Соматические клетки могут делиться без удвоения ДНК

Перед делением в соматических клетках эукариот удваиваются молекулы геномной ДНК, чтобы обе дочерние клетки получили равное количество генетического материала. Это — митоз. Альтернативный способ деления, при котором удвоения ДНК не происходит, — мейоз — задействован только при образовании половых клеток. Других способов деления эукариотических клеток известно не было. Но в недавней статье тайваньские ученые описали еще один способ. Они исследовали эмбрионы рыбок Danio rerio в период особенно интенсивного их роста, когда объем и площадь поверхности зародыша очень быстро увеличиваются. С помощью методики, которая позволяет отследить происхождение каждой клетки эпидермиса, авторы обнаружили, что часть клеток кожи у мальков данио делятся без репликации ДНК. Из-за этого на определенной стадии развития примерно половина всех клеток кожи у мальков имеет неполный геном. Несмотря на свою «дефектность» они выполняют важную функцию, помогая мальку быстро расти, а затем замещаются нормальными клетками. Скорее всего такой способ деления клеток встречается не только у мальков рыбок данио, а гораздо чаще.
Деление эукариотической клетки — это сложный и организованный процесс, который требует множества этапов контроля. Одна из фаз этого процесса — репликация ДНК. Она необходима, чтобы двум дочерним клеткам досталось одинаковое количество ДНК. Системы контроля над клеточным циклом четко следят за этим процессом. Пока нет сигнала о том, что репликация прошла полностью и успешно, клетка не может приступить к собственно делению. В норме исключение из этого «правила» лишь одно — второе деление мейоза, перед которым ДНК не удваивается и в результате образуются половые клетки с гаплоидным (одинарным) набором хромосом. Присутствие или отсутствие лишних хромосом связано с патологическими процессами, например онкологическим перерождением.
Благодаря клеточному делению многоклеточные организмы развиваются и регенерируют (то есть, по сути, восстанавливают утерянные клетки). В процессе эмбрионального развития деление клеток играет особенно важную роль, поскольку развивающемуся организму необходимо правильно заложить все ткани и органы. Естественно, важен и контроль за этим процессом.
Кожа взрослых позвоночных, их эмбрионов (начиная с определенной стадии развития) и личинок состоит из трех основных слоев. Каждый из них состоит из различных типов клеток и выполняет свою функцию. Нижний слой кожи (подкожно-жировая клетчатка) в основном состоит из жировой и соединительной ткани, которые составляют своего рода «подложку» кожи, связывающую ее с организмом. Также он играет роль теплоизолятора. Второй слой — дерма. Это соединительная ткань, которая придает коже упругость и эластичность. Сверху кожа покрыта многослойным эпидермисом, который преимущественно состоит из кератиноцитов (рис. 2). Компанию им составляют меланоциты (пигментные клетки) и клетки Лангерганса — иммунные клетки эпидермиса. Все вместе они обороняют организм от патогенов и других внешних факторов.
Эпидермис постоянно обновляется: его верхний слой, который состоит из уплощенных и, по сути, мертвых клеток-корнеоцитов (corneocyte), со временем слущивается. Однако он постоянно пополняется. Самый нижний слой эпидермиса (базальный слой) состоит из стволовых клеток. При их делении образуются кератиноциты, которые постепенно мигрируют наружу, со временем превращаясь в корнеоциты. У человека этот процесс занимает от 40 до 56 дней.
