. Б. Петров, 2001. Регресс в эволюции многоклеточных животных).
В последней части своей статьи Нильсен касается проблемы соотношения результатов молекулярной филогенетики (основанных исключительно на чтении нуклеотидных текстов) с данными обычной морфологии, изучающей структуру организмов. Из двух обсуждаемых конкурирующих гипотез одна, а именно гипотеза «первичности гребневиков», не имеет морфологической поддержки: у гипотетической эволюционной ветви, включающей всех многоклеточных животных, кроме гребневиков, нет ни одного общего уникального (синапоморфного) структурного признака. Напротив, у гипотезы «первичности губок» морфологическая поддержка есть: тут можно без труда назвать уникальные признаки для каждой из ветвей, возникающих на эволюционном древе. Нильсен — ученый, глубоко преданный классической морфологии, и его явно радует тот факт, что гипотеза «первичности губок» сейчас побеждает.
Источник: Claus Nielsen. Early animal evolution: a morphologist’s view // Royal Society open science. 2019. V. 6. № 7. 190638.
Сергей Ястребов
-
-
-
Приятно читать, не перегружен и в то же время охвачено все самое важное. Честно сказать что идея с гребневиками мне никогда не нравилась чисто из соображений логичности, если их инновации оказались столь успешными что не дали дальнейшего развития, однозначно они попали в эволюционный тупик, в то же время несовершенные губки имели огромный простор для эволюции. Эдакий общий закон.
-
Спасибо.
А зачем им развиваться дальше, если им и так хорошо?
Идея с гребневиками плоха прежде всего тем, что у нее нет морфологической поддержки. Конечно, если бы молекулярные филогенетики однозначно установили первичность гребневиков - пришлось бы это принять и думать: как теперь выглядит сценарий происхождения Metazoa в целом. Но, похоже, не придется...-
Отличная статья!
Спасибо большое!А зачем им развиваться дальше, если им и так хорошо?
А им совершенно не обязательно всегда было хорошо.
М.б., когда-то давно им почему-то захотелось бОльшего, или катаклизьма случилась, и вдруг сменилась среда обитания - в итоге какой-то толчок дал дальнейшее развитие.
При этом в среде, не подвергшейся изменениям, остались и формы, близкие к исходным.
Но, в рамках послеобеденного бреда и чисто с позиции диванного воЕна, вариант с первичностью ктенофор выглядит куда как интереснее :))) - губки логичнее, и потому, как минимум, скучнее :)))
-
-
Простите а зачем выводить предка животных из неотенической личинки губок, может плавающая форма была первичной, а губки это приспособления к сидячей жизни фильтратора.
Представте что ланцетники вымерли мы б выводили хордовых из личинок асцидий-
Нет, не выводили бы. Это не выдерживает критики. Личинка асцидии, лишенная метамерии, с мозаичным онтогенезом, на роль предка хордовых не подходит.
Сам вопрос, конечно, правильный, но (1) я не случайно подчеркиваю, что среди воротничковых преобладают сидячие формы, (2) хоаноциты губки настолько похожи на воротничковых жгутиконосцев, что тут легко предположить самое что ни на есть прямое родство.-
1. среди СОВРЕМЕННЫХ воротничковых а они не могут быть предками утратили этот важный ген ....
2 Хоаноциты губок похожи но волосковые клетки так же похожи +
так навскидку клетки эпителия тонкого кишечника - воротничковые без жгутика, сперматозоид - воротничковый без ворсинок
Просто логичнее первичность плавающей формы, насколько я понимаю личинка губки не питаются чисто для расселения
ну по мне цепочка колония воротничковых - плавающая бластея
( общий предок губок одна линия и остальных животных вторая)
схема Нильсена это избыточность
Давайте еще раз допустим у нас нет ни ланцетников ни пикай и тулимонстров ни круглоротых рыбы и оболочники
Так же писали личинка СОВРЕМЕННЫХ не может быть предком позвоночных потомушто мозаичный онтогенез а вот каких то древних асцидий .....
простите за несколько корявое изложение мыслей-
Не вижу, почему логичнее первичность плавающей формы, если почти все хоанофлагелляты - сидячие.
И насчет волосковых клеток не согласен. Хоаноциты губок похожи на хоанофлагеллят на ультраструктурном уровне, совпадение в мельчайших деталях (я об этом пишу), с волосковыми и прочими клетками нет такого.-
Возможно, логичнее - потому что формирование плавающих жгутиконосных колоний это вполне себе классический параллельный эволюционный тренд, вплоть до тех же воротничковых, которые практически инвагинируют для плавания. Экологические в комбинации с функционально-морфологическими соображениями подсказывают, что усложнение организации в сторону образования организма скорее пойдут из активной плавающей формы, нежели из сидячей колонии.
Тут, правда, вопрос, что считать "первичным" - сидячую колонию, из которой образовалась плавающая, или плавающую, из которой образовалось все остальное ;)-
Как то так, мне тоже трудно представить чтобы специализированная сидячая форма дала начало чему то принципиально новому
-
-
Ну смотрите, по Нильсену получается что допустим сидячая колония воротничковых стала губкой, на основании каких эволюционных механизмов они получили планктонную личинку если у них НЕ было планктонной фазы, + закономерность макроэволюции
Правило происхождения от малоспециализированных предков: новые крупные группы берут начало от сравнительно неспециализированных представителей предковых групп. ( эволюционный тренд)
Ну и у нас есть навскидку двустворки усоногие и оболочники и нигде сидячие формы ничего нового не дали, я правда не встречал ( может просто не интересовался выкладками что первично полип или медуза)-
1. Морским сидячим организмам обязательно нужна хоть какая-то расселительная стадия - это железная закономерность. Вот если бы такая стадия не возникла, было бы и вправду удивительно.
2. А как Вы отличаете сильно специализированные организмы от малоспециализированных? Строго говоря, специализированным является _любой_ организм, который в ходе эволюции выжил. Только направления специализации различаются.
3. У стрекающих первичен полип - это однозначно.-
Мне кажется, тут происходит типичное усложнение по диалектике (прости ее, господи) - первичен организм - метаболический мешок (или голый репликатор, если совсем вглубь улезть), потом он научился шарахаться (а _прилип_ еще раньше!!), потом перешел к многоклеточности, оставив шарахающуюся стадию на этапе рассселения (=усатые гаметы почти всех на свете, кроме голых мешков, ветром (течением) гонимых - тоже есть что проверить), потом возникли подвижные колонии, у которых вопрос расселения опять снят (что там у вольвоксо-подобной организации с подвижностью гамет? - она уже не безусловно необходима, может кто успел утратить?) - и, наконец, "наш" организм.
-
Я бы поставил под сомнение уже первую тезу этой диалектики. Почему Вы исходите из того, что первый организм - это голый мешок, который где-то там плавал? Например, по Мартину-Расселу-Кунину первые реплицирующиеся системы были замурованы в микрополостях пористых грунтов, и только много позже, обзаведясь мембраной, совершили клеточный побег в открытую воду.
-
Ключевое слово - активная подвижность.
Мешок - он и в африке мешок, хоть замурован, хоть болтается в растворе без руля и ветрил - ему слова никто не давал ;)
Можно было бы предположить, что из замурованного состояния первый побег вверх был - двигать хоть что-то, хоть слизь выталкивать a'la те диатомеи, черт бы их нюхал - забыл!, почему-то диктиостелла всплывает. Но это - боковая мысль, чисто для полноты.-
Толку от постоянного свободного плавания в исходно пустой среде, никаким "бульоном" на деле никогда не бывшей?
В природе чуть менее чем все прокариотные клетки находится в составе биоплёнок, а не в планктонной форме. Жизнь вообще тяготеет не к толще, а к границам раздела сред и сегодня: чуть менее чем вся океанская биомасса находится в прибрежной зоне.
-
-
-
-
-
-
-
-
-
Тут, однако, такое дело - есть у нас "шовинизм подвижности" - мы, билатерии, привыкли воспринимать подвижные формы как наиболее естественные, оглядываясь на всяких там растений и грибов через плечо и свысока. Меж тем, даже из перешедших к какой-никакой многоклеточности - 5 из 6 (- мои любимые, или и вовсе "9 из 10" по-простому) живут в основном сидючи, шастают только отдельные беспокойные представители, да и в одноклеточном состоянии, если без шор посмотреть - _активная_ подвижность, да еще с сенсорами и внутренней координацией - удел безусловно _непростых_ форм.
То есть с точки зрения снаружи от жизни - специализированны как раз активно подвижные формы, у них для этого целая организация должна быть, почти по Северцеву, ежели бы он об этом подумал ;) -
-
-
-
-
-
-
-


Также у губок нет мастигонем, у них бывают vanes - продольные складки на жгутиках, но по структуре они совершенно другие.
Спасибо за статью!
-
Спасибо за ссылку!
1. По поводу мастигонем у меня встречный вопрос: а у хоанофлагеллят они есть? В работах начала 2000-х годов тот же самый Карпов систематически называет волоски на жгутике хоанофлагеллят именно мастигонемами - следует ли считать, что это неверно? И насколько сильно отличаются vanes хоанофлагеллят от vanes губок?
2. Прошу прощения, но в статье по ссылке я нашел нечто другое, а именно утверждение, что ленты микротрубочек (microtubular bands) у хоанофлагеллят _есть_. Как раз в разделе сравнения там написано, что "choanoflagellates have, choanocytes have not radial microtubular roots organized in bands". Но из описания и многочисленных рисунков видно, что у губок от основания жгутика тоже отходят латеральные микротрубочки, пусть и не организованные в ленты. Разница есть, но не такая уж принципиальная. Если мне придется републиковать этот материал, я, конечно, уточню терминологию.
3. Основная идея статьи по ссылке, как я ее понял - доказать, что у хоанофлагеллят и губок ультраструктура жгутиковых клеток не идентична. Это ценная информация, но ведь то, что губки не являются прямыми потомками хоанофлагеллят, понятно и так.-
1. upd: у хоанофлагеллят мастигонемы структурно представляют собой что-то вроде гликокаликса, т. е. они крепятся к мембране по всей поверхности (ссылаюсь на Карпова), а у губок это выросты мембраны, организованные в продольные гребни. Терминология плавающая: кто-то говорит vanes, кто-то - mastigonemes. Кстати, у личинок губок они тоже бывают! Например, у пресноводных Spongillidae.
2. Всё верно, но тут номенклатурная заковырка. В клетке есть поперечный цитоскелет жгутикового аппарата - это радиальные микротрубочки, а есть продольный - это фибриллярные корешки, которые могут быть исчерченными. У многоклеточных под roots подразумевается именно продольный скелет обычно (отсюда формулировки из моего первого комментария), но латеральные пучки микротрубочек у протистов часто тоже называют roots. Суть такая: мы не можем сказать, что у хоанофлагеллят и губок - микротрубочки, а у всех остальных - исчерченные корешки, потому что это не антитезы, а разные структуры. У хоанофлагеллят помимо этих микротрубочек есть и исчерченные корешки, а у губок помимо фибриллярных корешков (исчерченных или простых) есть и микротрубочки. Статья по ссылке была призвана это продемонстрировать, но, наверное, только сильнее запутала :)
-
Но взгляд споткнулся на фразе "Человек вышел из животного царства". Понятно, что это оборот такой, но может было лучше написать "человек, являясь частью царства животных, постоянно сталкивался с другими животными". Царство тут может быть и не таксономическое понятие, если оно сейчас расплывчато.
