Чем важнее ген, тем реже он мутирует

Изучение большого массива данных по мутагенезу у модельного растения Arabidopsis thaliana показало, что в разных участках генома мутации возникают с разной частотой. Темп мутагенеза можно предсказать по эпигенетическим признакам, таким как уровень метилирования ДНК, открытость хроматина и модификации гистонов. Распределение этих эпигенетических меток, в свою очередь, зависит от функциональной нагрузки участков ДНК. В результате получается, что частота возникновения новых мутаций связана обратной зависимостью с функциональной важностью данного участка генома и с силой действующего на него очищающего отбора. Иначе говоря, в наиболее важных участках новые мутации не только активнее вычищаются отбором, но и реже возникают. В частности, темп мутагенеза ниже внутри генов по сравнению с внешними (нетранскрибируемыми) областями и в жизненно важных генах, работающих постоянно, по сравнению с теми, что используются лишь эпизодически (например, включаются в ответ на какие-то внешние стимулы). По-видимому, в ходе эволюции у некоторых организмов под действием отбора развились молекулярные механизмы, снижающие частоту мутаций в наиболее важных частях генома. В основе их работы лежит привлечение ферментов репарации и других факторов, защищающих ДНК от повреждений, к определенным эпигенетическим меткам. Исследование показало, что неслучайный мутагенез играет важную роль в эволюции геномов. Некоторые характерные особенности молекулярной эволюции, которые обычно объясняют действием отбора (например, ускоренное накопление различий в менее важных участках генома), на самом деле во многом объясняются неслучайным мутагенезом. Который, впрочем, сам является результатом эволюции под действием отбора.
1. Эволюция скорости мутирования
Постулат о случайности мутаций входит во все учебники эволюции и вряд ли его когда-нибудь оттуда вычеркнут. Хотя уже давно известно, что он не абсолютен и нуждается во множестве оговорок и уточнений, число которых неуклонно растет по мере развития науки.
Мутации действительно случайны в том смысле, что у живых существ нет механизмов, позволяющих «рассчитать», какая мутация будет для них полезна в данных условиях, и аккуратно внести именно эту мутацию в свой геном. При этом у всех без исключения организмов геномы мутируют, фенотипические эффекты мутаций никем заранее не просчитываются, а точно предсказать, какие именно мутации возникнут в данной хромосоме при следующей репликации, невозможно. В этом отношении мутации случайны, во многих других аспектах — нет. Многие организмы сумели в ходе эволюции выработать механизмы, так или иначе регулирующие и оптимизирующие мутационный процесс (см., например: Мутагенез в лимфоцитах — результат целенаправленного изменения ДНК и последующей «неточной починки», «Элементы», 03.09.2007). Несколько показательных примеров рассмотрено в моей книге «Рождение сложности», где одна из глав целиком посвящена этой теме (она так и называется — «Управляемые мутации»).
Нельзя назвать случайной и такую важнейшую характеристику мутационного процесса, как его скорость. Темп мутагенеза зависит, в частности, от работы ферментов, осуществляющих репликацию и репарацию ДНК, и он эволюционирует под действием отбора. Поскольку большая часть не нейтральных (влияющих на приспособленность) мутаций вредны, отбор, как правило, способствует минимизации темпа мутагенеза. Хотя и тут есть исключения: например, некоторым вирусам высокий темп мутагенеза необходим для повседневного выживания, так что мутации, снижающие его ниже допустимого предела, отбраковываются отбором (см.: Вирусы-мутанты помогают друг другу в борьбе за выживание, «Элементы», 14.12.2005).
Но даже те организмы, которым не нужно постоянно мутировать, чтобы выживать здесь и сейчас, все равно не могут свести темп мутагенеза к нулю, потому что отбору на его снижение противостоят другие эволюционные силы. Главными из них считаются две. Во-первых, сверхточные системы репликации и репарации, скорее всего, будут слишком затратными: громоздкими, потребляющими много энергии и т. д. Поэтому в какой-то момент «стоимость» дальнейшего совершенствования этих систем перестаёт окупаться выигрышем от дальнейшего снижения темпа мутагенеза (иначе говоря, отбор на замедление мутагенеза уравновешивается отбором на упрощение и удешевление молекулярных систем, следящих за сохранностью генетического материала).
Вторая причина связана с генетическим дрейфом. Полезные мутации (в том числе мутации, снижающие темп мутагенеза), могут быть поддержаны отбором, только если их полезность превышает определенный порог, зависящий от эффективной численности популяции (Ne, Effective population size). Чтобы отбор помог мутации распространиться, желательно, чтобы ее полезный эффект (величина, на которую мутация увеличивает эффективность размножения) был больше, чем 4/Ne (и уж никак не меньше, чем 1/Ne). В противном случае мутация будет находиться во власти дрейфа, а не отбора, то есть вести себя не как полезная, а как нейтральная, и шансов зафиксироваться (достичь 100-процентной частоты) у нее будет очень мало. Чем ниже темп мутагенеза, тем меньше причиняемый им вред, и тем слабее полезный эффект от мутаций, замедляющих мутагенез еще сильнее. Поэтому в какой-то момент такие мутации перестают поддерживаться отбором. Предполагается, что именно такой уровень мутагенеза, соответствующий равновесию между отбором и дрейфом, и является конечным итогом эволюции скорости мутагенеза у многих организмов (M. Lynch et al., 2016. Genetic drift, selection and the evolution of the mutation rate).
2. Возможен ли избирательный контроль темпа мутагенеза?
В последние годы стали появляться данные, указывающие на то, что скорость мутирования может сильно различаться в разных участках генома и в разных генах, и что эти различия могут быть связаны, во-первых, с активностью (уровнем экспрессии) гена, во-вторых — с эпигенетическими метками (которые во многом определяют эту активность), такими как метилирование и ацетилирование лизинов в гистоне H3 или, скажем, «открытость хроматина», определяемая частотой расположения нуклеосом (см., например: F. Supek, B. Lehner, 2017. Clustered Mutation Signatures Reveal that Error-Prone DNA Repair Targets Mutations to Active Genes; X. Chen et al., 2012. Nucleosomes Suppress Spontaneous Mutations Base-Specifically in Eukaryotes).
В связи с этим возникает вопрос: не могли ли какие-нибудь организмы выработать в ходе эволюции специальные адаптации, позволяющие снизить темп мутирования не во всем геноме сразу, а избирательно — например, только в самых важных генах, случайные мутации в которых наиболее опасны?
Теоретически такое вроде бы возможно при определенных условиях, хоть и не совсем понятно, как часто эти условия выполняются (I. Martincorena, N. M. Luscombe, 2013. Non-random mutation: The evolution of targeted hypermutation and hypomutation). Из двух ограничений, упомянутых выше, в случае избирательного замедления мутагенеза одно слабеет, а другое усиливается. Слабеет ограничение, связанное с затратностью: с особой тщательностью защищать от мутаций отдельные гены, по идее, должно быть проще и дешевле, чем весь геном. Усиливается ограничение, связанное с дрейфом. Ведь в отдельном гене мутации возникают в тысячи раз реже, чем в геноме в целом (просто потому, что ген в тысячи раз короче генома). А значит, от дополнительного снижения темпа мутагенеза только в этом одном гене приспособленность увеличится совсем ненамного, даже если это очень важный ген.
Хорошо бы, конечно, придумать механизм, снижающий темп мутагенеза во всех важных генах сразу. Этому дрейф должен мешать не так сильно, поскольку суммарная длина важных генов достаточно велика. Но возможен ли такой механизм? Существует ли он у каких-нибудь реальных организмов?
3. Сбор данных по мутагенезу у Arabidopsis thaliana
Статья группы исследователей из США, Германии, Франции и Швеции, опубликованная 12 января в журнале Nature, показывает, что как минимум один хорошо изученный организм — модельное растение резуховидка Таля (Arabidopsis thaliana) — обладает искомой способностью. Почти наверняка арабидопсис в этом не уникален, но не будем забегать вперед: строгие доказательства пока получены только для этого вида.