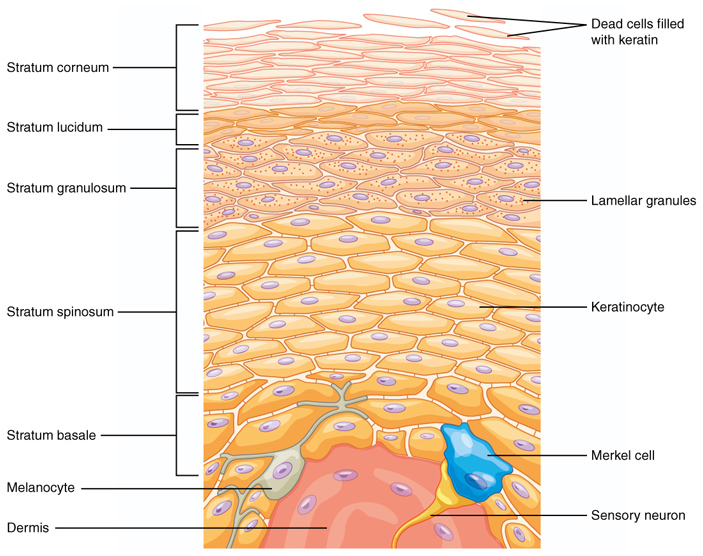
Рис. 2. Строение эпидермиса. Слева указаны слои эпидермиса. Роговой слой (stratum corneum) состоит из отмерших клеток-корнеоцитов, наполненных кератином. Блестящий слой (stratum lucidum) заполнен мертвыми клетками, в которых уже не осталось ни ядра, ни органелл. Зернистый слой (stratum granulosum) содержит клетки, заполненные ламеллярными гранулами (lamellar granules), и обеспечивает водонепроницаемость эпидермиса. Шиповатый слой состоит из нескольких слоев кератиноцитов (keratinocyte), в нем содержатся клетки Лангерганса (не показаны). Базальный слой — делящиеся клетки, которые обеспечивают регенерацию эпителия. В этом же слое расположены меланоциты (melanocyte), механорецепторы (клетки Меркеля) и другие типы клеток. Базальный слой снизу граничит с дермой (Dermis). Рисунок с сайта en.wikipedia.org
Все эти подробности о строении и функционировании кожи были раскрыты с помощью клеточных и молекулярных методов, изучения срезов эпидермиса и различных способов окрашивания. Однако мечтой многих ученых была возможность запечатлеть все эти процессы в живом организме. В 2007 году была разработана методика Brainbow, позволяющая проследить, как связаны между собой индивидуальные нейроны (см. картинку дня Разноцветные нейроны). Каждый из них окрашивается сочетанием флуоресцентных белков, и из-за того, что в каждой клетке разные белки имеют немного разные концентрации, получаются разные цвета. Окрашенные таким образом клетки можно рассмотреть с помощью микроскопирования, в том числе и прижизненного.
С помощью похожей методики можно проследить судьбу отдельных клеток, а также определить, как часто они делятся и куда потом мигрируют их потомки. В 2016 году исследователь из Университета Дьюка Чэнь-Хуэй Чэнь (Chen-Hui Chen) разработал метод, с помощью которого можно пронаблюдать, как восстанавливается кожа после повреждения у рыбок Danio rerio (C.-H. Chen et al., 2016. Multicolor cell barcoding technology for long-term surveillance of epithelial regeneration in zebrafish). Рыбки данио оказались отличным инструментом для изучения этих процессов. Их эпидермис устроен примерно так же, как и у людей, за исключением того, что покрыт не слущивающимися корнеоцитами, а поверхностными эпителиальными клетками (впрочем, они тоже со временем слущиваются) и слоем слизи. Нижний слой эпидермиса рыбок также представляет собой прослойку стволовых клеток.
