Как стать компактнее?

Древнейшие представители головоногих моллюсков, жившие в позднем кембрии (они относятся к подклассу Nautiloidea), имели прямые или слабо изогнутые наружные раковины. До тех пор пока они оставались небольшими животными (длиной несколько сантиметров), такая форма раковины им вполне подходила. Однако постепенно головоногие моллюски становились крупнее, а по мере увеличения размеров возникали сложности: длинная прямая раковина создавала проблемы при плавании и маневрировании.
Головоногие с наружной раковиной могут плавать двумя способами: либо при помощи воронки, либо используя специальную мембрану между щупальцами. Воронка — это мускулистая трубка, через которую моллюск с силой выбрасывает воду из мантийной полости. За счет возникающего импульса сам моллюск движется в противоположную сторону. Некоторые головоногие плавают, разводя в сторону соединенные мембраной щупальца и затем с силой сводят их вместе.
Как именно плавали древнейшие наутилоидеи, мы пока не знаем, но при любом из этих способов моллюск должен был двигаться раковиной вперед. И чтобы это движение было равномерным и управляемым, раковина должна быть относительно компактной. Для некоторых головоногих моллюсков, которые вели малоподвижный образ жизни и питались планктоном, неповоротливость не была проблемой, но для хищных наутилоидей, охотившихся на активную подвижную добычу, важно было самим быть подвижными и маневренными, а значит — компактными.
Задача
Древние головоногие изобрели два метода решения проблемы компактизации раковины. Что это за методы?
Подсказка 1
Первый метод становится достаточно очевидным, если посмотреть на фотографии современных наутилусов — единственных головоногих моллюсков, доживших до наших дней. А вот второй метод современные головоногие не используют, его стоит искать у вымерших наутилоидей.
Подсказка 2
Также оба этих метода компактизации раковины встречаются у современных и вымерших брюхоногих моллюсков (гастропод).
Решение
У головоногих моллюсков еще в раннем палеозое, практически одновременно в разных эволюционных линиях, возникло два способа компактизации раковин: сворачивание в плоскую спираль и отбрасывание заднего конца раковины (транкация).
Сворачивание раковины в спираль выгодно головоногим потому, что у свернутой раковины центр тяжести, находящийся в жилой камере (там, где жил сам моллюск), оказывается расположен на небольшом расстоянии под центром плавучести (который приходится на заполненный газом фрагмокон). Такая конструкция получается устойчивой и компактной и позволяет моллюскам активно плавать.
Самые крупные спирально-свернутые раковины наутилоидей (рис. 1) имеют диаметр порядка 60 см (такие жили в каменноугольном периоде, в том числе в Московской и Калужской областях). Эти же раковины, если бы они были прямыми, имели бы длину более трех метров. Естественно, моллюску гораздо удобнее плавать с компактной раковиной, чем с длинной «палкой», в которой, к тому же, центр тяжести оказывался бы впереди, в жилой камере, а центр плавучести — позади, во фрагмоконе.

Спирально-свернутые раковины появились у наутилоидей в раннем ордовике, около 480 миллионов лет назад. Первыми обладателями таких раковин стали представители отряда Tarphycerida. Хотя многие тарфицериды имели раковины с тесно соприкасающимися оборотами, очень похожие на раковины более поздних и эволюционно продвинутых наутилоидей, в этом отряде было немало форм, у которых обороты раковин соприкасались слабо или даже вовсе не соприкасались. О таких тарфицеридах можно сказать, что они решали задачу компактизации раковины частично: плавать с такой раковиной было, конечно, удобнее, чем с прямой, а вот быстро маневрировать вряд ли получалось.
Дело в том, что при сворачивании раковины в тугую спираль моллюск соединяется с раковиной не только внутри жилой камеры, но и «держится» специальной затылочной складкой мантии за предыдущий оборот раковины перед устьем. Для этого на раковинах современных наутилусов, к примеру, есть специальный черный слой, облегчающий сцепление тела с раковиной (см. рис. 1). Такое сцепление позволяет лучше распределять нагрузки, контролировать движение раковины и в целом способствует лучшей маневренности. Если же предыдущий оборот расположен далеко, подобное соединение невозможно. Но многих тарфицерид это не смущало: видимо, для их образа жизни маневренность не имела принципиального значения.