-
"Между Большим взрывом, запустившим образование химических элементов, и появлением условий, подходящих для возникновения жизни, во Вселенной произошло множество событий. Эти события складываются в космическую (неорганическую) эволюцию. С другой стороны, человек — главный герой исторического процесса, в ходе которого биологические изменения его организма перекрываются культурной наследственностью, действующей через обучение. История человечества связана в первую очередь с эволюцией культуры. Три эволюции — космическая, биологическая и культурная — составляют единый грандиозный процесс универсальной эволюции" (Dobzhansky, 1968).
Человек, разумеется, является в таксономическом смысле животным, так же как, например, вообще любая живая система является химической системой. Но его природа к этому не сводится. Кажется, это такая же тривиальность, как и то, что человек произошел от обезьяны. Впрочем, с точки зрения темы данной статьи это вообще неважно. -
Если взять такой подход, и развить до предела, то Человек образует новое царство - разумные преобразователи окружающей среды или что-то такое :) Причем эта группа может стоять даже выше царств, выше даже разделения живого и неживого. Так как разум, вполне вероятно, может быть и исусственный.
В этом смысле "вышел из царства животных" можно понимать, не как "возник из животных" (как я понимаю, это именно смысл фразы в статье), а как "покинул Животных, став выше их" (а это уже креационисты могут так понять).
Не сочтите за придирки. Статья очень хорошая! Это просто от чего-то я в эту фразу уперся.
-
Ни Добржанский, ни любой другой грамотный биолог не может отрицать, что человек остается животным. Смысл и высказывания Добржанского, и моей фразы в том, что он _не только_ животное, так же, например, как живая клетка - _не только_ химическая система. За придирки извиняться не стоит, когда они конструктивны - это вполне по делу. Но нельзя ж что ни шаг бояться креационистов, которые могут не так понять!
Спасибо за высокую оценку статьи, я в нее много сил вложил. -
Вы не поверите, но некоторые исследователи берут и именно так делают, утверждая, что систематическое положение человека выглядит следующим образом: царство Psychobionta, тип Psychozoa, класс Homines, отряд Verbales.
-
Интересно, это кто ж так делает? Я, если что, противник такого таксономического решения.
UPD. Тип — политерии (Politeria), порядок — соклетные (Syncytialia), класс — метаморфные (Metamorpha)... Но в то и дело, что с человеком ситуация принципиально другая.-
Я тоже не то чтобы поддерживаю, просто говорю о том, что всё уже придумано до нас :)
Вот тут, например, об этом можно прочесть: https://books.google.ru/books?id=mk4IDAAAQBAJ&pg=PA244&lpg=PA244&dq=Psychobionta+Psychozoa&source=bl& ;ots=592G3iwW0j&sig=ACfU3U1iYRFoWeC7INbQ8GIsdQCWA08iVQ&a mp;hl=ru&sa=X&ved=2ahUKEwj15NHoiOflAhVElosKHRlWCOIQ6 AEwCHoECAIQAQ#v=onepage&q&f=false
-
-
А то, что человеки - далеко не просто очнредные приматы ясно всякому "имеющему глаза". По экологической роли на уровне биосферы нынешние человеки уступают разве что только покрытосеменным.
-
А то, что человеки - далеко не просто очнредные приматы ясно всякому "имеющему глаза". По экологической роли на уровне биосферы нынешние человеки уступают разве что только покрытосеменным.
Ну например роль муравьев тоже огромна, что значит они уже не перепончатокрылые ?-
Перепончатокрылые, конечно. Но они _эусоциальные_ перепончатокрылые. А вот это вполне тянет на принципиально новый уровень организации живой природы. См. Уилсона.
-
Завоевание Земли читал признаю очень хорошей книгой, но суть в другом есть критерии систематики- пусть кладистики и если человек это род семейства гоминид близкий к роду шимпанзе значит так и должно быть. А всяких новых придумщиков что выдумывают ересь, в одну клетку с новыми хронологами и фонатами плоской Земли
-
-
-
И птиц. Чтож - они уже не пресмыкающиеся (диапсиды)?
И млекопитающих. Чтож - они уже не синапсиды? ;~]-
Ну птицы и млеки вполне себе обособленые групы на кладограмах
( монофилитические таксоны) а с рептилиямы вы правы тут беда черная :)-
птицы и млеки вполне себе обособленые групы на кладограмах
Род Homo - тоже монофилетический. ;~]
( монофилитические таксоны)
Но на каком основании в одном месте присваиваются ранги семейства, отряда или класса, а в другом - только лишь род?а с рептилиямы вы правы тут беда черная
Да какая там больно беда: достаточно просто заменить их на диапсид с архозаврами в качестве подгруппы. Вот с амфибиями беда куда большая хотя бы из-за лабиринтодонтов.-
Род Homo - тоже монофилетический. ;~]
Ну например сравнить генетическое расстояние ( дистанцию) коэффициент Нея + сравнить анатомию морфологию и ты ды,
Но на каком основании в одном месте присваиваются ранги семейства, отряда или класса, а в другом - только лишь род?
мы и так делаем поблажку для человеков по хорошему мы и шимпанзе ОДИН родДа какая там больно беда: достаточно просто заменить их на диапсид с архозаврами в качестве подгруппы. Вот с амфибиями беда куда большая хотя бы из-за лабиринтодонтов.
1. Нужно отрезать птиц и млеков таксон парафилитический
2. Может я пропустил неужто ВСЕ рептилии происходять из одной группы амфибий ( интересно почитать)-
Ну например сравнить генетическое расстояние ( дистанцию) коэффициент Нея + сравнить анатомию морфологию и ты ды...
А критерии этого разделения в каэдом случае будут выставляться также по желанию левой пятки таксономиста? Объективность тут где?Нужно отрезать птиц и млеков таксон парафилитический
С чего бы?? Млеки и так отрезаны в синапсиды, а птиц из диапсид никто и не исключал. "Пресмыкающихся" нужно либо полностью отждествить с диапсидами, а лучше - просто выкинуть.Может я пропустил неужто ВСЕ рептилии происходять из одной группы амфибий
Хуже - выходящих на сушу четвероногих была не одна линия. И как они меж собою связаны - чёрт ногу сломит.
А "рептилии" тут нипричём - дальше появляются _амниоты_, и они вполне монофилетичны.
-
-
-
-
-
-
-
Ну это Вы архея. А я, может быть, и нет:)
На самом деле, учитывая бактериальное происхождение 75% эукариотного генома и полную пересборку бактериального и архейного компонентов при симбиогенезе... не стал бы я говорить, что эукариоты - это археи. Эукариотизация - идеальный пример "морфологической сингулярности", о которой давеча говорил доктор Кузнецов в докладе (см. мой ЖЖ). Преемственности по отношению к архейному состоянию, за исключением генетической, там почти что и нет.-
Пришла к зайчику в избушку лиса, да его же и выгнала ;)
Спасибо, поглядю, пока не успел.
ЗЫ я успел приобщиться классической школе биологии (слава Саратовскому Университету! гип-гип и все такое) - и потому кладистику не люблю всей душою (но, хоть и морщася - да тру (с)), а люблю старую добрую биологию, которая умела осмысливать переходы в уровнях организации систем, и подстилающую ее диалектику, которая "изменяется с каждым новым научным отрытием (с)". Однако же, иногда можно и потроллить ;)
-
-
-
Это просто элементарный рибошовинизм тогда получается. Тут уже нет и следа объективности - чистый призвол: почему одни гены оказываются "равнее" других - не объясняется ниразу.
-
Это как раз элементарно - просто шовинизм не рибо-, а репликаторо-.
И как бы ни косили глазом презренные коацерватники, суть и основа жизни - репликаторы, и никому мы ее ниададим!-
Полноценный репликатор - минимальная клетка - имеет довольно немалый минимальный набор генов - генов "домашнего хозяйства". Убери один-два - и никакого самостоятельного репликатора уже не получится (для несамостоятельных гены вообще не нужны - см. вироиды, монстр Шпигельмана и т.п.).
У эукариот часть этих генов имеет архейное происхождение, а часть - и не малая - бактериальное. На каком - объективном, а не произвольном - основании преимущество в оценке родства отдано первым?
-
-
-
-
Можно и психозой с вербалиями выделять, а можно вводить систему на основе цветовой дифференциации штанов, предварительно приведя физиологию цветовосприятия в связь с культурно-социальными параметрами и критериями. Ненуачё.
Сопсно говоря, что угодно можно делать.
Вопрос один - зачем?
Но событие очень редкое. Проще представить плавающее сообщество воротничковых жгутиконосцев, которое потом стало трихоплаксом.
-
Ну, есть несколько неотенических семейств хвостатых амфибий: протеи, сирены и тому подобное. Частный пример, но зато не вызывающий ни малейших сомнений. Причем возможно, что даже предки всех хвостатых амфибий были неотеническими - я об этом писал: https://elementy.ru/novosti_nauki/432513/Genomy_khvostatykh_
amfibiy_s_samogo_nachala_byli_bolshimi
UPD. Нет, не проще. Это не объясняет сходства взрослых губок с воротничковыми, а личинок губок с эуметазоями. А неотеническая гипотеза прекрасно объясняет.
У самих воротничковых ее нет. Все это, вероятно, означает, что жизненный цикл общего предка воротничковых жгутиконосцев и многоклеточных животных был гораздо более сложным, чем жизненный цикл современных воротничковых (который вторично упрощен).Может не более сложным, а другим? Зачем одноклеточному синхронность при дроблении? Даже в колонии? А многоклеточный отъел яйцеклетку, и делит её на накопленном , синхронно для синхронизации стадийности между всеми клетками будущего тела. Или тот абзац тоже про "Brachyury" ?
-
Ну что другим - это наверняка. К сожалению, все промежуточные стадии тут, похоже, вымерли, и это очень усложняет дело. Даже половой процесс у воротничковых открыли только в 2013 году - до этого было непонятно, есть ли он у них.
-
ачем одноклеточному синхронность при дроблении
Как ни странно, по факту эта синхронность произошла именно на стадии колониальности, породив ценобий как противоположность иррегулярной колонии.
Кстати, дарю (бэз-вазд-мэзд-но! пчхи!) свою любимую давнюю идею - опосредованный теломерами лимит количества делений клеток как наследство ценобиальной организации.
-
-
-
У Лема там железные мухи были - это из другой оперы. Он это написал очень давно. Сейчас уже очевидно, что в недалеком будущем появятся автономные, человекоподобные роботы. Они постепенно будут замещать человека во всех сферах деятельности. Уборщики, строители, врачи, учителя, писатели... Пока наконец не заменят инженеров и ученых. Просто ученый-робот будет намного мощнее ученого-человека. С этого момента люди станут пройденным этапом эволюции. Многие не понимают, что до этого момента осталось всего несколько десятков лет. Остановить процесс вряд ли возможно...
-
1. А зачем вообще делать человекоподобных роботов.
2. Сейчас Вы воспроизводите Винджа с его технологической сингулярностью. Ну посмотрим.
Правда, не знаю, какое это все имеет отношение к теме статьи:)-
Извините, просто захотелось поболтать, тут в дискуссии разные виды эволюций упоминались. Спасибо за статью, очень интересно читать было и это очень приятно, что эволюция живых организмов настолько хорошо изучена.
-
Да это пожалуйста.
Я просто зацепился за Вашу фразу насчет _человекоподобных_ роботов. Сколь я вижу, их никто особенно не рвется делать - а зачем? По отношению к технологическим задачам, которые приходится выполнять роботу, человекоподобная форма - случайный побочный продукт чужеродной ему эволюции. Если делается, например, робот-пылесос, то ему и придается форма пылесоса. И уж тем более мне кажется исчезающе маловероятным, что автономная эволюция техногенной "жизни" (а-ля "Непобедимый" создаст что-то человекоподобное самостоятельно. Совсем наоборот, я думаю, что она будет абсолютно неантропоморфна. Не обязательно миниатюризирована, как у Лема - но главная его идея верна.-
а зачем?