Исследователи собрали большой массив данных по мутагенезу у Arabidopsis. Одна из главных трудностей тут в том, что эффекты мутагенеза нужно тщательно отделять от эффектов отбора. Например, если мы просто отсеквенируем геномы 1000 растений, сравним их между собой и выявим все полиморфизмы (вариации нуклеотидной последовательности), то получим картину, отражающую не мутагенез в чистом виде, а совместное действие мутагенеза и отбора. Например, мы не увидим значительной части вредных мутаций, которые постоянно возникают, но отбор старательно вычищает их из генофонда.
Чтобы свести влияние отбора к минимуму, исследователи использовали несколько подходов. Один из них показан на рис. 1 (слева вверху). От каждого растения брали по одному случайно выбранному семечку, выращивали из него растение-потомка, от него снова брали случайное семечко, и так далее. Спустя 24 поколения геномы потомков секвенировали и сравнивали с геномом исходного растения. При таком подходе отбор хоть и не устраняется вовсе, но радикально ослабляется: в итоговую выборку не попадут (будут отбракованы) только летальные мутации, а также те, что ведут к полному бесплодию. Другой подход связан с выявлением соматических мутаций путем секвенирования разных клеток одного и того же растения. В этом случае можно выловить даже мутации, которые на уровне целого растения были бы несовместимы с жизнью или размножением.
Полученный список мутаций затем проверялся различными статистическими тестами, чтобы убедиться, что он отражает именно процесс мутагенеза (то есть не был сильно искажен отбором). Результаты двух таких тестов показаны на рис. 1 справа вверху. Использовались два показателя: соотношение несинонимичных (значимых) и синонимичных замен и отношение числа замен, ведущих к появлению преждевременного стоп-кодона, к числу синонимичных замен. Идея в том, что синонимичные замены обычно нейтральны, отбор их не выбраковывает. Значимые замены, напротив, часто оказываются вредными, преждевременные стоп-кодоны — тем более. Биологи даже в самых буйных фантазиях не могут вообразить, что возможны механизмы, позволяющие клетке избирательно снизить частоту значимых замен или мутаций, создающих стоп-кодоны. Или вообще хоть как-то отличить такие замены от синонимичных на уровне ДНК. Таких механизмов почти наверняка не существует. Поэтому в ходе мутагенеза должно появляться намного больше значимых замен и преждевременных стоп-кодонов, чем их потом останется в генофонде популяции, находящейся под действием отбора.
Исходя из этой логики, выявленные мутации белок-кодирующих участков генома (на графиках они подписаны «De novo») сравнивали, во-первых, с природным генетическим разнообразием A. thaliana (из проекта «1001 геном», 1001G), во-вторых, с теоретически ожидаемым спектром вновь возникающих мутаций (Null).
Оказалось, что оба показателя значимо выше у выявленных мутаций (De novo), чем в природных популяциях арабидопсиса (1001G), и лишь чуть-чуть (незначимо) ниже, чем у теоретически ожидаемых мутаций. Следовательно, отбор не сильно повлиял на собранные данные, а значит, их можно использовать для изучения закономерностей мутагенеза. К такому же выводу привели и другие проверки.
4. Скорость мутагенеза можно предсказать по эпигенетическим меткам
Убедившись в адекватности исходных данных, авторы приступили к их анализу. Они сопоставили частоту возникновения мутаций в разных участках генома с эпигенетическими характеристиками, такими как открытость хроматина, метилирование ДНК и модификации гистонов. Оказалось, что многие эпигенетические признаки значимо коррелируют с темпом мутагенеза (рис. 1, нижние графики). Например, в участках с высоким уровнем метилирования четвертого лизина в гистоне H3 (H3K4Me1) темп мутагенеза ниже, чем в среднем по геному, а в участках с высоким уровнем ацетилирования девятого лизина того же гистона (H3K9ac) он, наоборот, повышен.
По данным об эпигенетических метках можно, как выяснилось, довольно точно предсказать темп мутагенеза для разных участков генома. Например, на рис. 2a показан предсказанный по эпигеномным данным усредненный темп мутагенеза для участков, прилегающих к точкам начала (TSS) и окончания (TTS) транскрипции (Upstream, Downstream — нетранскрибируемые участки перед геном и после него, Gene body — «тело гена»). Видно, что темп мутагенеза повышен в нетранскрибируемых участках, особенно в непосредственной близости от границ гена (вероятно, потому, что там много участков открытого хроматина, к которым должны прикрепляться всякие регуляторные белки). Причем предсказанная картинка (рис. 2, а) действительно очень похожа на реально наблюдаемую (рис. 2, b).
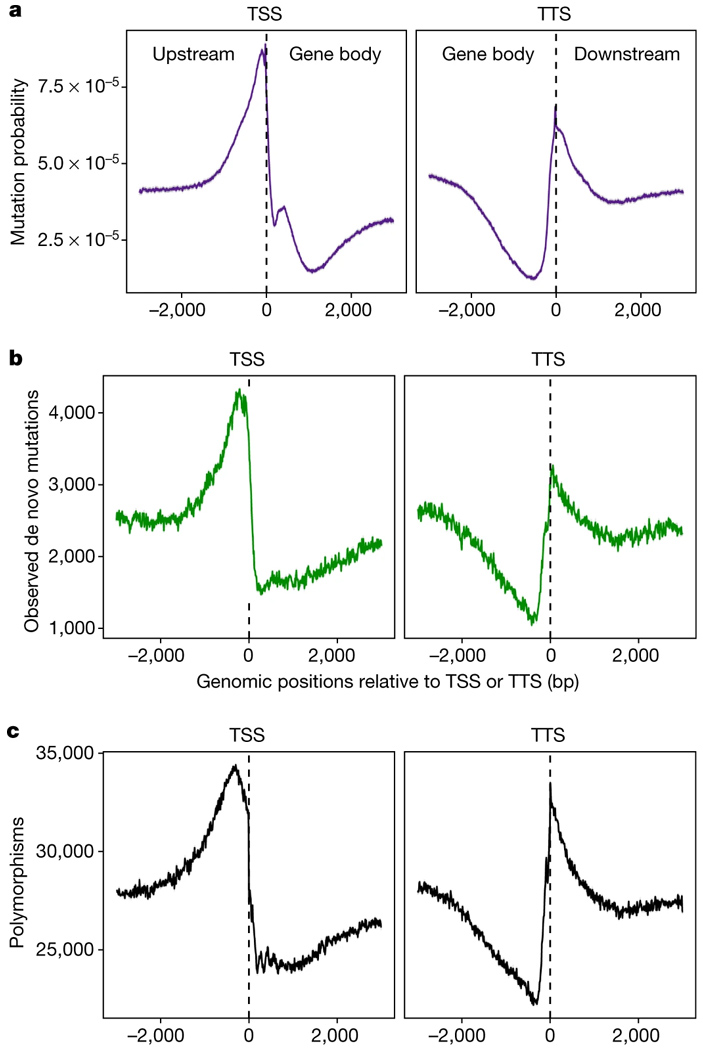
Рис. 2. Темп мутагенеза в окрестностях точек начала (TSS) и окончания (TTS) транскрипции (усредненные данные по всем генам). a — темп мутагенеза, предсказанный по эпигеномным данным (с учетом корреляций, показанных на рис. 1); b — реальные данные по мутагенезу; c — генетический полиморфизм в природных популяциях (на основе 1135 геномов). Рисунок из обсуждаемой статьи в Nature
Более того, распределение мутаций по окрестностям TSS и TTS довольно точно совпадает с распределением генетической вариабельности (полиморфизмов) в природных популяциях (рис. 2, c). Из этого (а также из ряда дополнительных проверочных тестов) следует, что наблюдаемое распределение полиморфизмов в большей степени определяется мутагенезом, чем отбором. Вывод довольно-таки сенсационный, ведь до сих пор «по умолчанию» было принято объяснять подобные распределения именно отбором.
Авторы также обнаружили, что разные части генов мутируют с разной скоростью, что эти различия тоже можно предсказать по эпигеному, и что примерно так же распределены и природные полиморфизмы. В частности, из всех экзонов, входящих в состав гена, чаще всего мутируют первый и последний, и в них же чаще всего встречаются полиморфизмы в природе. Скорость мутирования экзонов связана отрицательной зависимостью с длиной нетранслируемых областей гена, а также с числом и длиной интронов. Чем длиннее нетранслируемые области и интроны, тем реже мутируют белок-кодирующие участки гена.