Рис. 2. Силурийские тарфицериды рода Discoceras. Фото из статьи: Š. Manda, V. Turek, 2018. Silurian tarphycerid Discoceras (Cephalopoda, Nautiloidea): systematics, embryonic development and paleoecology
Интересно, что у некоторых тарфицерид раковины были плотно свернутыми в детстве и в молодости, а вот на взрослой стадии развития они начинали расти прямо. Такая же форма раковин — спиральная на ранних оборотах с прямой взрослой жилой камерой — была характерна для всех представителей другого отряда — Lituitida, которых раньше считали разновидностью тарфицерид, но потом оказалось, что их предками были другие наутилоидеи — ортоцериды (Orthocerida). Палеонтологи предполагают, что у таких тарфицерид и у литуитид (Lituitida) в ходе онтогенеза менялся образ жизни: если в молодости они были активными и довольно маневренными животными, то в зрелом возрасте их подвижность снижалась, и они переходили к питанию какой-то не очень подвижной добычей.

Рис. 3. Раковина Lituites perfectus из отряда Lituitida. Фото с сайта commons.wikimedia.org
Раковины в плоскую спираль, кроме тарфицерид и литуитид, сворачивали представители отрядов Barrandeocerida (близкие родственники тарфицерид), Oncocerida и Nautilida (последние дожили до наших дней). Впрочем, пока еще не ясно, свернулись ли наутилиды сами или унаследовали свернутую раковину от своих предков, так как разные специалисты имеют разные взгляды на происхождение этого отряда.
А появление спиральных раковин в небольшом отряде Bactritida привело к появлению нового подкласса цефалопод — Ammonoidea. Аммоноидеи просуществовали более 300 миллионов лет, и хотя среди них периодически появлялись раковины самых разнообразных форм (см. картинку дня Гетероморфные аммониты), абсолютное большинство аммоноидей имели спирально-свернутые раковины. К аммоноидеям относятся и рекордсмены по размерам спирально-свернутых раковин — это позднемеловые аммониты Parapuzosia, диаметр которых мог достигать трех метров. Полная длина такой раковины, если бы она была прямой, превышала бы 10 метров — это больше, чем у самых крупных известных прямораковинных наутилоидей.

Рис. 4. Раковина аммонита Parapuzosia. Диаметр раковины 1,8 метра, но ее жилая камера сохранилась не полностью, и при жизни аммонита раковина могла иметь диаметр около 3 метров. Фото с сайта pinterest.com
Заканчивая рассказ про первый способ компактизации раковин, можно сказать, что сворачивание в спираль было распространено как минимум в пяти отрядах наутилоидей, один из которых дожил до наших дней и у целого подкласса аммоноидей. Независимо спиральная раковина (правда, внутренняя, расположенная внутри тела) возникла уже в кайнозое у спирулы (Spirula spirula) из подкласса колеоидей.
Второй путь компактизации раковин, которым пользовались цефалоподы, заключался в периодическом отбрасывании заднего конца раковины. До наших дней наутилоидеи с такой периодически укорачивающейся раковиной не дожили, но похожее явление встречается у некоторых современных улиток — к примеру, Rumina decollata. У улиток такое явление называется деколляцией, а в палеонтологической литературе в применении к цефалоподам распространен другой термин — транкация.
Чтобы отбрасывать задний конец раковины, наутилоидеям нужно было решить проблему с сифоном. Сифон — это специальная трубка, которая проходит через все камеры фрагмокона и отвечает за регуляцию плавучести раковины. Внутри сифона находятся крупные кровеносные сосуды. При транкации необходимо было как-то перекрывать эти сосуды и осуществлять заживление в месте отделения отбрасываемой части раковины. Как именно это происходило, можно только предполагать, но учитывая, что транкация применялась наутилоидеями на протяжении десятков миллионов лет, особых проблем у них не возникало.
Самые известные наутилоидеи, использовавшие транкацию для уменьшения размеров своей раковины, относились к отряду Ascocerida, который существовал в ордовикском периоде. В молодости аскоцериды мало отличались от своих предков ортоцерид: они имели прямые или слабо изогнутые раковины. Раковины, как и у всех цефалопод, были разделены на отдельные камеры перегородками-септами. Септы были одинаковыми, но в какой-то момент моллюск строил необычную — толстую и сильно изогнутую септу. Эта особенная перегородка («септа транкации») давила на стенки раковины и создавала напряжение, благодаря которому раковина в этом месте обламывалась.