Позвольте не согласится, навскидку два аргумента —один этичный, другой нет.
У человеческой анатомии перед пылесосом/тележкой и т.п. есть как минимум одно преимущество — высокая проходимость, особенно в зданиях, на лестницах и прочих созданных людьми для людей ландшафтах.
А теперь представим, что мы можем создать что-то человекоподобное (и не уступающее человеку по силе/выносливости) наподобие "суррогатов", посадим в костюм управления пожарного, обеспечим минимальный пинг в обе стороны, и отправим эту МАШИНУ (которую труднее поломать (как минимум она не может отравиться угарным газом/задохнуться) и в случае чего можно починить) вытаскивать имущество/людей из горящего здания.
Число жертв пожаров пойдет на убыль, и семьям погибших на службе МЧС-ников не надо будет платить пособия (да, я понимаю что роботы все равно обойдутся дороже, но все-таки…).
Туда же спецназ/оперативников полиции (если у преступников нет способа перехватить управление).
Возможно, можно найти более эффективную для этого анатомию, чем человеческая, но ее еще надо найти.
И другая крайность — автономные, но жестко запрограммированные роботы-слуги. Да, можно нанять людей, но роботу можно в принципе придать любую внешность (и "в теории" любую модель "поведения"), а получится ли у кого-то нанять мыть посуду голливудскую кинозвезду, к примеру (и на бесчеловечное обращение машина жаловаться никому не будет, как где-то у Азимова про роботов — "они — наши рабы"). Неужели в мире нет богатых людей с самыми разными "причудами"?
Так что какой-то спрос на андроидов/гиноидов, наверняка бы был (хоть как на "игрушки" для желающих почувствовать себя рабовладельцами), а вот покрыл бы он затраты на разработку, вопрос конечно хороший.-
Затраты на разработку... Многие изобретения вначале были неокупаемы. Но разве это остановило прогресс? Первые роботы будут ужасно дорогие, но ведь они же подешевеют очень быстро. Компьютеры в 80-ых стоили дороже автомобилей и даже квартир. А если какие-то фирмы или какие-то страны решат не создавать роботов, то их обгонят другие фирмы и другие страны, которые захотят.
Применений роботам столько, что не перечислить. Например, военные - зачем убивать людей, когда воевать друг с другом могут роботы? Робот-врач. Робот-строитель. Робот-журналист. И наконец ученый.
А если кто-то будет создавать роботов нарочно ограниченных и глупых, чтоб не вышли из-под контроля, то всегда найдется кто-то, кто будет создавать роботов без этих ограничений. Ведь такие роботы мощнее. Скажем, в США в роботов засунут законы Айзимова. А в России или Китае на это забьют.-
Первые роботы будут ужасно дорогие, но ведь они же подешевеют очень быстро.
Доброго Вам утра!
Роботы уже далеко не первые - они УЖЕ ЕСТЬ полвека как.
И никакой муровской динамики в их отношении не наблюдается.
От Z3 Конрада Цузе до "закона" Мура прошло менее 25 лет, как и от метода Сэнгера до появления систем NGS.
От роботов Юнимейт до сего дня прошло уже более полувека - и всё что мы видим - прототипы-демонстраторы технологий Boston Dynamics по почти астрономической стоимости.
Муровская динамика работает ТОЛЬКО в отношении систем получения и обработки информации. Эффекторные системы - обратно воздействующие на физические объекты - ничего такого не демонстрируют. Иначе мы бы уже летали меж континентами за считанные центы за билет и за секунды в пути.-
-
Ну справедливости ради не 10, а всего около пяти и только в своём исходном виде. Что она замедлилась с 2 до 2,5-3 лет - давно понятно.
Речь как раз о том, что даже если брать за отправную точку наиболее оптимистичные сценарии, транслируемые производителями в СМИ - то и они уже "приказывают долго жить". Но это происходит в ближайшие годы, а не 10 лет назад.-
Даже более того, Мур сам сменил показатель степени в своем "законе" с одного на два года в 70-х годах.
А сейчас уже идет втискивание в этот квазизакон.
Экспонента развития была в связи со сменой масштаба на несколько порядков элементной базы от ламп к транзисторам. Никакого универсального или информационного значения эта эмпирическая закономерность не имеет.
Не надо плодить желтое мракобесие.-
Тем не менее после этой коррекции и в том исходном значении (количества транзисторов на чип / себестоимости одного транзитора и т.п.) эта динамика весьма неплохо соблюдалась до самых последних лет, что удивило даже самого автора. То, что правило Мура действовало полвека к ряду - это эмпирический факт и пытаться как-то его размазать, это всё равно что говорить, что крокодилы тоже в некоторой степени летают, раз вместе с птицами относятся к архазаврам, только "нызэнко-нызэнко", да и птицы тоже не сказать, что сильно высоко - всего лишь не выше тропосферы.
Не вижу в этом смысла, тогда как дискурс "жёлтого мракобесия" мы в ряде случаев к сожалению вынуждены принимать во внимание как стартовую позицию и тут далеко не такой критический случай, чтобы нужно было столь сильно проявлять технический ригоризм.
-
-
-
-
-
-
Я в своем комментарии писал о случаях, когда может быть целесообразно использовать конструкцию роботов, повторяющих именно анатомию человека (вплоть до неотличимости визуально/тактильно) (да, возможно не все из них), а не про танк/истребитель/лабораторию с искином внутри (зачем робот-танкист и робот-летчик? Если мы можем создать ИИ способный управлять этой техникой, почему бы не внедрить прямо в само устройство без лишних "посредников", в итоге получится проще, дешевле и надежнее).
И касательно сделанных мной оговорок про неавтономность и жесткие программы.
Глупых роботов делать явно проще и дешевле, чем умных, а тому кто полезет под пули/в огонь/в полуразрушенное здание/под обвал в шахте автономный интеллект вполне может мешать (3-й закон и все такое (в духе "Я, ТУДА, нет, не пойду"), а если системы самосохранения нет, то вдруг сие чудо техники полезет в самое пекло, и пропадет безо всякого проку). Ну и наделять роботов-слуг/игрушек (фактически, рабов) разумом (а не способностью поддержать диалог на уровне "блондинка из анекдотов"/современный чат-болталка, понимать и выполнять ПРОСТЫЕ приказы) — это уже форменный садизм. И тут помимо вопросов в духе "почему ты используешь визуальную копию своего врага/соперника/обидчика в качестве боксерской груши/вешалки для пальто/приставки к лотку кота/проч.?" возникнут куда более серьезные.
И да, у самого Азимова прекрасно описано, что три закона не всегда работают как задумано.-
Какие законы Азимова? Это утопическая фантастика. Реальные дроны в первую очередь запрограммированы на убийство.
-
Это был ответ на то, что где-то включат законы Азимова — что даже ЕСЛИ это сделать, не факт, что поможет обезопасить от причинения роботами вреда людям. Я не утверждал, что их кто-то включит в боевые машины (что само по себе нонсенс).
-
Реальные роботы не включают законов Азимова. Ни боевые, ни на конвейере, никакие другие. Смотрите вокруг внимательно. И судите не по словам фантастов, пусть и замечательных, а по реальности.
-
Я и не утверждал, что включают (как минимум чтобы они работали, машина должна быть способна оценить вред), я ответил на утверждение leonid_ge, что их ВКЛЮЧАТ контраргументом, что это (даже по словам их автора — не всегда возможно определить последствия плюс определение "человека", и наверняка еще можно много нюансов найти) не обеспечит безопасности , даже ЕСЛИ случится, и я не просто так уже во втором сообщении выделяю слово "если".
ПС. Собственно, я свой первоначальный ответ по законам и писал из соображений и в духе "Даже внутри "родных" НФ-романов это не работает так, как декларируется".
-
-
-
-
Любые реальные современные боевые системы, стратегии и тактики технически-развитых регулярных армий, а не отрядов террористов, запрограммированы не на убийство, а на НЕЙТРАЛИЗАЦИЮ боевых единиц, соединений и систем снабжения противника как можно меньшей ценой для собственных сил.
Какие-то убийства и разрушения при этом - просто более или менее приемлемый (в зависимости от политической обстановки) побочный эффект. -
-
-
-
-
-
-
Роботов создают самых разных. Одни фирмы делают роботов для конкретных задач, но другие пытаются создать универсального, похожего на человека. Поскольку такой робот будет действовать в среде, где существует человек, то по принципу "конвергентной эволюции" он в итоге будет очень похож на человека. (Имеется в виду технологическая эволюция, когда разные компании по всему миру стараются создать универсального робота).
Неуниверсальный, специализированный робот хорош для определенных задач, но он всегда будет сильно зависеть от человека. Например, его никогда нельзя будет послать на Марс. Мощь человека именно в том, что он универсален. И к этому будут стремиться компании, создающие роботов по всему миру. И они его в итоге обязательно создадут, не может быть иначе, это только вопрос времени.
А когда появятся наконец эти универсальные человекоподобные роботы, они очень быстро станут мощнее человека во всех областях человеческой деятельности, просто потому, что мозг человека не эволюционирует больше, а "мозг" роботов будет эволюционировать с нарастающей скоростью.
Конечно же, ничего подобного железным мухам в "Непобедимом" не может возникнуть само по себе. Техногенной автоэволюции сейчас нет. Всю техноэволюцию делает человек. Пока. Пока он не сделает себе замену, а это неизбежно. Вот тогда и начнется новая эволюция. Роботы-инженеры, роботы-ученые будут делать все более умных роботов.-
Неуниверсальный, специализированный робот хорош для определенных задач, но он всегда будет сильно зависеть от человека. Например, его никогда нельзя будет послать на Марс.
Снова доброго Вам утра! Кагбе на Марсе уже под десяток специальных роботов каталось за прошедшие десятилетия. И не бегало ни одного марсонавта.Всю техноэволюцию делает человек. Пока. Пока он не сделает себе замену, а это неизбежно
Своим высшим мыслительным функциям вполне возможно что и сделает и даже превосходящие в разы, а вот своим связанным с универсальным мозгом универсальным и сравнительно очень ресурсно-дешёвым рукам - нет.
Как не сделал и не сделает массовую замену пищи себе не из продуктов живой природы.
Как не сделала природа за миллиарды лет замену хлипкой и неуклюжей, но универсальной РНК в клетке.-
Про компьютеры тоже раньше думали, что они всегда будут узко специализированные. А что вышло? Вон ваш смартфон. В нем и телефон, и видеотелефон, и фотоаппарат, и телевизор, и электронная почта, и газеты, и книги, и энциклопедии, и навигатор, и соц. сети... (я что-то упустил?)
И всё это за 40 лет - в 80-е компьютеры были узко-специализированные, как сейчас роботы.
Вот и представьте, какие будут роботы через 40 лет.-
Смартфон - миниатюризированный комбайн со специальными функциями. Никакой универсиализации в нем я не вижу.
Результат - фотАчки котиков и лайки хомячков.-
"Универсальный - Пригодный для многих целей, выполняющий разнообразные функции" [словарь]
И робот, который будет и уборщиком, и слугой по дому, и врачом, и преподавателем, и инженером, и журналистом - тоже будет комбайн.
Да и человек - такой же комбайн.-
Нет. У человека могут быть свои цели. Он может продуцировать и цели и методы их достижения. В этом универсализм.
Вся беда с ИИ пока в том, что ИИ должен быть универсальным, разносторонним. Иначе он неизбежно упирается в рамки области функции определения.
Смартфон не способен квасить капусту.-
У робота-пылесоса есть свои цели. И у таракана есть свои цели. У человека [по его мнению] цели возвышенные. Это определяется только сложностью.