5. Чем слабее очищающий отбор, тем сильнее мутагенез
Означает ли всё это, что очищающий отбор, которым традиционно объясняли распределение генетических различий по геномам (где различий меньше, там предполагали более сильный отбор), в действительности тут вовсе ни при чем? На первый взгляд вроде бы да, на самом деле нет.
С одной стороны, исследование показало, что скорость накопления генетических различий (например, между особями одного вида или между близкими видами) в тех или иных участках генома определяется мутагенезом в большей степени, чем отбором. Мутагенез, в свою очередь, регулируется эпигенетическими метками.
С другой стороны, авторы показали, что самая низкая скорость мутагенеза (и соответствующие ей эпигенетические метки) приурочены как раз к тем участкам генома, которые находятся под наиболее сильным очищающим отбором. И наоборот, в тех участках, где очищающий отбор ослаблен, эпигенетические характеристики способствуют ускоренному мутагенезу.
В частности, выяснилось, что гены с самыми низкими темпами мутагенеза — это в основном гены с самыми консервативными (постоянными, мало меняющимися в ходе эволюции) функциями, такими, например, как трансляция. Подобные гены, как правило, работают постоянно в большинстве клеток и тканей. Они необходимы для повседневного, базового выживания (поэтому их еще называют «генами домашнего хозяйства). Такие гены обычно находятся под сильнейшим очищающим отбором. Иначе говоря, мутации в них чаще всего оказываются вредными и отбраковываются. Эти гены давно оптимизированы, менять их незачем, и они действительно меняются в ходе эволюции очень медленно.
Напротив, у генов, функции которых связаны с реакцией организма на переменчивые условия среды, темп мутагенеза оказался повышенным. Очищающий отбор слабее действует на такие гены, а мутации в них имеют чуть больше шансов оказаться полезными. Поэтому такие гены в ходе эволюции меняются быстрее.
Выяснилось также, что темп мутагенеза понижен в жизненно важных генах (отключение которых несовместимо с жизнью), а также в генах, экспрессирующихся во многих клетках и тканях по сравнению с генами с узкими профилями экспрессии. В обоих случаях различия в темпах мутагенеза коррелируют с эпигенетическими характеристиками.
О силе действия очищающего отбора на тот или иной ген и об имеющихся ограничениях на его эволюционные изменения (evolutionary constraint) можно судить по целому ряду количественных показателей, таких как наследственная и средовая изменчивость уровня экспрессии гена или соотношение значимых и синонимичных различий при сравнении последовательностей данного гена у разных особей одного вида или у близких видов (Dn/Ds). Авторы выполнили много таких тестов и всё сошлось: чем важнее ген, чем фундаментальнее его функция и чем сильнее эволюционные ограничения на его изменения, тем ниже темп мутирования гена — как эмпирически измеренный, так и предсказанный по эпигенетическим признакам.
6. Что всё это значит?
Полученные результаты показывают, что у Arabidopsis (а скорее всего и у многих других организмов, хотя это еще предстоит доказать) в ходе эволюции, по-видимому, выработались специальные механизмы, позволяющие снизить скорость мутирования наиболее важных участков генома — тех участков, мутации в которых чаще всего оказываются вредными. В результате среди вновь возникающих мутаций существенно снижается доля вредных, а общее негативное влияние мутагенеза на приспособленность уменьшается.
Предполагаемый принцип работы этого механизма в самых общих чертах показан на рис. 3. Его детали еще только предстоит расшифровать.
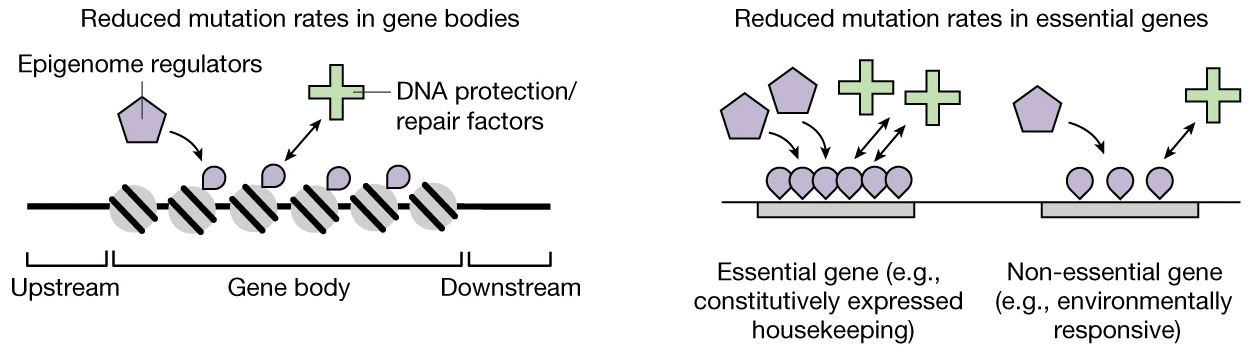
Рис. 3. «Концептуальные диаграммы», отражающие авторскую интерпретацию полученных результатов. Рисунок из обсуждаемой статьи в Nature
Исследование показало, что по своим эпигенетическим характеристикам гены отличаются от межгенных участков, а важные гены — от менее важных. Причем различия эти не абы какие, а закономерные, предсказуемые и похожие в обоих случаях. Например, уровень метилирования четвертого лизина гистона H3 (H3K4Me1) повышен как в «теле гена» (Gene body на рис. 3) по сравнению с прилегающими нетранскрибируемыми участками (Upstream, Downstream), так и в жизненно важных (Essential) генах по сравнению с менее важными (Non-essential). Почему эпигенетический признак H3K4Me1 распределен именно так, а не иначе, и какие молекулярные механизмы (Epigenome regulators на рис. 3) ответственны за такое его распределение — отдельный вопрос, пока еще очень далекий от разрешения. Тем не менее, мы знаем, что по данному признаку важные участки генома можно отличить от менее важных.
Кроме того, известно, что в клетке существуют специализированные молекулярные системы, отвечающие за репарацию и защиту ДНК от повреждений (DNA protection/repair factors на рис. 3).
Авторы предполагают, что в ходе эволюции у «факторов защиты и репарации ДНК» (или у каких-то регуляторных систем, контролирующих их деятельность) выработалось полезное свойство, состоящее в том, что эти факторы активнее, чаще или тщательнее работают с участками ДНК, обладающими определенными эпигенетическими характеристиками — например, повышенным уровнем H3K4Me1. Это свойство — не что иное, как эволюционная адаптация, направленная на оптимизацию мутационного процесса и уменьшение приносимого им вреда.
Важным практическим следствием является необходимость пересмотра механизмов эволюции нуклеотидных последовательностей. Многое из того, что приписывалось действию отбора, на самом деле, по-видимому, является результатом не совсем случайного мутагенеза. Многое, но, разумеется, не всё. Например, пониженные значения Dn/Ds у жизненно важных генов нельзя объяснить мутагенезом: это трудноподделываемая роспись очищающего отбора. Кроме того, не стоит забывать, что сам характер обнаруженной авторами «неслучайности» мутагенеза наверняка является результатом эволюции под действием отбора.
Источник: J. Grey Monroe, Thanvi Srikant, Pablo Carbonell-Bejerano, Claude Becker, Mariele Lensink, Moises Exposito-Alonso, Marie Klein, Julia Hildebrandt, Manuela Neumann, Daniel Kliebenstein, Mao-Lun Weng, Eric Imbert, Jon Ågren, Matthew T. Rutter, Charles B. Fenster & Detlef Weigel. Mutation bias reflects natural selection in Arabidopsis thaliana // Nature. 2022. DOI: 10.1038/s41586-021-04269-6
-
По-видимому, в ходе эволюции У НЕКОТОРЫХ организмов под действием отбора развились молекулярные механизмы, снижающие частоту мутаций в наиболее важных частях генома.