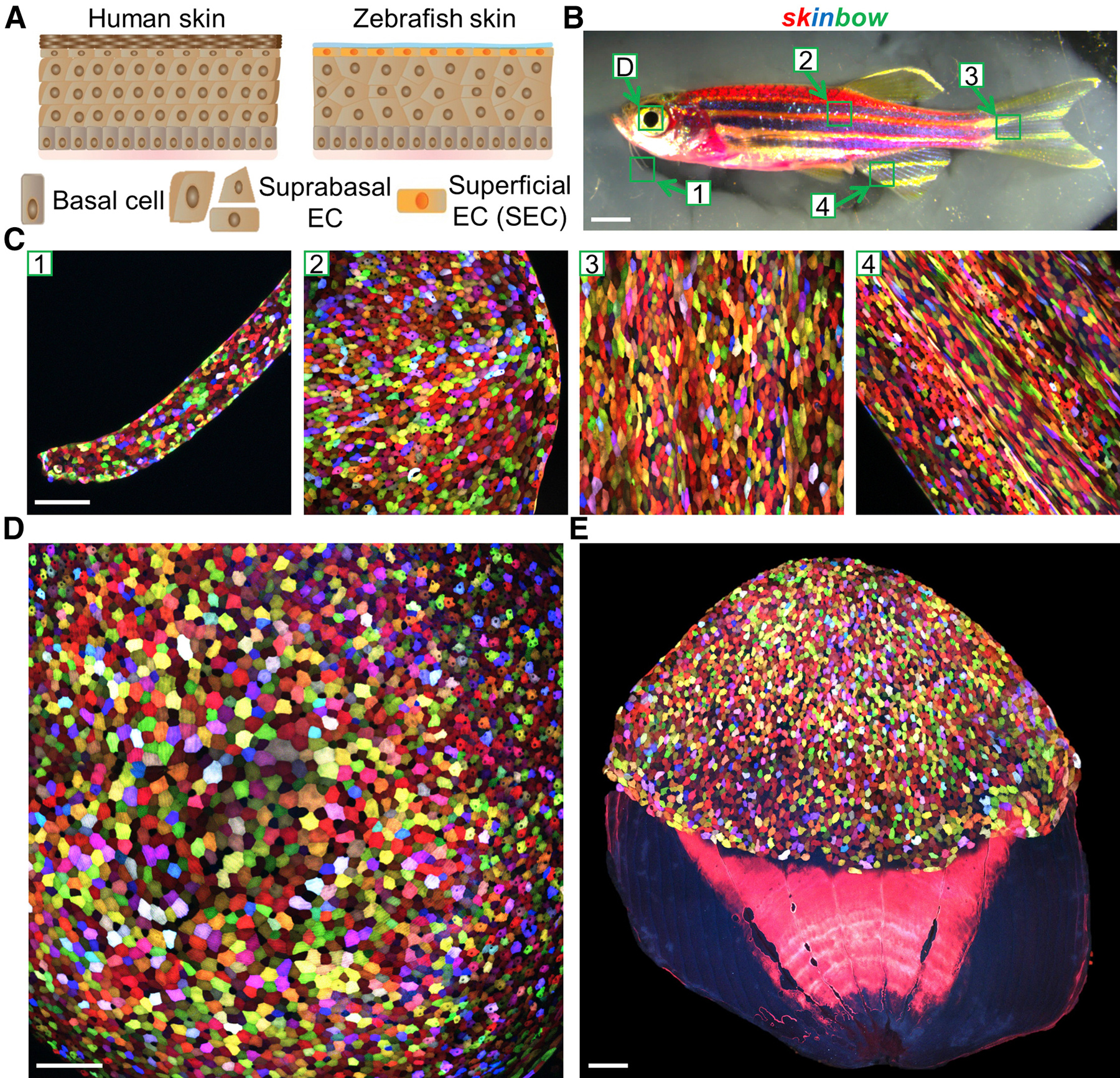
Рис. 3. Методика skinbow, позволяющая следить за клетками кожи Danio rerio. A — схема строения кожи человека (human skin) и рыбок данио (Zebrafish skin). Базальные клетки (basal cells) расположены снизу, супрабазальные клетки эпителия (Suprabasal EC (epitelial cells)) находятся над ними, а на поверхности расположены поверхностные клетки эпителия (эпидермиса) (Superficial EC). Сверху кожа Danio rerio покрыта прозрачной слизью (голубой слой). B — фотография рыбы, «в которой» работает система skinbow. Цифрами обозначены участки тела, соответствующие микрофотографиям на изображении C. Длина масштабного отрезка — 2 мм. C — микрофотографии участков тела рыбки. Каждая клетка окрашена разным сочетанием флуоресцентных белков. Длина масштабного отрезка — 100 мкм. D, E — многоцветное кодирование клеток эпителия роговицы (D) и чешуи (E). Длина масштабного отрезка — 100 мкм. Рисунок из статьи С.-H. Chen et al., 2016. Multicolor Cell Barcoding Technology for Long-Term Surveillance of Epithelial Regeneration in Zebrafish
Но то, как происходит развитие кожи, не менее интересно, чем то, как она функционирует. Поэтому в новом исследовании команда под руководством доктора Чэня решила применить схожий метод, чтобы изучить развитие кожи Danio rerio. Работа началась с создания генетических конструкций, которые бы позволили пронаблюдать за отдельными клетками.