Пока что робот-пылесос в своих целях и по сложности уступает даже таракану.
Но подождите. Роботы эволюционируют со скоростью в миллион раз большей, чем биологический вид Homo sapiens. Когда они станут достаточно сложными - и цели у них будут соответствующие. И на это уйдет не 40.000.000, а 40 лет.-
У робота-пылесоса нет цели. Это робот только в коммерческом названии. Это автомат.
Роботы пока не эволюционируют вообще. Они создаются людьми. Реальный ИИ пока существует (а точнее создается) отдельно от роботов.-
Робот-пылесос - автомат. И таракан-автомат. И человек-автомат. Цель робота-пылесоса - пропылисосить комнату. Пока цель не достигнута, робот работает. Когда комната убрана - он останавливается, цель достигнута.
Не важно, создается ИИ отдельно от роботов или вместе и вообще, где и как он создается. Важно то, что роботы эволюционируют. Так же, как эволюционируют компьютеры, телевизооры, самолеты, часы, стулья, одежда и т.д.
Оглянитесь вокруг. Любой "созданный человеком" предмет прошел долгую эволюцию. Стул, на котором вы сидите, лампочка, очки, - любая вещь.
Абстрагируйтесь от биологической эволюции с ее мутациями в ДНК и естественным отбором. Не важно, каким образом идет эволюция, главное, что она идет - самолеты, компьютеры, чайники меняются, совершенствуются, усложняются или упрощаются. Часы эволюционируют уже 3000 лет (от солнечных).-
-
По теории автоматов, человек, как и любой биологический организм, - это конечный автомат.
Интеллект - это не чудо и не "нечто мистическое и непознаваемое". Интеллект может быть формализован.
Интеллект человека работает на нейронных сетях. Да, нейронов 10 миллиардов, и это много, но с дальнейшим развитием компьютеров весь человеческий мозг можно будет смоделировать и запустить на компьютере.
-
-
-
Цель робота-пылесоса - пропылисосить комнату.
Это вы изволили глупость спороть. У пылесоса нет цели. Цель была у человека, в данном случае у программиста, который написал управляющую программу для робота-пылесоса
В качестве умозрительного примера, вот я возьму в руку камень и садану им по вашей голове. Вопрос: была ли у камня цель стукнуть вас по голове? Ответ: конечно не было. Цель была у человека, в данном случае, у меня.
Никакой принципиальной разницы между пылесосом, управляемым программистом, и камнем (в моей руке) не существует. Никакого интелекта нет ни в камне, ни в роботе-пылесосе.-
Вот интересно, вы биолог или философ? Вы тут излагаете по Канту, интересно, конечно. Цель есть у божественного провидения. Оно всё видитъ.
-
Упаси меня боже от философии! Я программист.
Нужно просто понимать, что в роботе-пылесосе, японском андроиде, и в любой программе, с которой вы общаетесь, сидя за компьютером, нет ни грана интеллекта.
На самом деле вы общаетесь не с программой, а с ее программистом, который в своей программе попытался ЗАРАНЕЕ предусмотреть всё, что вы будете делать с его программой, и для каждого вашего действия (нажал клавишу, сдвинул мышку и т.п.) подготовил некий однозначный ответ.
Пока что абсолютно весь интеллект сосредоточен в головах программистов, а также в головах математиков и прочих специалистов, которые разрабатывают алгоритмы. Вряд ли это принципиально измениться в ближайшие полста лет.
Алгоритмы распознавание текста и лиц - весьма не просты, но тем не менее это алгоритмы, и в вычислениях по этим алгоритмам ничего интеллектуального нет.
А вот чтобы литературно перевести Анну Каренину с русского на английский - вот это уже было бы нечто интеллектуальное. В 1960-х годах всерьез думали, что это будет сделано в ближайшие 10 лет. Но ничего не вышло. А сейчас об этом даже и не мечтают.-
Зря вы так скептически. Человек точно так же "запрограммирован", как и компьютер. Он делает только то, что заложено в его нейронных сетях – эволюцией и его жизненным опытом (а не программистами) - но это ничего не меняет.
Вам известен следующий классический опыт? При определенных травмах мозга весь интеллект и все способности работают, даже обучаться человек может, но он не может запоминать события. Так вот, к такому человеку в комнату входит врач и начинает с ним разговаривать:
- Как вы себя чувствуете?
- Немного спина ломит, а в остальном всё ок.
- Погода хорошая, не правда ли?
- О да, сейчас солнце, но после обеда обещали дождь.
И т.д.
Затем врач выходит и через минуту входит снова. И опять начинает разговор, используя те же фразы. И пациент отвечает АБСОЛЮТНО ТЕМИ ЖЕ фразами. Слово в слово. Этот опыт проводился много раз.
Вывод из него: человек так же запрограммирован, как и компьютер, а то что он не повторяет диалогов слово в слово - только потому, что он их помнит. Можно легко в Алексу заложить, чтобы она каждый раз немного по-разному отвечала на одни и те же вопросы.
На человеческий интеллект смотрят, как на чудо, и как на нечто непостижимое. Это только потому, что всех алгоритмов человеческого мозга еще пока не изучили. Раньше зрение казалось чудом. Теперь оно очень подробно изучено, и не только глаз и сетчатка, но и обработка изображения в коре мозга, и нейронные карты и т.д.
Погодите. Изучат и процесс формирования речи, и мыслей, и как работает сознание. И запрограммируют все это в компьютере, как уже запрограммировали распознавание изображений и речи.
Вы упомянули, что «общаясь с программой, вы общаетесь на самом деле с программистом». Это только для простых программ. Я ваш коллега. У нас в компании «программа» состоит из нескольких миллионов строк компьютерного кода, сотен библиотек, бежит на десятках серверов, «переговариваясь» друг с другом. Всё это писалось лет 15 примерно. Сотнями программистов. С кем конкретно из них я общаюсь, когда общаюсь с этой программой?
Человек отличается от простой программы только тем, что у него очень-очень сложная программа. Вы пытаетесь сравнить человека с простеньким ИИ, выполняющим небольшую конкретную функцию. Например, распознавание лиц. А вы представьте себе ИИ, который распознает не только лица, но и все предметы, и речь, и управляет телом робота, и может брать предметы, и выполнять работу, и помнит, что делал раньше, и может рассказать о том, что делал раньше. И может читать эмоции по лицам людей. И т.д. У него очень-очень много «простых» программ в распоряжении.
И на закуску. Просто представьте, что в какой-то момент смогут воспроизвести мозг человека в компьютере. Просто сделать полный скан мозга – абсолютно всех нейронов и связей между ними и симулировать все эти нейроны со всеми связями в компьютере. Получится копия личности человека в компьютере. Ее можно будет подключить к искусственному телу, видеокамерам, микрофону и т.д. Вот вам и человекоподобный робот. И его «мозг» можно будет потом бесконечно расширять и улучшать.
Насчет Анны Карениной – погодите немного. Google-translate пока что переведет ее не литературно, но понять этот перевод вполне можно. А до литературных переводов еще дойдет. Просто подумайте, как получается, что человек переводит Анну Каренину на английский? Во-первых, переводят обычно на родной язык. Переводчик сам прочел множество книг на родном языке. И на языке, с которого переводит, тоже. Он ПОНИМАЕТ текст - а не переводит, используя только словарь и правила грамматики, как гугл. Вот когда робот с ИИ будет существовать и обучаться среди людей, тогда и он сможет перевести Анну Каренину. Когда, читая «Князь Андрей увидел высокий дуб», этот робот нарисует в своем воображении этот дуб – вот тогда он сможет это перевести на английский. И это вполне возможно в будущем.-
Вы совершенно точно отметили, что для перевода нужно ПОНИМАНИЕ. Сейчас понимания нет даже при переводе пары предложений. Так что о ПОНИМАНИИ целого романа можно пока что только мечтать.
И, кстати, даже лица компьютер так и не научился распознавать. Это совсем не то, как человек распознает лица. Когда я узнаю знакомого человека на улице или на фото, я ПОНИМАЮ, кто это: как его зовут, где я его раньше видел, есть ли у него дети и т.д. И что важно : это МОЕ собственное представление о человеке. И есть много других людей, которые тоже могут узнать этого человека,и у каждого есть СВОЕ особенное представление. Это радикально отличается от нынешних баз данных, в которых хранится пара фоток и сотня слов (год рождения и т.п.).
То, что сейчас называют распознаванием (лиц, текста и прочего) - это просто условные технические термины. В этих программах нет интеллекта.-
Ну вы как в детском саду. ИИ, распознав лицо человека, вытаскиваете из базы данных запись, откуда узнает, как человека зовут, есть ли у него дети и вообще все данные о нем (в базе данных ФСБ о любом человеке данных больше, чем у вас).
Насчёт перевода - кто же спорит? Как только ИИ будет существовать в роботе, который будет жить в среде, в которой живут люди, тогда и он сможет понимать и литературно переводить. Что вы от него сейчас хотите? Дайте охотнику из племени пигмеев словарь и Анну Квренину на русском и попросите перевести на английский. Вон он человек, не ИИ, думаете переведет литературно?-
Жаль, что вы не читаете то, что я вам пишу,смотрите, например,то, что я написал о базах данных.
И я вынужден еще раз повторить: база данных ФСБ со всеми ее программами не имеет никакого интеллекта. Есть люди, которые запускают поиск, и они понимают, чего или кого они хотят найти. Сама по себе БД - это просто куча битов на диске. Она, БД, не понимает зачем и кого она ищет. Это может быть и матерый убийца и потерявшийся ребенок - для программы нет разницы. А для человека, который эту программу запускает, разница есть, и ПОНИМАНИЕ есть.
Вот возьмите учебник по высшей математике. Вы же не будете утверждать, что эта куча белых листов бумаги с какими-то темными пятнами на них ПОНИМАЕТ высшую математику. Так и с программами.
Не только пигмей, но и вы не сможете литературно перевести Анну Каренину. И я не смогу. Вот просто задумайтесь: почему?
А дело в том, что мозг отдельного человека со всем его интеллектом - это НИЧТО. Человек - существо общественное. Такие вещи как язык, культура, искусство - они"размазаны" по головам миллиардов людей. И настоящий ИИ возможен только на уровне общества, а не отдельной головы.-
Елки-палки, а я о чем? Третий раз вам пишу: когда ИИ будет в роботе, который будет жить в обществе людей, тогда этот ИИ сможет подняться на уровень человека. Сейчас такого пока естественно нет. Когда появятся роботы, андроиды, с руками и ногами, которые будут способны жить среди людей, тогда у них и разовьётся ИИ как у людей.
-
Вы изначально подходите к человеку как к необъяснимому чуду. ИИ все равно, кого она нашла в базе данных: убийцу или гения-поэта, а человеку не все равно.
Во-первых, если человек не слышал об этом убийце или поэте, ему будет так же все равно, как и ИИ.
Во-вторых, почему вы считаете, что ИИ не может понять, что такое убийца? Представьте себе робота. Он читал в интернете про убийцу. И вдруг распознал его на улице по лицу. Разве робот не поймет, что этот убийца опасен? Робот может сообщить в полицию, что увидел убийцу. Он может принять меры по обезвреживанию убийцы. В чем проблема? -
Я понял, в чем у вас проблема. Вы слову ПОНИМАТЬ придаете мистический смысл. А вы попробуйте разобраться, что это слово значит. Что значит, "я понимаю, что этот человек убийца"?