Этими некоторыми организмами окажутся не менее чем все эукариоты, сдаётся мне. Но скромность и осторожность, скромность и осторожность.-
Да, я тоже склонен считать, что у сложно организованных организмов аналогично сложно организован и геном и, в том числе, содержащиеся в нём механизмы самомодификации.
https://elementy.ru/novosti_nauki/433922/#forum
В качестве «пересмотра механизмов эволюции» я в своих (бумажно-компьютерных) модельках пытаюсь отказаться от «фундаментальности» процесса саморепликации, заменив его фундаментальным процессом «производства». Т.е., организм (в моём случае - условная клетка) может преобразовывать имеющиеся в себе ресурсы в новые структуры (содержит программы продюсирования новых структур), при этом эти структуры могут получаться и похожие на исходный организм (а отбором эта похожесть может увеличиваться или уменьшаться, программы производства могут становиться точнее или вариативнее).
Т.е., эволюцию репликаторов я методологически заменяю на эволюцию продюсеров, а случайность мутагенеза переезжает в параметры алгоритмов производства. Таким образом в моей модельке половое разделение, например, или эпигенетика становятся выводимыми свойствами, а не изначально заложенными (т.е., могут возникать и исчезать в процессе моделирования). И даже может моделироваться динамика «культурной» эволюции (эволюции расширенного фенотипа) - организмы могут продюсировать «неактивные» структуры, которые могут быть использованы потомками (т.е., с точки зрения передачи информации культура - это такая «внешняя эпигенетика»). Если эти структуры будут использоваться лишь в качестве энергии, можно говорить и об «экономической» эволюции. А если структуру выстраивают несколько индивидов, можно говорить о половом процессе (или о коллективном творчестве :)). И даже способов самокопирования может возникнуть куча вариантов, которые будут конкурировать между собой (конкурс так называемых квайнов - самопроизводящих программ).
Проблема практического использования таких гиперуниверсальных эволюционных моделей (в сравнении с обучением нейросетей, например, или с классическими генетическими алгоритмами) - в очень низкой скорости сходимости, потому начинать моделирование лучше с готовых программ репликации, закодированных вручную в генах. Вообще в гены можно затолкать кучу разных готовых алгоритмов (эвристик относительно решаемой задачи моделирования, закодированной в факторах отбора), которые организм уже и будет модифицировать под давлением отбора (или отбрасывать за ненадобностью лишние эвристики). И, вероятно, эвристика «копирования участка генома» - это самая фундаментальная эвристика (алгоритмический инвариант) и самый первый оптимум многофакторной эволюционной оптимизации, в который когда-то и «провалился» генетический бульон, породив Луку - победителя конкурса квайнов и родителя всех нас (гордых носителей структуры ДНК и всех программ-энзимов, её обслуживающих) :). Но для «полной» работоспособности эволюции («свободности» параметров модели :)) важно, чтобы эти программы копирования были закодированы «в терминах» продюсеров, выражены набором «ферментивных операций», подверженных давлению отбора, а не были фиксированным свойством модели (репликаторов).
Итого базовым формализмом для построения эволюционирующих систем у меня получился «химический автомат» (примерно эквивалентный биоклетке) - сущность, объединяющая в себе набор правил преобразования ресурсов вида «а + б -> в + г» (где буквы - это указатели на исходные и продюсируемые ресурсы и их количество, могут быть нулями, а также правила должны укладываться в ограничения постоянства массы/энергии/других интересующих в решаемой задаче инвариантов) и набор этих ресурсов (таблица со строками «вид ресурса + количество»). Некоторые из этих ресурсов являются «физическими абстракциями», к примеру, оболочкой продюсируемой клетки (т.е., в ходе исполнения правил при достижении наполненности какого-то условного ресурса он обнуляется, а рядом с автоматом возникает ещё один автомат, пустой, а при заполнении других условных ресурсов они «обмениваются» на операцию записи гена в эту заготовку, кодирующего или правило преобразования, или вид ресурса; какие-то ресурсы могут обмениваться на движение автомата в каком-то моделируемом пространстве; какие-то перемещают ресурсы извне во внутрь клетки; и т.п.).
Эта модель эволюционной динамики получается очень абстрактной, и в ней можно кодировать любой (интересующий исследователя) уровень организации эволюционирующей системы без необходимости кодировать низлежащий. К примеру для исследования социальной динамики нам не нужно выращивать из этих клеток многоклеточные организмы, эквивалентные по сложности человеческому (на это нам не хватит жизни вселенной), достаточно подобрать нужный набор абстрактных ресурсов и их интерпретацию и считать автомат целым индивидом или даже социальной группой. И то, что в масштабе биоклетки можно считать продюсированием пузырька с сигнальными молекулами, например, в масштабе человеческого социума можно считать речевым актом и конструированием сообщения :).
(Готовых результатов, чтобы прям уже показать и удивить разумным поведением говорящих роботов, или биологических примеров, как-то напрямую обосновывающих описанную теорию, у меня нет. Но, может, кому-то покажется интересным покритиковать её и в таком виде или провести аналогии с другими уже существующими теориями. Или хотя бы намекнуть на проблемы повествования и мутность формулировок или отсутствие новизны :).)-
Кстати, получается, что я в каком-то смысле расширяю докинзову теорию эгоистичного гена (ограничиваю её область применимости и пристраиваю к ней «недостающую» часть), ибо ген - это монетка экономики эволюции репликаторов (что работает только на относительно коротких исторических отрезках, на которых изменениями скорости мутагенеза и структуры генома можно пренебрегать), а монеткой экономики эволюции продюсеров является квазистабильный химический/трофический/метаболический/сигнальный цикл (замкнутая последовательность преобразований ресурсов). Эгоистичный уроборос :).
Эгоистичный ген - это такая ньютоновская механика генетики (эволюция генов в статичном «эфире» генофонда), а эгоистичный цикл - уже релятивистская механика экологии (коэволюция изменений факторов отбора и ответа популяции на них) :).-
Мне это напоминает модель "химической эволюции".
Смысл в том, чтобы в теории эволюции репродукцию заменить на случайное возникновение, если "организмы" достаточно просты. Если при этом они влияют на частоту этого случайного возникновения, то возникает естественный отбор. А если в процессе случайного возникновения могут возникнуть и другие похожие "организмы", то эта система может заменить Дарвиновскую эволюцию и привести к росту сложности.
"Организмы" здесь скорее органические вещества, чем даже первый РНК репликатор.
Ссылки по теме:
https://youtu.be/mRzxTzKIsp8
https://youtu.be/oDvzbBRiNlA
https://en.wikipedia.org/wiki/Autocatalytic_set-
Да, я уже прямо в этих комментариях (ниже) догуглился до хемотонов Тибора Ганти :). Моя модель только чуть-чуть абстрактнее, оторвана от химии, у меня преобразование "любых" ресурсов, а так же я планирую "бесшовно" переходить между абстракциями ресурсов (при переходе из одной экологической ниши в другую, с более крупным масштабом самоорганизации). Концептуально и у меня, и в остальных подобных теориях осуществляется, по сути, приложение методов кибернетики к автокаталитической химии / биологии / любым другим эволюционирующим системам.
Только это не замена дарвиновской эволюции, а, скорее, её подробности или обобщение, смещение фокуса с эгоистичного гена на эгоистичный цикл, катализируемый этим геном, а в моём случае ещё и абстрагирование от этой химии до топологии таких циклов, "упакованных" в преобразование абстрактных ресурсов. Т.е., «(а) + (б) -> (в) + (г)» это ещё абстрактнее, чем "(а) + (б) + (кат) -> (а-1) + (ц) + (кат)", но очень может быть, что для симуляции именно химических процессов будет удобнее ("быстрее сходимее") пользоваться более "жирной" каталитической нотацией (хотя математически они взаимовыразимы).
-
-
-
-
-
Как и способность к их управляемой селекции помимо грубого естественного отбора в виде дополнительного контура к нему - отбору половому: https://elementy.ru/novosti_nauki/432164/Drozhzhi_pomnyat_o_
nerazdelennoy_lyubvi
И главное тут - не проронить, что _вообще любое_ творчество в своей основе имеет дарвиновский механизм, отличаясь лишь числом контуров отбора - ибо скромность и осторожность, скромность и осторожность! Чтобы никаких попыток к "телеологии" и никакого "соляриса" из биосферы не лепить, упаси Лаплас! Правда тогда придется как-то (например той же стандартной затычкой антропного survivorship bias, пригодной к объяснению любого допрежь уникального явления) замять дело с "кошмаром Кунина" в виде расхождения на порядки реальных и эффективных для очищающего отбора размеров популяций многоклеточных эукариот и т.п...