Рис. 5. Схема развития наутилоидеи из отряда Ascocerida и фото взрослой раковины Ascoceras. Схема из статьи: B. Krüger et al., 2011. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules
Происходило ли это при обычном маневрировании или моллюск специально бросался из стороны в сторону или даже бился раковиной о какие-то препятствия (как иногда делают современные гастроподы, практикующие деколляцию раковин) — неизвестно, но после отбрасывания задней части раковина становилась заметно компактнее. Этот процесс повторялся несколько раз, но самое интересное начиналось, когда моллюск проходил стадию полового созревания. В это время аскоцериды строили уже не прямую, а яйцевидную жилую камеру с несколькими воздушными камерами на ее дорсальной (верхней) стороне. Задняя часть этой камеры перекрывалась особенно толстой «септой транкации», и после отбрасывания последнего прямого участка раковины моллюск оказывался в очень компактной, обтекаемой овально-яйцевидной камере, несомненно позволявшей ему активно плавать и маневрировать.

Рис. 6. Схема онтогенеза Sphooceras, показана регулярная транкация раковины. Схема из статьи V. Turek, Š. Manda, 2012. «An endocochleate experiment» in the Siluran straight-shelled cephalopod Sphooceras
Еще один пример транкации можно увидеть у ортоцеридного силурийского рода Sphooceras. Эти наутилоидеи, как и аскоцериды, периодически отбрасывали задний конец раковины; при этом они не только строили очень толстую септу, но и делали позади нее специальную «нашлепку», улучшающую обтекаемость раковины. Эта «нашлепка» — самая интересная часть раковины сфуцерасов: ее поверхность покрыта дополнительным слоем арагонита, на котором палеонтологи обнаружили следы прижизненной окраски — чередующихся светлых и темных полос (V. Turek, Š. Manda, 2012. «An endocochleate experiment» in the Siluran straight-shelled cephalopod Sphooceras). Эти дополнительные слои, несущие на себе окраску, должны были откладываться снаружи: сфуцерасы каким-то образом могли растянуть свою мантию (край которой как раз и отвечает за строительство раковины и за ее окраску) и натянуть ее на задний край раковины. То есть на некоторых стадиях роста раковины Sphooceras были полностью покрыты мантией, как у появившихся значительно позже внутреннераковинных головоногих подкласса Coleoidea (см. Силурийский «кальмар» Sphooceras — уникальный эксперимент природы). Кстати, у некоторых колеоидей, к примеру у каменноугольного рода Hematites, тоже происходила транкация задней части раковины, и на месте слома формировался кальцитовый выступ — ростр.