-
Это называется квалиа, и человек впал в философскую петлю: является ли философский зомби личностью или нет. Естественно, обученная нейронная сеть не есть алгоритм, созданный программистами, потому что как она работает, ее создатели объяснить не могут, результаты слабо предсказуемы, а иногда парадоксальны. В процессе "обучения" этот алгоритм рождается внутри и зачастую абсолютно непонятен. В этом и беда искусственных нейройнных сетей - они непрозрачны
-
-
-
-
-
Предложение о сканировании мозга в компьютер выглядит хм механистически. Этот какой-то вульгарный материализм. Я думаю, что в лучшем случае мы получим маугли.
Но, скорее всего, мы не сможем адекватно отобразить в компьютере мозг и тело человека со всеми его тонкими гормональными настройками.
Если уж и говорить о копировании человека, то тогда делать это с точностью до атомов и молекул. Но это не имеет никакого отношения к программированию.-
Вульгарный материализм? А вы представляете мозг человека как нечто, недоступное пониманию и изучению (подход идеалистов)?
Когда вы делаете porting программы с одной платформы на другую, скажем с Windows на Linux, вы тоже каждый атом и молекулу воспроизводите?
Вы подходите с точки зрения знаний, имеющихся сейчас и пытаетесь доказать, что воспроизвести мозг человека в компьютере невозможно. Я этим я и не спорю. Сейчас - нет, конечно, нов будущем будет возможно.
Во времена Леонардо да Винчи тоже невозможно было построить вертолет, который он нарисовал.-
Когда вы делаете porting программы с одной платформы на другую, скажем с Windows на Linux, вы тоже каждый атом и молекулу воспроизводите?
Хороший пример. Вы при этом выступаете как программмист, вы понимаете что делает программа.
А вот исходная программа и портированная, какими бы сложными они ни были, ничего не понимают о себе: кто их сделал, кто портировал, зачем портировал, и прочее.-
Согласен. Porting мозга в компьютер сделают люди. А когда этот мозг заработает в компьютере, это будет ИИ.
-
Согласен. Porting мозга в компьютер сделают люди
А зачем портировать мозг человека в компьютер? И даже если это когда-то будет возможно, то сколько это будет стоить?
Вам не приходило в голову, что родить ребенка, выучить его и сделать человеком - это намного более простое и перспективное дело, чем портирование мозга в компьютер?
С какого, как говорится, перепуга вы вдруг решили, что супер-компьютер - это более подходящее место для интеллекта, чем ваши собственные биологичесские мозги?-
Вот! Конгениально, как говорил Остап.
А зачем редактировать текст в компьютере, когда писать на бумажке и проще, и быстрее, говорили в 1950-м году. А зачем рисовать на компьютере, когда на бумаге в сто раз быстрее и удобнее?, говорили тогда же.
А затем, что один раз портировав мозг в компьютер, его можно бесконечно размножать, копируя + улучшать, добавляя нейроны и их сети. А мозг в человеческой голове ни скопируешь, ни улучшишь.
В этом и есть преимущество компьютеризации - копирование, увеличение быстродействия, расширение. Вот почему компьютеризация проникла во многие области и в мозг тоже придет дайте только время.-
-
ОК. Как вы думаете, почему биологическая эволюция совершенно независимо создала несколько видов летающих животных: птеродактили, птицы, летучие мыши, насекомые (причем у множества летающих семейств насекомых полет возник независимо)?
Потому что были ниши для этого.
Для роботов-андроидов очень много ниш имеется.
-
-
-
-
-
-
А вы представляете мозг человека как нечто, недоступное пониманию и изучению (подход идеалистов)?
Мозг человека можно и нужно изучать. Но я думаю, что представление о мозге как о супер-компьютере - весьма наивно и поверхностно.
В 19-м веке истово верили, что всю вселенную, до каждого атома, можно однозначно просчитать вперед хоть на миллиард лет. Ваши представления о программировании из той же серии.-
Ваш пример с просветом вселенной вперёд на миллиард лет не совсем подходит к мозгу человека. Ну давайте возьмём более простой пример: можно ли симулировать на компьютере мозг (а точнее нервную систему) пчелы? Можно сделать робота-пчелу, заслать ее в улей и чтобы она там прижилась среди других пчел? (Допустим, этот робот сможет выделять соответствующие вещества и реагировать на запах).
-
Ну давайте возьмём более простой пример:
Вот-вот! Вы рассуждаете точно, как вульгарный материалист рассуждал: возьмем бильярдный шарик,стукнем им по другому шарику, и можно заранее сказать, как они будут дальше двигаться. И далее он делал некорректное обобщение на всю вселенную.
Я подозреваю,что ваши обобщения на тему программирования столь же не корректны. -
-
-
VladNSK, у нас интересная дискуссия.
Я честно, не могу понять, почему если в результате биологической эволюции на Земле возникло такое интеллектуальное существо, как человек, то в результате технологической эволюции не может возникнуть интеллектуальный робот. Я не вижу никакого принципиального препятствия, барьера. ИИ уровня человека и сложнее - нарушает законы физики?
Может, вы видите этот барьер?-
Нужно сначала строго определить что это такое - интеллектуальный робот. По-моему так ИИ уже есть. Это вы, я, наши дети, соседи.
Можно ли сделать ИИ на какой-то другой, не биологической базе? Не знаю. И никто пока не знает. Но я думаю, что супер-компьютер вряд ли подойдет.
И да, могут быть барьеры. Например, термо-динамические, квантовые и бог знает еще какие.
Вот для насекомых есть известный барьер : они так устроены, что не могут быть достаточно крупными.
-
-
-
-
-
-
-
-
-
-
У пчелы 1 миллион нейронов. У человека 100 миллиардов - в сто тысяч раз больше.
А память-то компьютерная тут причем?
Операционная система MS-DOS вначале имела памяти 640 КБ. Сейчас память в Windows 64 ГБ - тоже в 100 тысяч раз больше.
Важна суть алгоритмов, которые еще предстоит разработать. А алгоритмы рождаются в голове программистов и на листочках бумаги, на которых программисты что-то для себя рисуют.
Вы, Леонид, уже в который раз тут пишите нечто конкретное о программировании (названия операционных систем, сколько гигов памяти и т.п.), и мне, как профессиональному программисту с большим стажем сразу видно, что в программировании вы ровно ничего не понимаете. Только без обид!
64 гб памяти - это просто пустое место. И в это пустое место вы можете загрузить, например, Тетрис, и из того, что у вас внутри 64 гб будет работать Тетрис, ну, никак не следует, что Тетрис - это крутая супер интеллектуальная программа.
Никакой принципиальной разницы между Тетрисом, который крутился в 640 кб MS DOS, и тем, что внутри 64 гб-ой Windows, нет. И, кстати, MS DOS-ий вариант был лучше, потому что там скорость отклика на нажатия клавиш была выше. -
Нет, специально будут все на планете стараться, чтобы не дай бог не сделать ИИ. Будут всё что угодно изобретать, только не ИИ. Во всех странах поголовно примут закон, запрещающий изобретать ИИ. Заодно и исследования мозга запретят.
Вы, Леонид, должны понять, что то, что сейчас называют модном словом ИИ, и то, что вы лично под этим подразумеваете - это два совершенно разных понятия.
Нынешний ИИ, о котором сейчас везде стали трещать, это всё та же компьютеризация из 60-х. Дело безусловно хорошее, и этим нужно заниматься, но никакого настоящего интеллекта, в вашем и моем понимании, там пока что не предвидится. Увы! -
ОК. Как вы думаете, почему биологическая эволюция совершенно независимо создала несколько видов летающих животных: птеродактили, птицы, летучие мыши, насекомые (причем у множества летающих семейств насекомых полет возник независимо)?
Вот уж ...
Потому что были ниши для этого.
Для роботов-андроидов очень много ниш имеется.
А для розовых слонов с чайником Рассела там тоже ниша есть? Вопрос риторический.
-
-
-
-
-
-
-
Про компьютеры тоже раньше думали, что они всегда будут узко специализированные.Это когда это "всегда" было?? Со времён Тьюринга и фон Неймана были даны четкие критерии компьютера как универсального вычислителя. Остальные устройства называются словом "калькулятор" и пр.
И всё это за 40 летА роботы - и ныне там. Q.E.D.
Вот и представьте, какие будут роботы через 40 лет.Судя по предыдущим 60 - такими же специализированными промышленными машинами или редкими дорогими игрушками.
-
И всё это за 40 лет
То есть эволюцию компьютеров с 1990-го по 2020-й нужно судить по предыдущим 60 годам (с 1930-го по 1990-й)?
А роботы - и ныне там. Q.E.D.
Вот и представьте, какие будут роботы через 40 лет.
Судя по предыдущим 60 - такими же специализированными промышленными машинами или редкими дорогими игрушками.
Роботы просто позже начинают. Для роботов прежде всего нужны хорошие компьютеры, которых не было 60 лет назад. Но они есть сейчас.-
То есть эволюцию компьютеров с 1990-го по 2020-й нужно судить по предыдущим 60 годам (с 1930-го по 1990-й)?
Как минимум с 1965. Google: "закон Мура".Роботы просто позже начинают.
Т.е. вся сложнейшая, дорогая, но экономически-прорывная реальная промавтоматика начиная с автоматов фирмы Юнимейт 60-х годов и далее по промышленно развитым странам - это всё были игрушечки, а вот то, что всецело на основе всех этих наработок демонстрирует в своих дорогущих прототипах Boston Dynamics в последние годы на деньги своего правительства (DARPA) и венчурных инвесторов в отчаянных поисках какой-то прикладной рыночной ниши - вот это ТруЪ-начало робототехники и оно щас как попрёт, как попрёт?..
Выглядит не очень убедительно - не находите?-
Да вы просто представьте, что прилетели на машине времени в 1975-ый и показали там электронщикам и программистам айфон или айпад [правда, там интернета нет :(].
Не забудьте упомянуть, что такая штучка есть у каждого ребенка.-
...А также рассказать, что даже мильон таких штучек через полвека не проходит тест Тьюринга, и, будучи подключенной к соотв. манипуляторам, не сможет ни комнату убрать, ни ребенка перепеленать.
-
Ну а если слетаете в 2060-ый, смогут они и комнату убрать, и ребенка пеленать, и тест Тьюринга пройти. Прогресс не стоит на месте.
-
смогут они и комнату убрать, и ребенка пеленать, и тест Тьюринга пройти.
Конечно смогут. В виде прототипа фирмы типа Boston Dynamics за полмиллиарда долларов штука, который не пойдёт в серию по означенной причине и через десяток лет встанет экспонатом на вечный прикол в музее технической истории между Конкордом и Сатурном-5.
А также в 2060 наступит технокомунизм, молочные реки и кисельные берега.
Вот только эту пластинку играли как раз тем вашим слушателям - причём ещё до означенных 70х - причём всё поющееся обещалось к наступлению ДО нашего сегодняшнего дня. Уже в 70х она изрядно пожухла и годилась только для худфантастики.
Вы в курсе, что сейчас идёт тот месяц и год, в котором происходило действие фильма "Бегущий по лезвию"?
Чего-то по-новее за полвека не придумалось?
-
-
-
Тест Тьюринга - одна из тех совершенно бесполезных идей, из которых сделали идола и много лет поклоняются ему.(Еще одна такая же идея - кот Шредингера).Нет никакого смысла устраивать для ИИ слепой тест на выявление его отличий от человека.Это сродни тому, что в какой-нибудь биологической фирме решили бы принимать на работу только выпускников биофака МГУ и устраивали бы кандидатам слепой тест. Отличить выпускника биофака МГУ от выпускника биофака СПбГУ элементарно - спросить, какой профессор вел такой-то курс или расположение комнат в здании, или какие бутерброды можно купить в буфете МГУ. Вот только имеет ли всё это отношение к биологии?