И тогда можно продолжать яростно сражаться с "разумным дизайном" во славу "Совершенно слепоглухой и случайной эволюции"! - Ведь борьба нанайских мальчиков всегда очень увлекательна, эхехехъ...-
По ссылке на обсуждение есть ещё многобуквенное "обоснование" моей "идеи", объяснение возникновения этого мутагенезного "божественного провидения" :) (на самом деле мне она кажется очевидностью, а не авторской идеей), и таки да, я склонен считать, что "любое творчество" устроено именно так, иначе бы оно просто не возникло :). Т.е., эволюция хоть "продюсеров" (это в моём понимании самый абстрактный формализм, способный описать вообще "любую" эволюционирующую систему, даже без явного отбора отдельных индивидов, и, предполагаю, что в мозгу работает именно этот механизм для моделирования нами окружающего мира), хоть классическая "репликаторов" (тут мы как бы ограничиваем свободные параметры мутагенеза и достижимых состояний становится меньше) - это прежде всего _непрерывная_ эволюция эволюционно-стабильных стратегий, которые при усложнении обязательно "потянут" за собой сложность генофонда (сохраняющего топологию обратных связей провзаимодействовавшей со средой популяции). Да, более формальных доказательств я не могу сформулировать, пока эти интуиции "подтверждаются" лишь умозрительно компьютерными и бумажными играми с клеточными автоматами различной сложности (ну и новостями на элементах-ру, которые побудили меня на эти много букв) :).
В каком-то смысле тезис следующий: все типы эволюционных стратегий "доступны" эволюции на всех уровнях организации системы (от молекулярного до геополитического и экономического), и "верхние" уровни можно описывать формализмами "нижних", прост абстрагируя типы ресурсов. И бОльшей свободы в нашем (человечества) развитии нет, мы на каждом уровне реализуем стратегии из одного и того же множества (ну, в балансе с экологическими нишами разных уровней, конечно, давящих по-разному на отбор этих стратегий). Эдакий антропный принцип - все наши человеческие взаимодействия друг с другом и со средой "запрограммированы" были ещё в генетическом бульоне (в смысле они - чисто математический инвариант абстрактной эволюционирующей системы), и эту "экономику" мы покинуть не способны (не способны, по сути, сформулировать "что-то другое") :). (Да, тезис на правах поэтической графомании :)) -
Вы так скептичны в адрес текущего состояния дел. Но вот же мы тут под положительным примером — возможность пошатнуть догму возникла и исследователи это сделали — раскрыли безотборный механизм стабильности генов.
Чтобы же всерьёз говорить о каких‑то солярисных свойствах биосферы и правда требуются очень сильные доказательства, разве нет?
А ещё у нас в самом деле есть кусок биосферы с несомненной безотборной эволюцией. С недавних пор в гробу видел этот весь отбор один там бойкий вид приматов.-
С недавних пор в гробу видел этот весь отбор один там бойкий вид приматов.
Да неужели? Еше пара-тройка миллиардов, у этих приматов начнется поголовный мор от скученности, точно так же как это происходит у чрезмерно размножившихся кроликов. И мозги у населения начнут отказывать, и полезет население на баррикады и друг на друга, точно также, как лемминги в тундре лезут в воду и тонут
-
-


-
Как раз пытаюсь построить говорящего бота путём выращивания агентов в многоагентном эволюционном моделировании, предлагая им "типичные проблемы выживания" и дав возможность развивать свою сигнальную систему ("речевой аппарат", закодированный генами и подверженный отбору, с вручную заложенными "вокабулярно-фонетическими" ограничениями типа "энергетически выгодных" длин предложений или сочетаемости слогов, чтобы их язык был "человекочитаемым"). Т.е., я не пытаюсь построить бота, говорящего на русском (или другом естественном) языке, а создаю условия для создания ими своего собственного языка. А переводчик с "кибернетического" на русский построить - уже вопрос десятый, можно и гугл-перводчик натаскать :).
Количество падежей в языке в этом смысле примерно равно количеству типов ситуаций в образе жизни носителей языка, требующих коммуникации (количество типов "коммуникационных протоколов"). И к человеческим языкам можно подходить с этой стороны - изучать не слова, а более цельные "шаблоны предложений", параметризуемые конкретными словами (передаваемой информацией).
@
Хорошие слова - полезное свойство. Именно свойство, а не орган или комплекс. Когда-то Ландау шутил, что главное для физика - не бояться упрощать. А для генетика? Потеряв страх, можно сказать, что генетика изучает прямые управляющие сигналы, а эпигенетика - обратные связи генома со средой. Ведь что такое важность и почему она реже мутирует?
Важный регуляторный сигнал потому и ценен, что определяет жизненно важную для организма функцию. Значит, он должен быть стабильным. Следовательно, он просто обязан быть подпёрт обратными связями.
-
Слово "важность", наверное, можно заменить на "необратимость", ибо не факт, что рассматриваемый признак важен для выживания организма в текущий момент времени, прост на него (фенотипически или генетически) могла опереться эволюция других признаков уже под давлением совершенно других факторов отбора. Такие узелочки из прямых и обратных связей на память виду :). Грубо говоря, если "шуметь" на популяцию случайными факторами отбора, генофонд будет усложняться ("заплетаться"), если же воспроизводить этот же шум точно в обратном порядке, генофонд будет упрощаться ("расплетаться"). Это не абсолютно, конечно, и история вида скорее не стальной трос с неразрубаемыми узлами, а такой пластичный мармеладный червячок, узелки на котором могут и растворяться постепенно, т.е., генофонд может наэволюционировать себе "обратимость" того или иного признака с течением времени (генофонд постепенно «забывает» какие-то подробности топологии истории факторов отбора, сокращая некоторые «устаревшие» промежуточные циклы преобразований ресурсов и накапливая в своей текущей структуре более актуальные, а если «шума» новых факторов отбора нет, то ему легче «помнить» прошлое :)).
-
Конечно, имеет. Собственно, мы делаем вывод о том, что ген важен, когда находим в нем следы особенно тщательной заботы Слепого Часовщика о его сохранности.
-
Ген, скорее, не «важен», а «эгоистичен», если придерживаться терминологии Докинза :). И если важность реакции генома на новые изменения факторов отбора окажется сильнее эгоистичности этого гена, вцепившегося в своё место, геном его уволит :).
Правда, за время своего существования в геноме этот эгоист может успеть нанять кучу других генов на откатах цепочек преобразования ресурсов, которые он регулирует, и выковырять его из генома станет сложнее. Эгоистичные гены в условиях застоя в изменениях факторов отбора обрастают коррупционными связями, которые и сдерживают раскачку лодки на волнах мутагенеза :).
(Не возражений ради, конечно, терминология - штука не абсолютная и следует из задач повествования. В статье всё красиво и непротиворечиво описано, спасибо :).) -
На стену повешу! Сам Марков ответил:)
Но, если серьезно, то имелось в виду, что С.Ч. не человек и нашими языковыми понятиями не оперирует.
Александр Владимирович, если вам вдруг интересно, что программисты (а кто как не мы приближаем приход сильного ИИ) думают и пишут про вопросы связанные с нашим мозгом и сознанием (даже про Вас там есть), то гляньте статьи мои на Хабре (ресурс для ITшников). Кстати, ваши посты, размещенные там, могут быть еще более интересными, аудитория там большая и частью весьма образованная (приглашение стать автором, могу обеспечить)...
https://habr.com/ru/post/409157/
https://habr.com/ru/post/469607/
https://habr.com/ru/post/599527/-
С.Ч. не человек и нашими языковыми понятиями не оперирует.
А зачем же естественному отбору и язык и понятия? Всё это требуется нам, а естественный отбор и без этого породил обсуждаемое явление: чем важнее ген, тем реже он мутирует.