Ученые создали три линии рыбок. Две линии несли в себе ген Cre-рекомбиназы (см. Cre recombinase). Этот белок узнает специфические сайты ДНК (они называются LoxP) и может их либо вырезать, либо переворачивать на 180 градусов. Чтобы запустить работу этого гена, нужно активировать промоторы — последовательности ДНК, которые запускают синтез мРНК. В генетических конструкциях были два таких промотора: Tet3G — запускается добавлением антибиотика доксициклина и TRE — запускается добавлением тамоксифена (рис. 4). Такая система надежно контролирует запуск синтеза Cre-рекомбиназы. Перед всей этой конструкцией находится либо промотор убиквитина (универсального белка домашнего хозяйства; эту линию рыбок авторы назвали Tg(SEC:iCre#1)), либо промотор гена KRT19, который кодирует белок, экспрессирующийся в поверхностных клетках эпидермиса в районе туловища рыбы (линия Tg(SEC:iCre#2)). Интересно, что, независимо от использования первой или второй линии с рекомбиназой в качестве одной из родительских, окрашивались только поверхностные клетки эндотелия. Однако авторы не объясняют это и преимущественно используют рыб, происходящих от линии Tg(SEC:iCre#2).
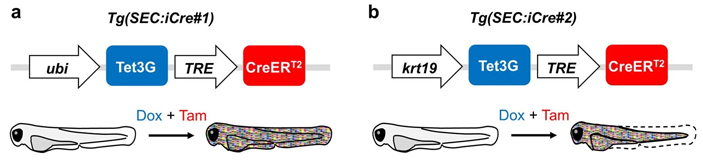
Рис. 4. Генетические конструкции в родительских линиях рыбок данио. a — в линии Tg(SEC:iCre#1) экспрессия конструкции, содержащей Cre-рекомбиназу, контролировалась промотором убиквитина (ubi). b — в линии Tg(SEC:iCre#2) экспрессия конструкции, содержащей Cre-рекомбиназу, контролировалась промотором гена KRT19. При скрещивании рыбок этих линий и рыбок, у которых в геном были встроены гены, кодирующие флуоресцентные белки и loxP-сайты, образовывалось гибридное потомство. В этих гибридах при добавлении доксициклина и тамоксифена экспрессировалась Cre-рекомбиназа, из-за чего происходили изменения в уровне экспрессии флуоресцентных белков. В результате мембраны поверхностных клеток эпидермиса приобретали один из 60 оттенков. Рисунок из дополнительных материалов к обсуждаемой статье в Nature
Третья линия рыб также несла генетическую конструкцию, в которой присутствовали loxP-сайты — последовательности ДНК, с которыми связывается Cre-рекомбиназа (см. рис. 1, a). Эти сайты — своего рода дорожные знаки, указывающие на то, что с этим фрагментом ДНК должна сделать Cre-рекомбиназа: вырезать его или же разрезать и поменять направление. Еще в этой конструкции присутствовал ген гистона H2B, дополненного последовательностью синего флуоресцентного белка BFP2, а также разделенные loxP-сайтами гены флуоресцентных белков RFP (красный цвет), YFP (желтый цвет) и CFP (синий цвет). Гены RFP, YFP и CFP содержали добавку в виде последовательности PALM, которая нужна была для того, чтобы белки распределялись именно по мембране клетки, а не оставались в ее цитоплазме.