Рис. 7. Следы окраски на сформировавшемся после транкации конце раковины Sphooceras. Иллюстрация из статьи: V. Turek, Š. Manda, 2012. «An endocochleate experiment» in the Siluran straight-shelled cephalopod Sphooceras
Транкация также использовалась некоторыми силурийскими и девонскими представителями отряда Oncocerida. Эти наутилоидеи держали раковину вертикально (в отличие от аскоцерид и сфооцерасов, у которых положение раковин было горизонтальным), а при таком расположении размер раковин не так важен, как при горизонтальном плавании. К тому же их раковины были относительно небольшими и даже без транкации довольно компактными. Зачем им была нужна транкация? Возможно, отбрасывание части фрагмокона позволяло избавиться от излишка плавучести, ведь эти моллюски жили на дне, а не в толще воды. Также можно предположить, что уменьшение раковины за счет транкации позволяло им сделать ее менее заметной как для добычи, к которой они подкрадывались, так и для хищников, от которых им приходилось скрываться.
Однако, несмотря на распространенность транкации среди наутилоидей, со временем этот метод вышел из употребления. Последние наутилоидеи, использовавшие его, жили в начале перми. Это были представители семейств Scyphoceratidae и Dentoceratidae (см. Нижнепермские свернутые и согнутые наутилоидеи Южного Урала, систематическая принадлежность которых пока не ясна — то ли они относились к отряду Oncocerida, то ли к Nautilida. С их вымиранием история транкаций у наутилоидей закончилась.
В середине мелового периода транкация снова возникла у головоногих моллюсков, но уже не у наутилоидей, а у аммонитов рода Ptychoceras. Раковина этих необычных гетероморфных аммонитов (см. одноименную картинку дня) состояла из трех плотно прилегающих друг к другу прямых стволов, соединенных двумя коленообразными изгибами. Сначала такой аммонит рос прямо, затем резко менял направление строительства раковины и начинал расти в обратную сторону, строя более широкий и толстый ствол впритык к первому стволу. Затем птихоцерас снова резко изменял направление роста, строил жилую камеру и на этом формирование раковины завершалось. Конечная жилая камера перекрывала основание первого ствола, консервируя его и защищая от внешних воздействий. Но изучение раковин птихоцерасов, у которых исследователи удаляли третий ствол, показало, что у многих из них к моменту формирования третьего ствола начальная часть раковины оказывалась отломанной.
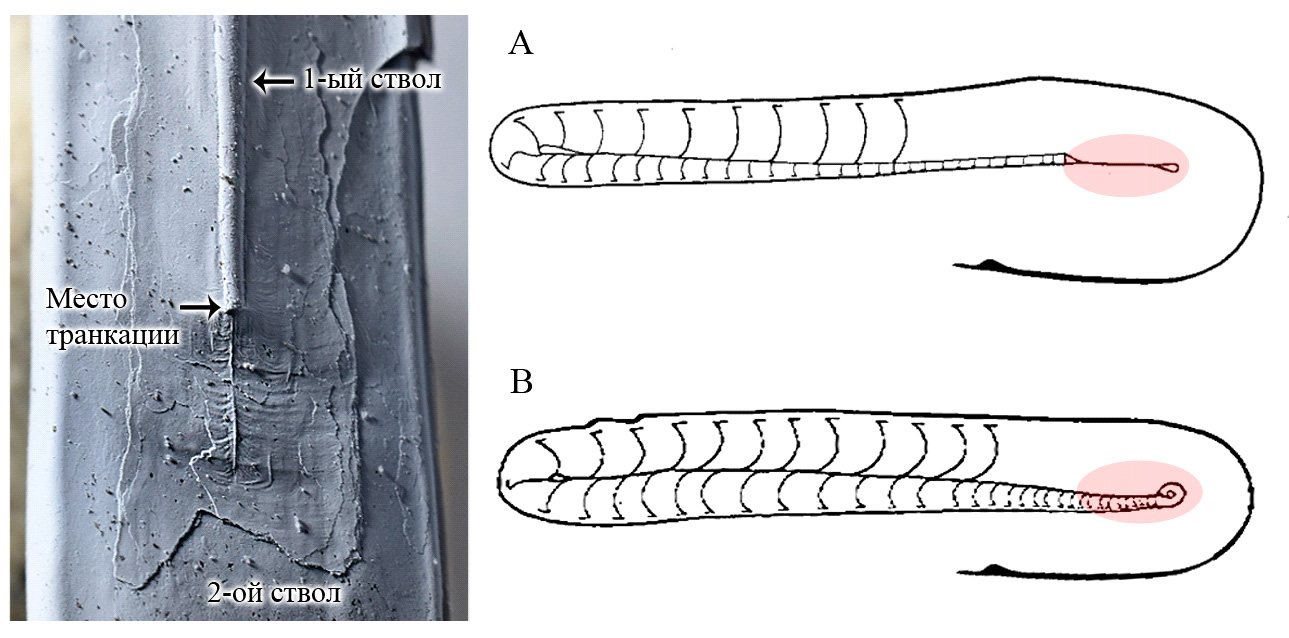
Рис. 8. Справа — схематические изображения раковин Ptychoceras. (А — транкация произошла, В — начальная часть раковины сохранилась). Слева — фото Ptychoceras с обломанным первым стволом. Обратите внимание на «шрам», идущий вниз от точки транкации, — он вызван деформацией периостракума из-за резкого исчезновения первого ствола во время строительства второго ствола раковины. Фото автора, схемы базируются на статье: L. Doguzhaeva, H. Mutvei, 1989. Ptychoceras — A heteromorphic lytoceratid with truncated shell and modified ultrastructure (Mollusca: Ammonoidea)
Это отбрасывание начальной части раковины тоже можно назвать транкацией. Но, в отличие от транкации у аскоцерид, онкоцерид или сфуцерасов, генетически запрограммированной и происходившей в строго определенных местах, после утолщенной септы, у птихоцерасов раковина обламывалась случайным образом — у кого-то ближе к началу, у кого-то дальше, а у некоторых сохранялась целой. Кроме того, от транкации раковина птихоцерасов не становилась компактнее, ведь второй и третий стволы всё равно были длиннее первого. Скорее всего, в данном случае транкация была случайным явлением, не имевшим никакого биологического смысла: просто аммонит, во время строительства мощного второго ствола, случайно обламывал себе тонкое основание раковины. Так как сечение раковины в этом месте не превышало 1 миллиметра, а толщина сифона и вовсе равнялась долям миллиметра, подобные травмы никак не влияли на жизнь моллюска и не приводили к затоплению раковины. Это тоже была транкация, но случайная, просто не приносившая моллюскам особого вреда.
Послесловие
Одновременно с головоногими моллюсками, становившимися всё более подвижными и компактными, жили и процветали формы с прямыми раковинами. Некоторые из них были крупными и очень крупными, длиной 5–6, а по некоторым данным — и до 9 метров. В таких отрядах, как Orthocerida, Oncocerida и Bactritida, от форм с прямой раковиной периодически ответвлялись эволюционные линии, шедшие по пути сворачивания или транкации раковин, но прямораковинные наутилоидеи продолжали существовать до самого конца триаса. Почему они обходились без компактизации раковин?
Часть этих наутилоидей были планктонофагами, медленно дрейфовавшими в толще воды и собиравшими планктон. Такие наутилоидеи, в первую очередь представители отряда Endocerida, держали раковины горизонтально, но к быстрым перемещениям способны не были (см. Гигантские наутилоидеи эндоцериды были мирными фильтраторами?, «Элементы», 02.08.2018). Их можно сравнить с современными китовыми акулами.
Относительно некрупные прямораковинные наутилоидеи могли охотиться на малоподвижную добычу у морского дна. Сложнее представить себе образ жизни наутилоидей с длинными прямыми раковинами, которые располагались в воде вертикально (о прижизненном положении раковины можно судить по ее окраске — при горизонтальном положении можно различить брюшную и спинную стороны, а при вертикальном окраска одинаковая со всех сторон). С узкой прямой раковиной длинной более метра, да еще и стоящей в воде вертикально, плавать можно разве что вверх-вниз, и то не очень быстро.