Любой самый продвинутый ИИ всегда можно будет слепым тестом отличить от человека, точно так же, как и любых двух людей на земле всегда можно отличить слепым тестом. Всегда можно отличить американца от англичанина, москвича от петербуржца и т.д. Только какой смысл в подобном тесте при ответе на вопрос: "Хорош ли этот ИИ или плох?" Совершенно никакого. Тесть Тьюринга - никому не нужная бессмыслица.-
Тест Тьюринга - одна из тех совершенно бесполезных идей, из которых сделали идола и много лет поклоняются ему.
Вот только специалисты лингвисты, семантики и пр. разработчики успешных потребительских продуктов типа Помощник Google, Siri, Cortana и Alexa об этом не в курсе.
Только нужно не забывать, что всякая идея, чтобы быть плодотворной, не обязательно должна сходу (да и вообще) воплощаться в полной исходной своей форме. Попытки летать как птицы не привела к созданию серийного пассажирского орнитоптера, но разве это повод отрицать полезность самолётов?Нет никакого смысла устраивать для ИИ слепой тест на выявление его отличий от человека.
Зато очень даже есть смысл устраивать тест на комфортность диалогового интерфейса для потребителя.
Вы берётесь утверждать, что итоговый желаемый результат для обоих этих тестов НИ В ЧЁМ, НИКОГДА И НИКАК не совпадает?Еще одна такая же идея - кот Шредингера
Парадоксы КМ весьма наглядно моделируются парадоксами обычной, "чистой-классической" теории вероятностей. Оттуда становится ясно, что возникают они совершенно неизбежно во всякой дискретной вероятностной среде. Просто квантовый уровень реальности - именно такая среда: "Б-г" таки "играет в кости".
-
-
-
Скоро, видимо, образуется новое царство роботов. И начнется новая эволюция - уже не биологическая и не культурная, а роботная, когда наконец появится первый вид "робот разумный самостоятельный". В этой эволюции тоже, по всей видимости, будут присутствовать изменчивость, отбор, иерархия.Ерунда. Ставлю тыщу баксов на то, что автономная репликация на другой архитектуре кроме известных нам двух классов биополимеров с процессом трансляции с эффективностью и универсальностью хотя бы приближающейся к природным по порядку величины в этой вселенной невозможна.
Процесс РНК-полипептид (вне конкретного набора аминокислот, азотистых оснований нуклеотидов и кода их трансляции) ВСЕГДА будет лежать в основе полного цикла автономной репликации массивов наследственной информации с какими бы то ни было фенотипическими надстройками вплоть до цивилизации любой сложности, существующей сколь-нибудь геологически-длительный срок (хотя бы ярус).
-
-
Ставлю тыщу баксов на то, что автономная репликация на другой архитектуре кроме известных нам двух классов биополимеров с процессом трансляции с эффективностью и универсальностью хотя бы приближающейся к природным по порядку величины в этой вселенной невозможна.
То есть, репликация бинарного кода внутри компьютеров невозможна?
Кстати, речь была совсем не об этом. Эволюция роботов вовсе не пойдет по пути репликаторов, как биологическая. Зачем ей это? Разве технологическая эволоюция использует репликаторы? Массовое производство самолетов, компьютеров и холодильников работает по-другому. Вот таким же способом будут массово производиться роботы. На заводе.-
Вы правы с этим примером. Компьютерные вирусы - это уже реальность. Вот когда они станут себя сами модифицировать ... Так сказать мутировать.
-
Больше не станут: ЕМНИП, самомодифицирующийся код запрещён на уровне архитектуры процессоров чуть ли не с первых Пентиумов - как раз для вящей стабильности и безопасности систем.
Собственно этой же цели способствует и сама фоннеймановская архитектура и глубокое разделение функций РНК и ДНК (причём какая ирония - вторая произошла как раз от вирусов!) во всех земных клетках, ядра и цитоплазмы у эукариот и линий соматических и герминативных клеток у многоклеточных.
Евгений Кунин "Логика Случая", глава "Цикл Дарвина-Эйгена". Читать до просветления.-
Самомодифицирующийся код не запрещён. На него наложены некоторые ограничения, связанные с работой систем кеширования и предварительной выборки. Только начиная с Windows XP SP2 по умолчанию можно запретить запись в страницы с кодом и это ограничение легко выключается самим кодом. Так что технических препятствий нет.
-
-
-
То есть, репликация бинарного кода внутри компьютеров невозможна?
То есть автономная репликация самих компьютеров невозможна.
Вирусы не могут быть живыми системами отдельно от живых клеток (собственно как и самцы не считаются отдельными видами организмов).Кстати, речь была совсем не об этом. Эволюция роботов вовсе не пойдет по пути репликаторов, как биологическая. Зачем ей это? Разве технологическая эволюция использует репликаторы?
Именно. Строго говоря цивилизация - это не более чем расширенный фенотип человеков с его симбионтами (с/х культурами).Вот таким же способом будут массово производиться роботы. На заводе.
Человеками или его видами-потомками.-
Вот таким же способом будут массово производиться роботы. На заводе.
Вначале. А потом и самими роботами, интеллект и возможности которых превысят человеческие. На возникновение видов-потомков человека потребовались бы миллионы лет. А на возникновение разумных роботов несколько десятков лет.
Человеками или его видами-потомками.-
Вначале. А потом и самими роботами, интеллект и возможности которых превысят человеческие.
Видите ли в чём дело: как утверждает капитан Очевидность, интеллект сам по себе - это ВНЕЗАПНО не философский камень и не скатерть-самобранка.
Есть ещё и такая презренная наука как "экономика" (в части ресурсной себестоимости тех или иных производств), а в ней - такое понятие как EROEI - по итогу прямо вытекающее их действия законов таких наук как "физика" и "химия"...
Знаете и чего и как состоит РЕАЛЬНО ВЕСЬ ЦИКЛ проиводства каких-нибудь высокоточных актуаторов для роботов или станков? Какие химические элементы используются для производства высокотехнологичной продукции? Каковы их запасы (не только на Земле, но и в Солнечной системе в целом) и в каком виде они там лежат и как и чем происходит их добыча? Знаете ли Вы что значит термин "капитал" на самом деле? И почему в мире страны и компании-производители хайтех-станков можно пересчитать по пальцам (иногда - одной руки)?
Знаете ли Вы сколько сейчас осталось компаний производящих микросхемы по передовым техпроцессам (10/7 нм)? Знаете ли Вы, что случилось с мировым рынком жестких дисков после наводнения, затопившего один цех по производству шпинделей в Малайзии?
Если после познания этого всего Вы сохраните прежний техноэнтузазизм - это будет выглядеть интересно.-
Всё очень просто. Человеческий интеллект уже есть. Не нужно изобретать велосипед. Просто возможности человеческого мозга создаются в комьютере. Уже распознавание изображенией, в частности человеческих лиц ИИ делает лучше и быстрее человека. Распознавание речи уже работает. Генерация речи тоже. ИИ уже водит автомобили. Подождите пару десятков лет - и всё, что может человек (интеллектуально), вплоть до сочинения стихов, постановки диагноза больному, изобретениям, доказательствам теорем и т.д., - всё будет делать ИИ. А дальше - он будет это делать лучше и быстрее человека.
И что, скажите, тогда остановит появление автономного универсального робота, который заменит человека во всех сферах деятельности и который, наконец, станет улучшать самого себя и себе подобных?
Экономика, политика, государство и т.д. - всё это никуда не денется. Только ведущим звеном во всем этом будет не человек, а самостоятельный робот. Не из-за того, что роботы решат захватить власть, как в голливудских фильмах. А из-за того, что ИИ в любой деятельности превзойдет человека во много раз.-
Подождите пару десятков лет - и всё, что может человек (интеллектуально), вплоть до сочинения стихов, постановки диагноза больному, изобретениям, доказательствам теорем и т.д., - всё будет делать ИИ. А дальше - он будет это делать лучше и быстрее человека.
ОК - это всё замечательно.
А как насчёт чего попроще - ну например убраться в комнате чуть выше пола, перепеленать лежачего, сделать внутривенную инъекцию (не рассказать как, а именно СДЕЛАТЬ), аппендэктомию в любой райбольнице, принять роды, зашпаклевать и покрасить стены в комнате, ну и отремонтировать поломки у того же робота?
Таки как обстоят дела с автоматизацией труда уборщиц, строителей, медсестёр, ремонтников, а? А с экономикой их внедрения?Только ведущим звеном во всем этом будет не человек, а самостоятельный робот. Не из-за того, что роботы решат захватить власть, как в голливудских фильмах. А из-за того, что ИИ в любой деятельности превзойдет человека во много раз.
Ви таки не поняли. Речь не о "ведущем звене", а о _базовом_. Самэц - сколь угодно большой и умный - может быть главой сколь угодно большого прайда самок, но без них он оставит след разве что только в палеонтологической летописи в исторически-кратчайший срок.
Человеки без ИИ существуют преспокойно, но вот ИИ без облуживащих его человеков (или видов-потомков) исторически очень быстро станет кучкой вторцветмета.-
Вот именно поэтому и для этого роботы (или, если угодно, ИИ) станут универсальными и самостоятельными. Специализированный робот не сможет "убраться ниже пола", принять роды, зашпаклевать.
[Кстати, а вы можете принять роды и зашпаклевать стену?]
Именно в этом и дело, что наиболее экономически выгодный робот будет универсальный. То же самое произошло с компьютерами. Даже на МКС персоналки с Windows, айпады и айфоны, а не специализированные компьютеры.
Пока что компьютерам не хватает механики - они не могут двигаться и т.д. И поэтому человек пока остается универсальным существом, которое может "принять роды, зашпаклевать..."
Но это только пока.
Возможности человеческого мозга ограничены. Робот, который только что шпаклевал стену, может сразу же принять роды - через интернет скачает программу, как принимать роды. А человеку, чтобы принимать роды, нужно год учиться.-
Вот именно поэтому и для этого роботы (или, если угодно, ИИ) станут универсальными и самостоятельными.
А сингулярный технокоммунизм наступит к 2040 году, ведь "что нам стоит дом построить - нарисуем - будем жить"!
Кажется, мы слишком рано стали забывать слово "волюнтаризм". ;~]Кстати, а вы можете принять роды и зашпаклевать стену?
Как обладателю красного диплома медицинского ВУЗа, живущему в собственном доме, довелось принимать участие (что называется "на подхвате") и в том и в другом.
А ещё - уже целиком самостоятельно - находить и перепаивать вздутые конденсаторы в материнской плате, собирать и настраивать ПК (на уровне "аникейщика"), калибровать температурные датчики в промышленной теплице и pH-метры в своей нынешней лабе (профессия называется "КИПовец"), изготавливать микробиологические шпатели стеклогнутельством на газовой горелке, мастерить генератор сухого снега из углекислотного огнетушителя и пенопластовой коробки, затирать "заводскую прошивку" ДНК-праймеров в пробирках с реагентами ультрафиолетом как в старых EPROM-чипах и в какой только реальной продуктивной деятельности только не довелось поучаствовать благодаря судьбе, Родителям и Учителям...
Поэтому-то меня и столь умиляет регулярно наблюдаемый в интернетах техноэнтузазизм диванных робоконструкторов.Именно в этом и дело, что наиболее экономически выгодный робот будет универсальный.
Ещё раз: вы вообще в курсе из чего и как делаются прецизионные актуаторы для роботов и станков и как и откуда это всё берётся? И из чего состоит человеческий организм?Пока что компьютерам не хватает механики - они не могут двигаться и т.д.
Уже практически могут: https://www.bostondynamics.com/
Но Вы таки поинтересуйтесь - сколько это всё стОит. И тенденций к удешевлению "по Муру" там нигде особо не просматривается.То же самое произошло с компьютерами.