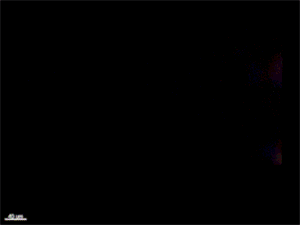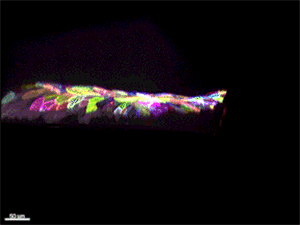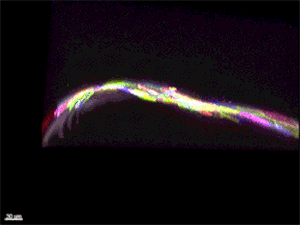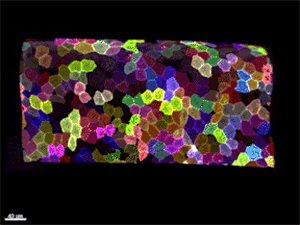
Рис. 5. Многоцветное кодирование поверхностных клеток эпидермиса у живого малька Danio rerio. 3D-реконструкция, полученная с помощью конфокального микроскопа. Видео из обсуждаемой статьи в Nature
При скрещивании линий данио образуются гибриды, которые содержат и одну из конструкций с Cre-рекомбиназой, и флуоресцентные белки. На 4-й день развития гибридных рыбок добавляли тамоксифен и доксициклин на 2–3 часа. В результате начинался синтез мРНК, а затем белка Cre-рекомбиназы, которая начинала вырезать или менять порядок генов флуоресцентных белков из второй конструкции. Из-за этого каждая клетка получала свою комбинацию флуоресцентных сигналов (таким образом в обсуждаемой работе было получено 60 различных оттенков), которая сохранялась у ее потомков. Разработанный метод назвали palmskin.
Чтобы изучить развитие кожи мальков Danio rerio, авторы осматривали их с помощью микроскопа на восьмой и десятый день после оплодотворения (соответственно, на 4-й и 6-й день после индуцирования «окраски» путем добавления тамоксифена и доксициклина). За эти два дня мальки на 10% увеличивали длину тела и на 22% — площадь ее поверхности. Число поверхностных клеток эпидермиса при этом росло на 21%. По распределению цветов было видно, что некоторые клетки могут быть потомками одной клетки. И именно это озадачило исследователей больше всего. Дело в том, что поверхностные клетки эпидермиса не делятся — они теряют способность делиться уже на второй день после оплодотворения. Делиться способны лишь базальные клетки.

Рис. 6. Делящиеся поверхностные клетки эпидермиса малька данио. Снимки делались каждые 15 минут в течение 12 часов. Стрелками отмечены фиолетовая и светло-зеленая делящиеся клетки. Видео из обсуждаемой статьи в Nature
Однако анализ микрофотографий подтвердил, что делятся именно поверхностные клетки эпидермиса, а не подлежащие клетки. Они делятся вдоль и со временем становятся более круглыми. Как оказалось, клетки могут делиться максимум два раза, что приводит к образованию четырех дочерних клеток.
Следуют ли эти клетки обычным правилам деления? В частности, удваивают ли они свою ДНК?
Чтобы ответить на эти вопросы, ученые использовали добавку EdU (5-Ethynyl-2'-deoxyuridine) — аналог тимидина, который встраивается в реплицирующуюся ДНК и позволяет определить ее присутствие с помощью микроскопии. Авторы работали с линиями рыб, у которых флуоресцентные белки экспрессируются либо только в поверхностных, либо только в базальных клетках эпидермиса. Как и предполагалось, последние делились, а поверхностные клетки не подавали никаких признаков типичного деления с предварительным удвоением ДНК (рис. 7).