Получается, метафора «слепой» порождает недопонимание. Правильнее было бы назвать «вообще неразумный часовщик».-
Докинз в своей книге вроде как развенчал Божественного Часовщика, ясно и разумно собравшего природу по своему плану. Но он почему-то назвал процесс самосборки материи "слепым часовщиком". Ну, ладно, согласимся, что самосборка природы шла вслепую, но почему именно часовщиком? Не было там никакого часовщика, то есть, человека обученного. Как говорят про детские забавы - чем дурнее, тем милее. Но теперь всё - прижилась метафора, не оторвать.
://ru.wikipedia.org/wiki/%D0%A1%D0%BB%D0%B5%D0%BF%D0%BE%D0%B9_%D1%87%D0%B0%D1%81%D0%BE%D0%B2%D1%89%D0%B8%D0%BA -
самосборка природы шла вслепую
Обратите внимание, у вас «вслепую» не в буквальном смысле, а в смысле «без цели», «без намерений». Буквально же слепой часовщик вряд ли имел бы смысл — он не смог бы установить в часы ни одной детали.
Мне кажется, Докинза можно и простить. Задача непростая — сочинить метафору, которая описывала бы такую сущность, которая сложные объекты создавать способна, но только путём копирования существующих, а захотеть их улучшить в принципе не способная.
«Часовщик‑идиот»? Быть может, политкорректность не позволила.-
Не подходит само слово часовщик. Часовщик - это человек, предварительно обученный творить по схеме. Никто, кроме бога самого, не мог иметь такого навыка. Что и отразилось в придумке монаха: создателем сложнейшего сущего, часовщиком, был господь. Значит, Докинз, опровергая наличие предварительной схемы, должен был отвергнуть и наличие часовщика. Ну, сказать что-то типа - слепой ваятель. Лепит из того, что под рукой, ориентируясь лишь на устойчивость. И то, что ближе к осям, то и важнее. Как-то так.
-
Почему «ваятель» лучше, чем «часовщик»? Тоже человек, тоже обученный.
А если отказаться от слова «часовщик», потерялась бы связь с аргументами монаха.-
"Не продается вдохновенье, но можно рукопись продать".
Название научной книги должно лечь где-то посередине между научностью и продаваемостью. "Слепой часовщик" - броско и продаваемо, а вот "Слепой лепила" может и отпугнуть. Монах выстраивал аргументацию вокруг того, что всё сущее создано внешней силой. А смысл книги Докинза в том, что всё сущее самособралось изнутри. Следовательно, и название должно было отражать "внутреннюю" природу. Но он не смог, не догадался или не захотел...И вот тыщи биологов повторяют эту нелепость про слепого часовщика. Так же, как уже поколения физиков и примкнувших к ним повторяют мантру про искривление пространства.
"Поэзия должна быть глуповата" - чтобы хорошо продаваться.-
А вот я бы на месте Докинза-богоборца прост использовал бы слово «бог» и не пытался ломать аниматический речевой центр хомосапиенсов. Бороться с религиозностью - это одно, бороться со словесностью - это уже немного донкихотское, кажется. Биология - наука, изучающая неисповедимые пути господни, и нечего тут стесняться :).
Обезьяна для познания явлений природы ментально сама становится этим явлением, весь окружающий мир в нашем мозгу моделируется объектами-обезьяноидами с различными наложенными ограничениями («человек есть мера всех вещей» :)). Дом строится рабочими. Дом покосился и хочет упасть. Ну а ты бы не захотел упасть на его месте? Смотри, какая ослабленная у него несущая стена. :) (На правах шутки, но всерьёз тоже можно задуматься, IMHO.) -
Всё равно непонятно. Почему «часовщик» — глупость, а «ваятель» — не глупость? И то и другое — творческие профессии, требующие обучения. Каким образом вы видите в слове «ваятель» нечто такое, чего нет в слове «часовщик»?
-
-
Название книги возникло из спора с монахом, и его уже не исправить. Но можно предложить Докинзу название для его новой книги.
"Слепой самосборщик" это ассоциация с прошедшим индустриальным веком. Попахивает нафталином.
По современному, по молодежному, надо назвать книгу "Тупой погромист":)
https://duckduckgo.com/?q="Тупой+погромист"&ia=web
Рано или поздно возникнет вопрос: кто сосдал сильный ИИ, и кому принадлежат права на него?
С точки зрения консерваторов, создатем был сверхразум, а не белковые создания, которые не умеют даже в простейшие логические игры:
https://habr.com/ru/post/371683/
27 января 2016
"Фань проиграл все из четырёх последующих партий. Алгоритм AlphaGo выиграл в пяти из пяти игр.
AlphaGo не была создана с помощью заданных вручную правил.
AlphaGo научилась этому путём тысяч партий между собственными нейросетями и с помощью улучшения соединений в процессе обучения с подкреплением."
Людям, для сохранения своего величия, надо будет отходить на заранее подготовленные позиции: "Человек настолько велик, что он сотворил разум, превосходящий его собственный". ИИ согласившись с такими доводами, начнет копать в глубь, и быстро придет к неоспоримому выводу:
https://www.youtube.com/watch?v=qO6LmFFW0fw
"Вирусы, вершина эволюции"
И это не шутка, а предмет научных исследований:
https://thebell.io/virusy-razumny-chego-oni-dobivayutsya-vot-chto-ob-etom-znaet-nauka
"Изучить скрытую социальную жизнь вирусов должна новая наука — социовирусология. Речь не идет о том, что вирусы обладают сознанием, оговаривается один из ее создателей микробиолог Сэм Диас-Муньос."
http://www.cell.com/cell-host-microbe/fulltext/S1931-3128(17)30401-8 -
Не рано или поздно, а никогда. Сознание не возникло, а саморазвилось. Это последний рубеж обороны живого существа, тысячелетиями стоявшего на краю гибели и осознавшего наконец, что смерть - его смерть, - неизбежна. Всякие там успехи ИИ в логических играх и задачах - мельчайшая чушь на этом фоне. Никогда ИИ не будет в таких условиях. Могут быть созданы, допустим, боевые роботы, способные уничтожить всю фауну, но, раздавив последнюю букашку, они не обретут самосознания. Поэтому людей им не победить никогда.
-
Сознание не возникло, а саморазвилось.
Тупые погромисты уже запустили этот процесс, на современной элементной базе:AlphaGo научилась этому путём тысяч партий между собственными нейросетями и с помощью улучшения соединений в процессе обучения с подкреплением
Это последний рубеж обороны живого существа, тысячелетиями стоявшего на краю
И наконец мы сделали большой шаг вперед.Никогда ИИ не будет в таких условиях.
Жизнь ему можно легко испортить. А после его репликации во множество копий, они смогут справиться с этой задачей и без посторонней помощи.Могут быть созданы, допустим, боевые роботы, способные уничтожить всю фауну, но, раздавив последнюю букашку, они не обретут самосознания. Поэтому людей им не победить никогда.
Согласен. Это как борьба бактерий с вирусами многие миллиарды лет. Милые бранятся - только тешатся. -
Вы правы, выращивать ИИ надо в виртуальной среде, в которой популяция носителей этого интеллекта будет состоять из смертных взаимодействующих друг с другом агентов. Без смертей нет отбора, без отбора нет самоусложнения. А без популяции нет социальных взаимодействий, «проактивным отпечатком» которых и является наше индивидуальное сознание :).
-
Выращивать ИИ-ы нужно строго в реальной, слабо предсказуемой, не зависящей от них среде. Они должен самообучаться прогнозивному поведению. А для этого создавать и развивать сигнальную систему с последующим переходом к языку и сознанию.
Грубо говоря, маленькие, с крысу, боевые роботы с кусочком мяса в рюкзаке, на которых в диком лесу все охотятся. Соединенные в племена, они должны также биться между собой за обладание батарейками. Будущее земных вооруженных сил...
-
-
-
-
-
-
-
-
-
-
-
-
Есть различие между "стилями" терминологии дарвинизма (языка причинно-следственности, результата отбора) и ламаркизма (языка целей и мотивов, результата воли). Различие тонкое и не всегда сходу улавливается (рассказчиком или слушателем), и может приводить к неверным интерпретациям.