С компьютерами и секвенаторами произошла совершенно естественная вещь, ибо работа с информацией самой по себе - ресурсно весьма дёшева.
С роботами же дело обстоит существенно иначе.Робот, который только что шпаклевал стену, может сразу же принять роды - через интернет скачает программу, как принимать роды.
Ежу ясно, что если залезть на одну ступеньку, то можно забраться на всю лестницу.
Но вы покажите для начала хотя бы робота-шпаклёвщика, который вместе с обслуживанием и расходниками будет стоить меньше зарплаты бригады опытных строителей за две сотни лет при той же производительности и качестве. -
Но вы покажите для начала хотя бы робота-шпаклёвщика, который вместе с обслуживанием и расходниками будет стоить меньше зарплаты бригады опытных строителей
То же самое было справедливо для компьютеров 80-х. Редактировать фотографию на таком компьютере было не в пример дороже и невыгоднее, чем ретушировать бумажное фото. Я работал в начале 90-ых в институте с GPS-навигатором - прибор+софт был очень сложный и во много раз дороже атласа автомобильных дорог.
Вы по-другому смотрите на будущую эволюцию роботов, чем я. Вначале сделают андроида, который сможет передвигаться, брать предметы, манипулировать ими, как человек. Да, такого робота не возможно использовать, он страшно дорогой, никакой прибыли от него - одни убытки. И тем не менее его будут разрабатывать, потому что очень многие разработки делаются вовсе не для сиюминутной выгоды, а для науки и будущего. А когда разработают - вот тогда его можно будет научить шпаклевать, роды принимать и проч.
Ведь и мобильные телефоны, когда их разрабатывали в 90-ых, никто не собирался использовать в качестве видеокамер, GPS-навигаторов.
И перья у динозавров вначале были не для полета совсем.
Эволюция :)-
А когда разработают - вот тогда его можно будет научить шпаклевать, роды принимать и проч.
Но стоить он при этом будет всё так же - условно как Конкорд или Шаттл - и в итоге встанет на прикол в музее рядом с оными.
Когда строили конкорды и шаттлы тоже никто особо не думал, что космопланы и сверхзвуковые лайнеры окажутся тупиком технической эволюции.
-
-
-
-
Пока что компьютерам не хватает механики - они не могут двигаться и т.д.
Вы отстали от жизни лет на 20-ть минимум. Сейчас уже даже обычные школьники делают роботов. Сложность вовсе не в механике (с этим нет вообще никаких проблем). Сложность в написании алгоритмов, которые этой механикой должны управлять.
-
-
-
-
-
Уже есть принтеры, которые печатают сами себя.Не надо обманывать - нет таких принтеров.
Ну или прочитайте ещё раз исходное собщение, найдите там слово АВТОНОМНАЯ и если не знаете его смысл - посмотрите в словаре.
Принтеров таких не просто нет, но и не может быть размером меньше какого-нибудь крупного острова в силу катастрофы ошибок при копировании любой аналоговой системы (а макропринтер - такая система принципиально) - как прямое следствие действия второго начала термодинамики.
Ещё настоятельно рекомендуется ознакомиться с понятием порога Эйгена.
Уже есть принтеры, которые печатают сами себя.Принтер не может сам себя напечатать. Решение о том, когда конкретно начать печать, принимает человек. И при этом человек понимает, для чего он это делает, например, для прикола. А когда принтер работает, он не понимает, что конкретно он печатает: свою копию, копию принтера другой фирмы или бублик с дыркой.
А если серьёзно, то дискуссия получилась интересная. Насколько я понял, leonid_ge стоит на том, что человеческий разум подобен компьютерной программе, то есть не более, чем набор if/else, только очень-очень большой и сложный, а VladNSK утверждает, что всё не так просто. В принципе, аргументы leonid_ge местами очень убедительны, и хотя мне симпатична позиция VladNSK, толковых контр-аргументов (именно по этому вопросу), к сожалению, не прозвучало (или я проморгал).
Но вот есть у меня один вопрос. Если действительно возможно просимулировать (любым образом: с нуля, или просто скопировав) человеческий интеллект — каких вычислительных мощностей это потребует? Какая производительность должна быть у процессора или кластера, чтобы гонять такой алгоритм в реальном времени, сколько места всё это дело займёт, сколько будет потреблять энергии и выделять тепла? Кроме того, собранную и сгенерированную (ведь наш ИИ должен уметь не только собирать и выдавать информацию по требованию, но и обладать способностью к самообучению, то есть, генерированию информации, способности самостоятельно делать выводы и обнаруживать взаимосвязи) информацию нужно ещё и где-то хранить, причём, объём будет очень большой. Интернет, или другая подобная сеть, не обязательно глобальная, в принципе, вариант, но не такой уж хороший. Подозреваю, что было бы чрезвычайно смешно (или грустно...) наблюдать, как гипотетический андроид вдруг зависнет из-за невовремя оборвавшейся связи, лага или недостаточной скорости. К тому же, человек, как ни крути, в полном объёме базу своих знаний и впечатлений всегда носит с собой, в голове. Компромиссный вариант: доступ в сеть, но наиболее важную информацию хранить у себя постоянно (в принципе, как и у людей: "в голове не держу, но знаю, где посмотреть!").
Сам же отвечу на свой вопрос.
leonid_ge упоминал, что
>> У нас в компании «программа» состоит из нескольких миллионов строк компьютерного кода, сотен библиотек, бежит на десятках серверов, «переговариваясь» друг с другом. Всё это писалось лет 15 примерно. Сотнями программистов.
Но это всё равно тупая программа, жёстко выполняющая свои алгоритмы. Никакого интеллекта в ней нет. Она не способна создать что-то новое, она не умеет самообучаться, ставить перед собой цели и достигать их, и вообще делать что-либо, помимо того, для чего она предназначалась. Она большая, сложная, над ней долго и упорно работали, предоставили ей десятки мощных серверов — но Скайнета не получилось (к счастью!). И как-то сомнительно, что даже самые мощные суперкомпьютеры осилят такую задачу, как истинный искусственный интелект, хотя бы более-менее приближённый к человеческому. Нет сомнений, что в некоторых отношениях компьютеры оставляют человека позади. Но в других выигрывает таки человек.
Технический прогресс: разработка новых технологических процессов, выпуск всё более мощных и сложных процессоров — здесь не поможет. Технология не может всё время развиваться семимильными шагами. Рано или поздно она дойдёт до предела, после которого революций уже не будет. Возможно, вычислительная техника уже дошла до него. Уже нет таких прорывов и огромных скачков, что раньше. Весь прогресс сводится к увеличению количества ядер и памяти — идёт количественное развитие, но не качественное. Реализация искусственного интеллекта, если она возможна, потребует какого-то качественного скачка, чего-то принципиально нового. Можно вспомнить про квантовые компьютеры. Но во-первых, далеко не факт, что они способны на это, во-вторых, нет уверенности, что они вообще "взлетят", в-третьих, на данный момент они всё ещё находятся в предзародышевом состоянии и, по-видимому, ещё долго из него не выйдут.
И вот ещё один вопрос, на который я уже сам ответить не смогу. Допустим, более-менее литературно перевести книгу с одного языка на другой достаточно сложный алгоритм сможет. А вот сможет ли он сочинить новую? Картину нарисовать? Сонату написать? Изобрести что-либо новое, как человек изобрёл компьютеры и роботов?.. может даже замену себе же :)
Да, я знаю примеры процедурно сгенерированных текстов, музыки и изображений. Но все эти алгоритмы опять-таки основываются на заложенных в них их авторами шаблонах. Они не дают нового, только производят вариации старого. О настоящем искусственном интеллекте можно будет говорить только тогда, когда он научится изобретать и вообще делать что-то, что не было предусмотрено его создателями.
-
О настоящем искусственном интеллекте можно будет говорить только тогда, когда он научится изобретать и вообще делать что-то, что не было предусмотрено его создателями.
Поверьте программисту! Современные компьютеры очень часто делают то, что не предусмотренно их создателями. Из-за этого у программистов все проблемы.
Скачки в технологиях обязательно будут. Технологии все время скачут и чем дальше, тем быстрее. Проблема людей в общем (и многих писателей-фантастов в частности), что они совершенно не могут вообразить что-нибудь, что не похоже на уже существующее. Вы опасаетесь, что предел быстродействия компьютеров достигнут. Но обязательно появятся новые технологии, каких сейчас и вообразить невозможно, и быстродействие снова побежит в гору.
И еще вы о человеке говорите, как о каком-то волшебнике. Вы пишите: "Сможет ли ИИ написать книгу? Или картину?" - подразумевая, что писание книг и картин - это непостижимое чудо. Но ведь в голове писателя работают нейронные сети, которые позволяют ему писать книгу, а в голове художника - картину. Когда-нибудь ученые и инженеры смогут отследить работу этих сетей и создать алгоритмы, которые смогут писать книги и картины.
Кстати, уже сейчас есть ИИ, которое может написать картину. Я на выставке видел такого робота. У него видеокамера, он рисует карандашом то, что видит в камеру. Причем, он даже тени делал с помощью штриховки, и у него получались объемные предметы, хотя и просто карандашом. Конечно, это еще не Сальвадор Дали, но все-таки...
Я думаю, что сейчас основная проблема ИИ в том, что он не "живет" в мире людей. Когда сделают антропоморфного робота, который сможет существовать в обществе людей - вначале, может быть, как уборщик, или помощник, но он будет способен к обучению, к некому - вначале примитивному - пониманию речи, мимики и эмоций людей...
Тогда этот робот будет существовать среди людей и учиться у них. А параллельно инженеры будут оснащать таких роботов все большим количеством способностей. Пойдет эволюция роботов-помощников. И в какой-то момент они научатся хорошо понимать речь человека и говорить, выполнять различную работу руками, и постепенно достигнут уровня людей. Ну, а потом и обгонят человека.
Не стоит забывать, что как только один робот чему-то новому научился - это моментально становится достоянием всех роботов. Это может стать, как аппликации в смартфонах. Скажем, простой робот понимает простую речь, а кто-то написал аппликацию, позволяющую понимать иносказательную речь. А кто-то - аппликацию, позволяющую роботу сопоставлять речь и визуальные роботы. А еще кто-то аппликацию, позволяющую роботу мыть посуду, играть на пианино... Ведь всем этим вещам люди тоже учатся, не с рожденья же умеют. И один человек может только очень маленькую часть того, что могут люди целом.
В общем, фантазировать можно сколько угодно. Поглядим... -
xDDGx
xDDGx, я рад, что вам эта дискуссия интересна. Я вот еще что хотел бы упомянуть. Мозг человека "берет" совсем не быстродействием. Какое уж тут может быть быстродействие, когда скорость распространения нервного импульса не более 100 м/с, а скорость сигналов в компьютере 300,000,000 м/с?!
В мозгу 10 млрд нейронов, но даже когда вы напряженно решаете математическую задачу, "работают" над ней всего каких-нибудь несколько сотен, иногда тысяч - это известно из сканов мозга. Мозг силен не быстродействием. А тем, что он очень долго учится. Мозг взрослого человека двадцать (скажем) лет "строил" свою базу данных. Создавал связи в ней. Обучение - очень длительный процесс. Сколько часов на катке нужно провести, чтобы научиться как следует кататься? Сколько часов нужно провести над чтением книг и над разговорами, и над сочинением, чтобы стать писателем? А художником?
Людям мозг кажется чудом просто потому, что они не знают, как он работает, то есть не знают, каким образом решает ту или иную задачу. А когда что-то не знаешь, оно ВСЕГДА кажется чудом. Которое невозможно воспроизвести. Любая технология, если не знаешь, как работает, не отличима от чуда - как сказал один из ученых (не помню кто).