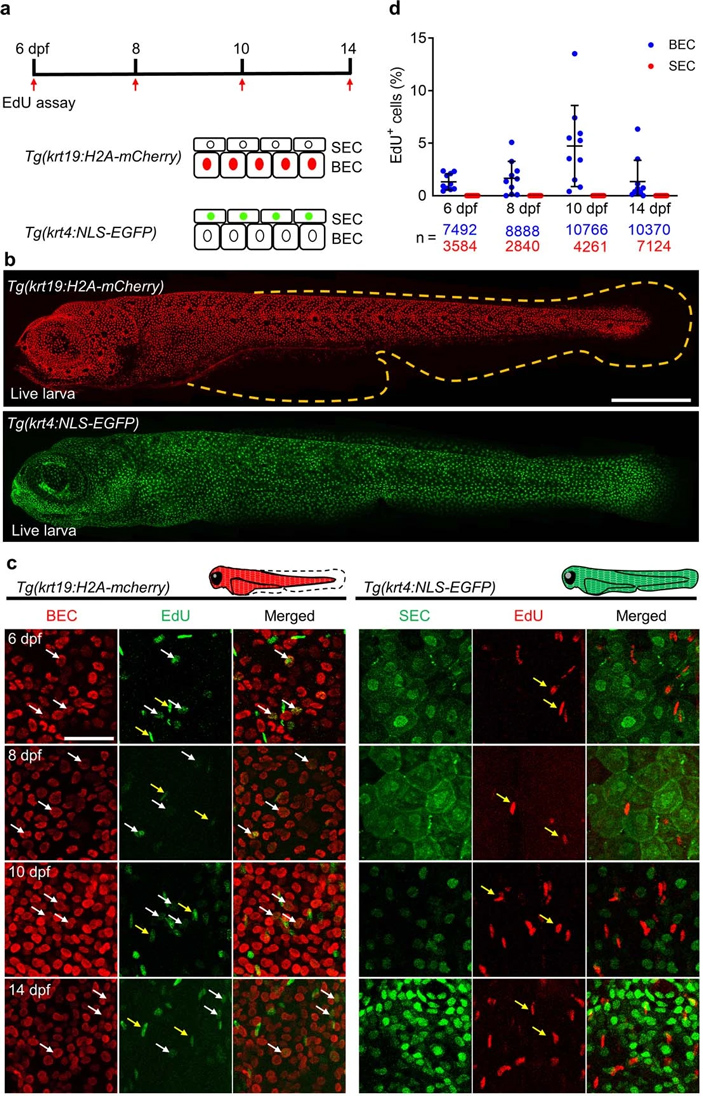
Рис. 7. В поверхностных клетках эпителия отсутствует репликация ДНК. a — схема эксперимента. EdU добавляли на 6, 8, 10 и 14 день после оплодотворения мальков двух линий Danio rerio: в линии Tg(krt19:H2A-mCherry) красным цветом выделены все клетки базального слоя эпидермиса, в линии Tg(krt4:NLS-EGFP) зеленым цветом выделены все поверхностные клетки эпидермиса. b — мальки этих линий. Желтым пунктиром показаны контуры малька. Длина масштабного отрезка — 500 мкм. c — результаты детекции EdU (и реплицирующейся ДНК) в базальных (слева) и поверхностных (справа) клетках эпидермиса. Белыми стрелками обозначены клетки, в которых происходит репликация. Желтыми стрелками — реплицирующиеся клетки, не относящиеся к поверхностным или базальным клеткам. В популяции поверхностных клеток эпидермиса не обнаружено репликации ДНК. D — количественные результаты. Синими точками обозначены базальные клетки (BEC), красными — поверхностные. По вертикальной оси — процент клеток с сигналом от EdU. Длина масштабного отрезка — 50 мкм. Иллюстрация из дополнительных материалов к обсуждаемой статье в Nature
Объяснение налюдаемой картины может быть только одно: клетки делятся, но в результате образуются клетки с неполным геномом. Ядра полученных клеток имели менее интенсивное окрашивание ДНК и ассоциированных с ними гистонов. Более детальный анализ с помощью проточной цитометрии показал, что большинство клеток действительно имеют количество ДНК, соответствующее неполному диплоидному набору, но со временем эта ситуация меняется из-за слущивания эпидермиса. Поверхностные клетки эпидермиса у взрослой рыбы имеют типичный диплоидный набор хромосом. Но в мальке Danio rerio поверхностные клетки напоминали аномальные раковые клетки: у них образовывались микроядра, хромосомы распределялись не по двум полюсам клетки, как при обычном делении, а по трем, были и другие аномалии.