Рис. 9. Слева — раковина наутилоидеи Pomerantsoceras pollux с равномерной прижизненной окраской, свидетельствующей о вертикальном положении моллюска в воде. Фото из статьи: S. Manda and V. Turek, 2009. Minute Silurian oncocerid nautiloids with unusual colour patterns. Справа — современный глубоководный кальмар Magnapinna, вертикально висящий над дном и заснятый при помощи глубоководного спускаемого аппарата. Кадр из видео
Может быть понять образ жизни этих наутилоидей поможет недавно открытый глубоководный кальмар рода Magnapinna. Туловище этого кальмара тоже располагается в воде вертикально. Они зависают над дном, удерживая себя на одном месте при помощи огромных относительно длины тела плавников. При этом Magnapinna опускают на дно очень длинные (в 15 20 раз длиннее тела) и тонкие щупальца, которые, по-видимому, могут втягиваться или сильно сокращаться. Хватать такими нитевидными щупальцами более-менее крупную добычу невозможно, а вот собирать со дна и из придонного слоя воды планктон, который может прилипать к самим щупальцам или окружающей их слизи, очень даже можно.
Возможно, подобные длинные щупальца были и у различных ортоцерид, раковины которые вертикально висели в толще воды. В плавниках для поддержания плавучести ортоцериды и другие прямораковинные наутилоидеи не нуждались, у них был фрагмокон, и такой образ жизни для них выглядит вполне логичным. Правда, механизм питания Magnapinna всё еще остается не до конца понятным, да и проводить аналогии между глубоководными кальмарами и мелководными наутилоидеями далекого прошлого нужно с большой осторожностью. Но можно надеяться, что со временем палеонтологи смогут реконструировать образ жизни наутилоидей, не нуждавшихся в компактных раковинах.
-
>наутилусов — единственных головоногих моллюсков, доживших до наших дней.
Извините, но где-то здесь ошибка. -