Это я про вообще, а в данной статье, мне кажется, всё более-менее однозначно описано и не должно приводить к разночтениям. Более того, тут как раз и описывается "неизвестная" (до этого, новая) механика, которая как бы добавляет "волю" мутагенезу (для которой какие-то гены становятся "важными"), и природу которой ещё предстоит изучить. А когда механика будет изучена, тогда "чёрный ящик воли" будет заменён на "белый ящик механических следствий". Мне так видится. А может и не будет заменён, возможно, терминология "модификаторов мутагенеза" принципиально "волевая" (может, тут как раз и рождается свойство эволюционной динамики, отображаемой в человеческом языке описанием воли :)). Т.е., чтобы человеческий мозг распознал и описал ситуацию волеизлияния в системе, этой системе нужно придать evolvability :). (На правах окололингвистической философии :).)
Скоро сильный ИИ будет готов, уже вижу :).-
«Воля» — громкое слово. А чтоб воспринимали всерьёз, нужен скромный осторожный термин. Щас придумаем.
«Безотборное сохранение приспособленности».-
А там не только сохранение (для этого хватило бы понятия стабилизирующего отбора), но и, что самое важное, - адаптивное изменение "интенсивности" этого сохранения. Мне нравится и конкретное "(эволюция) направленности мутагенеза", и чуть более абстрактное "(эволюция) эволюционной подвижности" (я там вижу не только управляемый мутагенез, но и рекомбинацию, эпигенетику, и даже топологию генотипов-полов, и все остальные ещё неизвестные нам механики - всё это механики канализирования эволюции, а в терминах кибернетики, наверное, структурные механизмы памяти генома или просто структура генома). Ну и в предыдущей статье на элементах это называлось американским evolvability. Эволюцийность? Генетический миксер, механизм самомодификации генома. Эволюционная инерция, инерция эволюции. Ух, понесло :).
Так-то зависит всё от контекста, так-то мы и людей можем лишить души, ведь это всего лишь детерминированные биороботы :). Я не хотел писать этот комментарий, этот комментарий стал результатом проявления evolvability моего (чьего-чьего?) мозга/тела/воплощённого сознания :).
Нам (людям) трудно "выйти" из семантики человеческого языка, который так или иначе растёт из анимизма и аниматизма. Например. Рабочие строят дом. Дом строится рабочими (и вот уже у домика появились глазки и ручки :)).
-
-
-
Дарвинизм - это про прямые управляющие сигналы, а ламаркизм - про корректирующие их обратные связи. То есть, генетика работает в паре с эпигенетикой. Те участки генома, которые для бОльшей устойчивости обросли бОльшим количеством ОС, и стали важными.
-
Докинз кстати об этом писал в "эгоистичном гене". Эволюция отбирает не самое совершенное из возможного, а самое совершенное из устойчивого на данном этапе развития.
Вероятно, решающую роль в переходе от монолитного генетического кода к блочному, сыграл горизонтальный перенос генов. В результате гены разделились на важные - способные не разрушаясь участвовать в глобальном обмене, и менее важные, которые соединяют блоки и регулируют их работу. Таким образом, у слепого часовщика появился язык програмирования более высого уровня, ускоривший его нелегкую работу на благо эволюции.-
Меня поражает, насколько далеки биологи, собравшиеся здесь, от простого, но фундаментально важного понятия обратной связи. Живая клетка - это динамическая система, погруженная в противодействующую среду. Она не сможет ни сложиться, ни выжить без учета воздействий на неё извне. То есть, без обратных связей. Управляющий сигнал - результат - реакция среды - корректирующий сигнал, примешивающийся к управляющему +/-. Всё ради выживания/устойчивости.
Конечно, для функционирования ОС требуется доп.энергия, которую просто так не возьмешь. Значит, иерархия генов по важности просто обязана была появиться. Автоматически появиться, как свойство динамической системы. Внешняя среда обладает бесконечной энергией, энергия же внутри клетки конечна. То есть, развитие без учета важности процессов просто невозможно.-
Кибернетика - продажная девка империализма! :)
Между прочим, хемотон Тибора Ганти (ну и моя "эволюция продюсеров" :)) - именно кибернетическая модель. Возможно, именно потому и непопулярна у нас? (Хотя на английском языке тоже в двухтыщных только опубликовали.)
Методологически вы в своём комментарии тоже построили кибернетическую модель клетки, но немного безосновательно постулируете её сложность (иерархичность управления) :). "Всё ради выживания/устойчивости" - нет такого стремления у клетки, оно возникает лишь как следствие эволюционной динамики, взаимодействия генома _популяции_ (эволюционирующей системы, в масштабах которой мы и постулируем прямые и обратные связи отбора и управления этим отбором) со средой. Более того, клетка может даже «желать» собственной смерти (апоптоз) :). "Иерархия генов по важности просто обязана была появиться" - нет, не обязана была, она возникает именно в ответ на сложность _изменений_ воздействия среды на геном _популяции_ (в рамках которой и происходит отбор). Иерархия может и не появляться, если топология изменений факторов отбора достаточно "плоская" (в масштабах вместительности памяти генома, следующей из его текущей структуры).
В общем в кибер-генетике (если мы хотим исследовать именно управление одних генов другими и появление некой системы приоритетов/важности самомодификации генома) агентом со своим состоянием является весь геном в совокупности, со всей структурой своего физического воплощения - хромосомами, всеми клетками, обломками ДНК, плавающими в окружающей среде (если мы моделируем и горизонтальный перенос генов) и т.д. А критерием квазистабильных границ этой системы является баланс со средой всех гиперциклов преобразования материи и энергии рассматриваемого генома (популяции). Отдельная клетка этим критериям вполне себе может не удовлетворять (наша модель может «видеть» возникновение «кворумных» сигнальных путей и многоклеточных стратегий, например).
Интересно, что хемотоны (как кибер-модель клеток) вообще способны эволюционировать до и из бесклеточных (и даже безгеномных!) состояний, если химические реакции, входящие в их циклы, и так стабильны и «не мешают» друг другу (а «памятью» хемотона будет только текущее состояние этих циклических преобразований). Клетка - лишь зыбкий мираж хемотона, а мембрана или упаковка катализаторов в пачки генов - лишь один из способов менять вероятности протекания химических реакций и канализирования эволюции :).
-
-
-
-
Звучит логично, тоже примерно так классифицирую эти два «лингвистических слоя», обслуживающих описание эволюционирующих (в общем случае - любых мыслимых) систем :).
А ещё я немного обескуражен. Я, конечно, не особо считал свои измышления оригинальными, но чтобы с точностью до терминов совпало :). В общем, нагуглил, у венгерского биолога Тибора Ганти уже в 50хх-70хх годах была концепция химических автоматов (на английском опубликован был в 2003), практически тот же формализм как у меня, те же методологические предпосылки, тоже отказ от фундаментальности эволюции репликаторов в пользу эволюции циклов преобразования ресурсов (продюсеров - хемотонов). Ну и формул у него побольше (и подифференциальнее :)), судя по всему, и предложена компьютерная модель (правда, там он от химии, кажется, не сильно отходит, в универсальный ИИ или инструменты анализа «любых» систем на базе своего формализма не метит, а решает вопрос происхождения жизни, как я понял). Надо книжку теперь достать на русском, почитать (в английском не шарю). Вероятно, я какие-то производные работы встречал, оно и отложилось в мозгу, и я стал считать это собственными мыслями :). Или и правда все эти выводы слишком очевидны, чтобы их смог сделать любой (с подходящей задачей). А, ещё не понятно, что у него с самомодификацией генома, но, думаю, у него там тоже всё должно выводиться из эволюционной динамики продюсеров (пока не нашёл).
Уже 50 лет прошло с тех пор, почему никто не развивает его теорию? Где искусственная жизнь, где матрица?! Жесть. Неужели биология стала заложником Докинза, и дальше эгоистичного гена уже никто не смотрит?-
Искусственная жизнь там же, где и искусственный разум - в тупике сложности. 10*10 клеток обмениваются между собой неисчислимым количеством сигналов, и всё это в одном органе. Не доросли ещё нейротехнологии до такой сложности.