Почему же в организме присутствуют такие клетки? Вероятно, это имеет свою практическую пользу. Пик деления поверхностных клеток эпителия и пик увеличения поверхности тела малька совпадают: оба пика наблюдаются на 14-й день после оплодотворения. Поэтому авторы предположили, что функция такого необычного деления заключается в том, чтобы получить достаточное количество эпидермиса для быстрого роста и при этом сохранить ресурсы организма. При таком делении общий размер клеток не увеличивался — потомки клеток использовали плазматическую мембрану и цитоплазму материнской. Тем не менее одно деление увеличивало поверхность, которую покрывает клетка, на 26%, а два — на 59%, а сами клетки уплощались и со временем слущивались.
Это предположение проверили в интересном эксперименте. Размер рыбы можно регулировать с помощью изменения плотности выращивания: чем больше рыб живет в ограниченном объеме воды, тем меньшего размера они вырастают. Авторы показали, что рыбы, жившие в менее стесненных условиях, были больше, а их поверхностные клетки эпидермиса делились чаще.
Но самый интересный вопрос — что же все-таки регулирует такое деление? Так как деление явно связано с запросом на увеличение поверхности, авторы предположили, что в этом процессе участвуют активируемые натяжением ионные каналы PIEZO. За открытие этих белков Ардем Патапутян получил в прошлом году Нобелевскую премию по физиологии и медицине (см. Нобелевская премия по физиологии и медицине — 2021, «Элементы», 08.10.2021). Рецепторы PIEZO можно инактивировать, добавляя ионы гадолиния, либо, наоборот, активировать с помощью агониста Yoda1. Оказалось, что поверхностные клетки эпидермиса соответственно уменьшают или увеличивают частоту своего деления. Наконец, авторы протестировали эффект удаления гена piezo-1 у Danio rerio: без него поверхностные клетки эпидермиса значительно снижали частоту своего деления. Таким образом, их деление зависит от поверхностного натяжения, которое возникает во время роста рыбы.
Вредит ли такое «ненормальное» деление всему организму? Видимо, нет — даже несмотря на то, что возникающие нарушения (например, деление клетки, которая предварительно не удвоила количество ДНК) обычно приводят к апоптозу, как это происходит во взрослых тканях. Это еще одна из загадок новооткрытого способа деления клеток. Обычно возникающие аномалии четко регулируются различными системами защиты: от систем контроля клеточного цикла до апоптоза и иммунного ответа, которые удаляют аномальные клетки. Как поверхностным клеткам эпидермиса удается избежать этого контроля — неизвестно.
Отмечу, что это не единственные ткани организма, которые в норме справляются с отсутствием полного набора хромосом. Например, в половине клеток печени отсутствуют те или иные хромосомы, однако же эти клетки способны делиться. Они преодолевают системы контроля ради того, чтобы, например, справляться с повреждением печеночной ткани (A. W. Duncan et al., 2012. Aneuploidy as a mechanism for stress-induced liver adaptation).
Кроме того, пока непонятно, присущ ли новооткрытый способ деления клеток только рыбам или же этот процесс является эволюционно консервативным и встречается у других групп организмов, в то числе и млекопитающих. Дальнейшие исследования помогут ответить на этот вопрос. Однако уже точно ясно, что в школьные учебники следует добавить еще один вид деления клеток.
Источник: Keat Ying Chan, Ching-Cher Sanders Yan, Hsiao-Yuh Roan, Shao-Chun Hsu, Tzu-Lun Tseng, Chung-Der Hsiao, Chao-Ping Hsu & Chen-Hui Chen. Skin cells undergo asynthetic fission to expand body surfaces in zebrafish // Nature. 2022. DOI: 10.1038/s41586-022-04641-0.
Екатерина Грачева
-
>>Кроме того, пока непонятно, присущ ли новооткрытый способ деления клеток только рыбам
а он вообще присущ кому-нибудь, кроме их ГМО? Может, это артефакт генетической модификации вылез.-
Ссылка на работу 2012 года в том самом абзаце, что вы процитировали, говорит, что это возможно. Вы специально только первую часть процитировали, чтобы ваше предположение вызывало меньше сомнений, или вы действительно читаете настолько избирательно? Хотя сама по себе мысль логичная, не могу спорить
-
>>Вы специально только первую часть процитировали, чтобы ваше предположение вызывало меньше сомнений, или вы действительно читаете настолько избирательно?