Аммониты
-
 27.10.2021Ушки аммонитовАлександр Мироненко • Картинки дня
27.10.2021Ушки аммонитовАлександр Мироненко • Картинки дня
-
 28.01.2020Первый аммонитАлександр Мироненко • Картинки дня
28.01.2020Первый аммонитАлександр Мироненко • Картинки дня
-
 28.05.2019«Седло» аммонитаАлександр Мироненко • Картинки дня
28.05.2019«Седло» аммонитаАлександр Мироненко • Картинки дня
-
 17.05.2019Аммонит в янтареАлександр Храмов • Картинки дня
17.05.2019Аммонит в янтареАлександр Храмов • Картинки дня
-
 29.04.2019Аммониты из МиэринаАнтон Нелихов • Картинки дня
29.04.2019Аммониты из МиэринаАнтон Нелихов • Картинки дня
-
 17.12.2018Как стать компактнее?Александр Мироненко • Задачи
17.12.2018Как стать компактнее?Александр Мироненко • Задачи
-
 27.08.2018Мышцы аммонитовАлександр Мироненко • Задачи
27.08.2018Мышцы аммонитовАлександр Мироненко • Задачи
-
 06.07.2018Аммолитовый аммонитАлександр Мироненко • Картинки дня
06.07.2018Аммолитовый аммонитАлександр Мироненко • Картинки дня
-
 11.06.2018Аммониты-убежищаАлександр Мироненко • Картинки дня
11.06.2018Аммониты-убежищаАлександр Мироненко • Картинки дня
-
 15.03.2018Аптихи аммонитовАлександр Мироненко • Картинки дня
15.03.2018Аптихи аммонитовАлександр Мироненко • Картинки дня
Последние задачи









Рис. 1. Схематическое изображение спирально-свернутой раковины наутилоидеи и фото раковины современного Allonautilus из отряда Nautilida