У меня, как у программиста, такое видение: для каждого "умения" в мозгу должен быть алгоритм. То есть "программа", которая это умение выполняет. У человека тысячи умений. Значит, у него в мозгу тысячи алгоритмов. Откуда они взялись? Частично заложены генетически, частично - обучением. (Не важно, что алгоритмы выполненны на нейронных сетях, а не в силиконе - главное, что они есть).
ИИ и роботы пока в зачаточном состоянии. Они тогда станут мощными и будут конкурировать с человеком, когда в ИИ (в роботах) будет очень много разных умений (то есть алгоритмов).
Все умения людей не заложены в одном человеке, а распределены во всем человечестве. Каждый человек владеет, может быть, десятитысячной долей всех умений.
То же самое должно произойти с роботами. Количество умений роботов будет расти, но каждый конкретный робот будет обладать небольшой частью умений.
(Кстати, это уже есть в мире компьютеров: количество программ и аппликаций в мире грандиозно, но на каждом конкретном компьютере их не больше нескольких десятков).
Разница между современными компьютерами и будущими антропоморфными роботами - в содержании умений. Все умения компьютеров сейчас - получать ввод от человека через клавиатуру, мышь и т.д., и выдавать человеку обратно вывод на экран, принтер... иногда запускать разные устройства.
Антропоморфному роботу нужны будут другие умения: ходить по улицам и в помещениях, открывать двери, брать в руки предметы, совершать действия над предметами, разговаривать с людьми...
Эти умения вполне достижимы при современных, а уж тем более, будущих технологиях.
Мои 5 копеек насчет мысли что нервная система слишком полезна чтобы исчезнуть. Возможно это произошло так давно, при такой архаичной экологии когда вероятность исчезновения была выше чем во все последующие эпохи. Вспоминается "эдиакарский сад" например, все что связано с этим словосочетанием, когда крупная жизнь была, но совсем другая, а вот в кембрий там уже все наши собрались :)
У меня одно мелкое замечание к фразе "но есть уникальная система межклеточной сигнализации, основанная на использовании разнообразных пептидов". Уникальности тут особой нет. Такая система межклеточной сигнализации очень похожа на кишечную эндокринную систему билатерий (секреторные клетки с сигнальными пептидами прямо в пищеварительном эпителии), а в меньшем масштабе подобный способ регуляции встречается и в других ресничных эпителиях, например, в трахее человека или ноге улитки. Стоит заметить, что среди пептидов трихоплакса есть четыре инсулина и один предположительный холецистокинин - типично кишечные гормоны.
-
А вот это - очень хорошее соображение!
Получается, гребневики отвалились на стадии, когда подвижная(?) колония уже научилась координировать внутреннее пищеварение, но еще не успела приобрести выделенную систему для интеграции восприятия и движения и их координации.
Надо смотреть, как там у губок с "пищеварительной координационной системой".-
Правильно, на той стадии, на которой находится трихоплакс.
И это соображение идеально ложится на нильсеновский сценарий.-
Я искал регуляторные пептиды у губки Amphimedon (тогда это была единственная губка с прочтенным геномом) и не нашел, а у трихоплакса вполне были (https://www.ncbi.nlm.nih.gov/pubmed/24747483). Надо будет повторить поиск, потому что появились геномы для еще нескольких губок, в том числе известковых, да и алгоритмы с 2015 года улучшились.
-
-
Видимо, мы все ждем от науки предсказательной функции. Из-за этого такой живой оффтоп от эволюции к ИИ и робототехнике.
-
-
Если бы ещё историки человеческие приняли эволюционный подход, взяв экономику с демографией на место экологии, а лингвистику на место биоинформатики - вообще бы прекрасно стало...
-
-
А работы Турчина Вы читали?
В.Ф. Турчина? Про метасистемный переход? Так как раз от этой концепции и отталкиваюсь! Только этапов там гораздо меньше получается и в несколько других местах.Лингвистику на место биоинформатики - это как?
Да также как и сейчас - через методы. Только смелее руководствуясь "утиным тестом". Также как физики правомочны называть кварк-глюонную плазму жидкостью, поскольку она наилучшим образом описывается через гидродинамику и т.д.
Смелее проводить аналогии между обретением ДНК и письменностью, параллельной неспецифичной репликацией у эукариот и книгопечатанием как раз используя принцип метасистемного перехода.
Тогда становится понятно, почему устная традиция столь же сравнительно невелика и изменчива как РНК-геномы, а до изобретения книгопечатания у народов как правило было лишь одно священное писание как одна хромосома у прокариот и всего несколько существенно меньших плазмид, а худлит массово появился только с появлением книгопечатания, как изобилие мусорной ДНК у эукариот...
И лингвисты - это получаются мемоинформатики, а филологи - системные меметики.
Вот такЪ.-
-
А какую предсказательную силу Вы ждёте, например, от таксономии, которая тут в статье разбирается? От этого она резко становится ненаучной что ли?
Сколько конкретных предсказаний было сделано по дарвиновской теории эволюции?
Сколько прогнозов погоды на месяц вперёд помогла сделать климатология?
Как теории гравитации и Ньютона и Эйнштейна помогут предсказать конкретные положения планет через сотни миллионов лет?
Нелинейность множества сложных природных процессов начиная с задачи трёх тел разом делает ненаучными и теории гравитации и эволюции и всю климатологию?
Поппер, к слову, сам сделал исключение для дарвиновской теории эволюции. И что - он это зря сделал?
На самом деле для дарвинизма (равно как и для кибернетики и для меметики) давно уже есть решение без исключений - но речь сейчас не о том, а о том, что всякий метод - даже такой всеобъемлющий как принцип фальсифицируемости - имеет свои правила и рамки применения.-
Предсказание от таксономии таково: когда мы продолжим изучать гребневиков, мы будем несколько чаще обнаруживать признаки, сходные с билатериями и стрекающими, нежели сходные с губками.
Можно ли подобное предсказание извлечь из идеи о том, что история человечества подобна эволюции жизни? Насколько надёжно будет такое предсказание?-
когда мы продолжим изучать гребневиков, мы будем несколько чаще обнаруживать признаки, сходные с билатериями и стрекающими, нежели сходные с губками.
"Верёвка есть вервие простое" - чрезвычайно высокой "предсказательной силы" утверждение, да-съ.Можно ли подобное предсказание извлечь из идеи о том, что история человечества подобна эволюции жизни? Насколько надёжно будет такое предсказание?
Мы будем обнаруживать литературу нерелигиозного назначения в гораздо бОльшей доле чем религиозного и устойчивое сохранение научного метода в полной мере только в обществах, освоивших книгопечатание, например. Контрпример и будет фальсифицирующим - подобно "кролику в докембрии". И т.д. и т.п...
Какие проблемы?-
В чём разница. Моё предсказание никто не стал бы произносить, пока считалось, что гребневики отделились раньше губок. Ваше же можно сделать без всякой генетики — из знаний об обществе (истории, социологии, экономики).
Понял ваши параллели для религии и худлита. Но для научного метода‑то у вас что? Где у эукариот «наука и техника» и чем она отличается от «религии»?
-
-
-
-
-
-
-
Но вот меня зацепило про нервную систему: рассчитывать найти продвинутую нервную систему в предковых организмах - это всё-равно что пытаться откопать квантовый (или нейронный) компьютер, произведённый в Древнем Шумере. :)
Т.о. отвергать сходу "протонейронные" элементы мне видится несостоятельным.
Главное у продвинутой нейронной системы на мой взгляд - это выполняемая ею функция. Функция передачи сигналов. Плюс контроль за исполнением. В принципе, те же протокариоты и археи элементарно передают между клетками различные сигналы: будь то производные орг.кислот, углеводов, пептидов, нукл.кислот и т.д. Это, однако, не означает, что все эти организмы обладают "нервной системой" или занимаются "нервной деятельностью". Скорее, что антропоцентризм человеков приписывает избыточные черты этой самой нервной системе. :)
С т.з. исполняемой ФУНКЦИИ вот эти все коммутации можно, наверное, проследить до какого-нибудь LUCA... И тут можно позвать в трэд камрада А.Маркова. :]
-
На самом деле - только до микоплазмы: https://www.ncbi.nlm.nih.gov/Complete_Genomes/SignalCensus.h
tml
Если у LUCA и были уже какие-то регуляторы - то их наследие какого-то судьбоносного значения не имеет: если погулять по приведённой базе, то взору откроется множество самых разных наборов генов самых разных регуляторов, вовсе не обязанных иметь единый корень - также как и породившие многие из них вирусы.
А А.В. по прокариотам не специалист - лучше тут вышеотметившегося Михаила Никитина спросите или меня. =]-
только до микоплазмы
Пофиг на микоплазму! Чего там со слизневиками?..А А.В. по прокариотам не специалист - лучше тут вышеотметившегося Михаила Никитина спросите или меня.
Окей. Давайте пригласим обоих! :)-
Пофиг на микоплазму!
Так вам же вроде как были интересны истоки клеточных сигнально-регуляторных систем. А чтобы их отследить, надо рассмотреть и сравнить наиболее простых из современных свободноживущих организмов, как это делалось при определении минимального и древнейшего наборов генов "домашнего хозяйства".Чего там со слизневиками?
Миксомицеты - эукариоты.
А миксококков и их ближайшей родни - сорангий - наиболее сложных прокариот - см. в приведённой таблице в группе Deltaproteobacteria.
У сорангий, кстати, в два раза больше генов чем у дрожжей.
P.S. Кстати среди миксококков - по большей части облигатных аэробов, есть и облигатные анаэробы - но их геном в два раза меньше. Задался вопросом: каков же максимальный размер генома бывает у облигатных анаэробов и у каких. Оказалось, что около 7 мегабаз и свыше 5 тысяч генов - причём у одних из наиболее (если не самых) многочисленных обитателей нашего кишечника - бактероидов (но далеко не самых распространенных видов - B. ovatus и B. cellulosilyticus)! Возможно это систематическая ошибка наблюдателя (геномы бактерий организма человека в базах данных сейчас составляют бОльшую долю), но тот факт, что у бактероидов число генов и геном больше чем у тех известных облигатно-анаэробных миксококков уже говорит в пользу этого предположения.
-
-
Но у меня по итогу сложилось впечатление, что тут уже происходит "подметание крошек" - и каких-то существенных перемен парадигмы далее ожидать уже не приходится. Возможно уже сейчас можно принимать представленный Нильсеном вариант за новый "канон".
Насколько это впечатление соответствует реальному положению вещей?
Нашёл небольшую опечатку:
>> Воротничок, которому воротничковые жгутиконосцы обязаны своим названием — это прозрачная воронка, окружающая жгутик и состоящая из множества отдельных микроворсинок, длинных тонких выростов с микрофиламентами внутри (см. рис. 1, А).
Тут явно неправильно указан номер рисунка, наверное, должно быть так:
<< (см. рис. 2, А)
>> Первые бесспорные остатки настоящих многоклеточных животных появляются в летописи не более чем за 13–15 миллионов лет до начала кембрия. (см. Кимберелла)
А что насчёт остальных вендобионтов? Могли ли они быть потомками губок? Или представлять собой отдельную ветвь/и, достигшую многоклеточности независимо? Или наиболее вероятно, что многоклеточность у животных возникла только один раз?





Рис. 1. Eocyathispongiа — эдиакарское ископаемое возрастом 600 млн лет (за 60 млн лет до кембрийского взрыва), предположительно являющееся древнейшей известной губкой. A — общий вид ископаемого, B1–B3 — отверстия для входа и выхода воды крупным планом. Иллюстрация из статьи: Z. Yin et al., 2015. Sponge grade body fossil with cellular resolution dating 60 Myr before the Cambrian