я читаю довольно-таки подряд, надеюсь, как и вы. В процитированном мной фрагменте говорится о _новооткрытом способе_. Способ, описанный для гепатоцитов 10 лет назад, вряд ли можно назвать новооткрытым, не так ли? Значит новооткрытием здесь названо деление именно поверхностного эпителия, а гепатоциты упомянуты постольку-поскольку.
Да и вообще, с биологической точки зрения, какой смысл организму надробить тяп-ляп клеток поверхностного слоя, когда внизу есть базальный эпителий? Если он растет нормально - он ограничивает скорость роста площади поверхности, тяп-ляп поверхностные клетки просто складками будут ложиться. Если базальный тоже ускоряется - его потомки и так будут становятся поверхностными в увеличенном количестве.
-
-


-
-
Пол заметки посвящено подробностям прицепления к клеткам флуоресцирующих молекул, а вы хлобысь - условная раскраска.
https://ru.wikipedia.org/wiki/Флуоресцентная_микроскопия-
Прошу прощения, там на 1е условная раскраска. А на рисунке 6 просто разделение уже существующей выросшей до максимального размера клетки со всем содержимым на 2 куска. В ней (и потомках) вероятно, почти прекратилось образование флуоресцентных молекул, недаром же отмечено, что новые клетки, полученные таким путём, не растут.
-
-
-
>>К тому же окраску придают уже наработанные белки, которые при делении никуда не деваются.
при делении они, понятно, никуда не деваются. Но как насчет обычного метаболизма? Индукцию цвета сделали на 4 день, снимать начали на 8 день - допустим четыре дня цвет накапливается. В допматериалах статьи есть фото клеток, разделившихся в первый день наблюдения и прослеженных пять дней - они одинаковые по интенсивности до конца.
Почему конструкция перестала работать и увеличивать интенсивность там, где она осталась?
Ладно, допустим конструкция включается на пару часов, всю клетку закрашивает и замолкает навсегда
Почему почти нет черных клеток? Поступившие нормальным путем от базального эпителия уже после индукции должны быть черными. Из разделившихся до индукции половина должна быть черными. Может, они не делились до индукции? Может, они от этой индукции и начали делиться?
Последние новости












Рис. 1. Многоцветное кодирование поверхностных клеток эпидермиса рыбки Danio rerio. a — схема генетической конструкции, обеспечивающей разнообразие разных цветов. Конструкция состоит из гена гистона H2B, дополненного последовательностью синего флуоресцентного белка BFP2 (окрашивает ядра клеток в синий цвет), и генов флуоресцентных белков RFP (красный цвет), YFP (желтый цвет), CFP (синий цвет) с последовательностью palm (она позволяет экспрессировать белки на мембране клетки). Гены разделены Lox-последовательностями (показаны треугольниками), распознаваемыми Cre-рекомбиназой, которая вырезает или меняет порядок флуоресцентных белков (см. Cre-Lox recombination). В итоге в разных клетках разные цветовые белки синтезируются с разной интенсивностью, что придает клеткам один из 60 оттенков. b — поперечный срез малька Danio rerio на 8 день после оплодотворения. Экспрессия гена BFP2 происходит только в поверхностных клетках эпидермиса (зеленые). Красным цветом обозначен слой базальных клеток (окрашенный антителами к белку p63, который синтезируется в базальных клетках). Ядра клеток обозначены белым цветом (DAPI). Длина масштабного отрезка — 20 мкм. c — малек Danio rerio (8 дней после оплодотворения), экспрессирующий систему Palmskin только в поверхностных клетках эпителия; длина масштабного отрезка — 100 мкм. d — увеличенный участок кожи малька. e — с помощью окрашивания системой Palmskin можно увидеть границы клеток и их поверхностные микрогребешки; длина масштабного отрезка — 20 мкм. f — многообразие оттенков поверхностных клеток эпителия. Рисунок из обсуждаемой статьи в Nature