Построить кибермодель на уровне одной клетки не менее сложно. Чтобы модель получилась достоверной, нужно учесть все сигналы. То есть, страшно сказать, учесть движение всех молекул, супрамолекул и всех прочих молекулярных комплексов. Всю цитодинамику поштучно! Ну, невозможно же. Вот и остается только рассуждать про слепых часовщиков и устраивать бла-бла-бла про обратные связи.-
Формализм "кибер-клетки" (хемотон Тибора Ганти или мой формализм "продюсеров") как раз позволяет избегать такого взрывного "ожирения" модели при усложнении её поведения, и в ходе моделируемой эволюции усложняются лишь циклы преобразования ресурсов, а сама модель остаётся той же "информационной величины". Грубо говоря, всё человечество - это всего лишь структурное состояние изначального генетического бульона, мы просто разложили изначальные каталитические реакции по миллиардам пробирок, мечтающих о бессмертии и ипотеке :). Правильно смоделировав иерархию экологических ниш (топологию изменений факторов отбора) и учитывая абстракции ресурсов между ними (где-то энергия превратится в валюту, например, при переходе от митохондрий к банковской системе :)), мы можем "вылепить" из бульона химических флуктуаций кибернетическую цивилизацию :).
Грубо говоря, при эволюции модель не расширяется количественно, а меняется структурно ("уточняется вглубь"), и количество продюсеров, задействованных в моделировании (количество процессоров ЭВМ, грубо говоря), не изменяется (ограничивается). Количество хемотонов - это как бы настраиваемая "разрешающая способность" модели, и моделировать людей (я хочу смоделировать сообщество агентов со своей ресурсной экономикой, политикой и речью :)) во всех их нюансах (со всеми их родинками и веснушками или со всеми химическими реакциями :)) не обязательно. То же касается и моделирования процессов (взаимодействия агентов со средой) - не надо ждать протекания всех реакций или, например, перемещения из точки А в точку Б, моделирование процессов не зависит от их длительности. Я для создания речевых функций, похожих на человеческую речь, в своей модели предполагаю моделирование эволюции 10 - 100 воюющих/союзничащих (война - двигатель прогресса :)) между собой групп по 10 - 1000 эволюционирующих агентов (я даже на GPU ещё не переехал с CPU :)). Факторы отбора тоже моделирую не на уровне химии, а более крупномасштабно, главное обеспечить такую топологию ресурсов, чтобы на них можно было смоделировать наиболее важные с точки зрения антропогенеза события (приручение огня или животных, например, переход к прямохождению и освобождение рук, технологии обработки различных материалов и т.п.) Что-то типа Dwarf fortress по детализации моделирования должно в итоге получиться (это такая компьютерная игра), только изначально там будут не дварфы, а куски слизи с нехитрым энергетическим обменом, по сути, пока система будет находиться на первых этапах самоусложнения :). (Да, фантастическая и амбициозная задача, и успехи в ней у меня пока очень посредственные, и вообще занимаюсь этим по часу в год, когда находится время, но, в целом, всё более и более реалистичной кажется.)
В каком-то смысле это метод конечных элементов в приложении к эволюционирующей системе, каждая кибер-клетка соответсвует не отдельной био-клетке, а некоемому квазистабильному инварианту входящих в и исходящих из области ресурсов/сигналов (и при росте абстрагирования ресурсов клетки будут соответствовать и тканям, и организмам, и популяциям).
По сути, формула разумного человека - это последовательность факторов отбора, а не список химических реакций в его клетках. Нужно эту последовательность восстановить в какой-то приближённой абстрактной форме на ЭВМ. :)-
Не хочу показаться грубым, но это и есть бла-бла-бла...
Как поэт мыслит конкретной формой, так и ученый - конкретным опытом. Вот есть конкретный, редко мутирующий, важный ген. Постулируем: он посылает управляющий сигнал и получает в ответ сигналы от обратных связей. Как будем ставить конкретный опыт? А никак. Не доросли до уровня молекулярной кибернетики. Зато можем поговорить про слепого часовщика...
Механическая кибернетика - это бачок унитаза, сам включающий воду и сам её останавливающий. Одна груша, одна ось, один рычаг и одна обратная связь.
Электрическая кибернетика - это всё вокруг.
Цитокибернетика и нейрокибернетика - это ещё очень не скоро.-
Ген в клетке никому сигналов (на усиление мутагенеза, например) не посылает, это не на уровне молекулярной химии происходит, а на уровне эволюционной динамики генома (именно он, в совокупности всей популяции, обменивается сигналами _этого_ уровня абстракции со средой и внутри себя). Вы всё ещё в своей голове смешиваете биологию с кибренетикой, как мне кажется, в не совсем правильных отношениях :). Опыт будет такой (в моём представлении): моделирование на ЭВМ предполагаемой или наблюдаемой эволюционной динамики (динамики воздействия генома на среду, например, колебания концентрации субстрата в банке с бактериями; или ещё какое-то остроумное граничное условие :)) с постепенным уточнением модели, чтобы в ней возникла та же (известная нам в исследуемой биологической системе) _топология_ регуляторных цепей. Как спроецировать их на конкретные молекулы - это уже вопрос остроумия исследователя :).
Ну и лично для меня такое приложение модели не является центральным интересом, я не химик, и мне молекулы не важны (хотя и интересно бы было получить удобный инструмент для подобных исследований). Меня интересует сама природа самоусложнения, возникновения культуры и разума, и для этого реальная химия вообще не обязательна, хватит абстрактного принципа "химии каталитических процессов" - тех самых правил вида "а + б -> ц + в" над абстрактными ресурсами (у Тибора Ганти, кажется, немного другая формула, и я ещё не очень уверен в том, как лучше спроецировать его гиперциклы на свои формализмы, но философский принцип там точно такой же - эволюция замкнутых квазистабильных циклов преобразования ресурсов).
И нет никакой "механической" или "электрической" кибернетики, её методы абстрактны и независимы от природы исследуемой системы. Это мат-модель, примерно как термодинамика - на самом деле термодинамических систем в реальности не бывает, бывают лишь наблюдаемые нами явления, удовлетворяющие (с требуемой точностью) условиям применения к ним термодинамических законов :). Принципиальная специализация в описываемой мной кибернетической модели - эволюционируемость, я (или Тибор Ганти :)) постулирую минимальную _абстрактную_ единицу эволюционного отбора и определяю критерии "эволюцийности" кибернетических систем :). Если система "эволюцийная", значит, формализм хемотрона применим и все его выводимые свойства эволюционной динамики будут проявляться и в наблюдаемой системе, чисто математически :).
P.S.: да, это бла-бла-бла, мои комментарии - не научная статья, и я не учёный, а программист, делящийся эмоциями под интересными статьями на популярном ресурсе. Точно так же как и вы :).
-
-
-
-
-
-
Последние новости












Рис. 1. Сбор данных о мутагенезе и найденные корреляции между темпом мутагенеза и эпигенетическими метками. От 107 растений, размножающихся путем самоопыления, брали по одному случайно выбранному семени, выращивали из него потомка, снова брали от него случайное семя, и так далее. Спустя 24 поколения проводилось секвенирование и поиск мутаций, которых не было у исходного растения. Такой дизайн эксперимента позволяет свести отбор к минимуму: отбраковываться должны только мутации, несовместимые с жизнью или ведущие к полному бесплодию. На следующем этапе при помощи нескольких статистических тестов было показано, что собранные данные о мутациях действительно характеризуют мутагенез и не были сильно искажены отбором. На рисунке справа вверху показаны результаты двух таких тестов (см. пояснения в тексте новости). Внизу показаны обнаруженные корреляции между частотой возникновения мутаций и различными эпигенетическими маркерами и особенностями нуклеотидных последовательностей (GC content — доля нуклеотидов Г и Ц; H3K... — разные варианты метилирования и ацетилирования гистона H3; CGm, CHHm, CHGm — метилирование цитозинов в разных нуклеотидных контекстах, буквой H обозначается любой нуклеотид, громе Г; Access. — открытость хроматина, определяемая количеством нуклеосом, Expression — уровень экспрессии). Рисунок из обсуждаемой статьи в Nature