Выделена новая супергруппа эукариот

Новое молекулярно-филогенетическое исследование позволило выделить эволюционную ветвь эукариот, имеющую очень высокий ранг, но состоящую целиком из одноклеточных организмов: она включает малоизвестные группы коллодиктионид, мантамодад и ригифилид. При этом ее ранг значительно выше, чем, например, ранг ветви заднежгутиковых, в которую входят грибы вместе с многоклеточными животными (последняя находится внутри мегагруппы Amorphea). Если это открытие подтвердится, оно может потребовать серьезного пересмотра представлений о том, как устроено эволюционное древо эукариот в целом.
Одним из самых ярких достижений биологии начала XXI века стало создание совершенно новой системы эукариот — обладателей сложной клетки. Эукариоты — гигантская группа организмов, в которую среди прочих входят многоклеточные животные и зеленые растения. Область биологии, исследующая границы и родственные связи групп очень высокого ранга (таких, как животные и растения), называется мегасистематикой. Попросту говоря, мегасистематика — это «царствоведение» (В. А. Красильников, 2004. «Царствоведение», или мегасистематика). Напомним, что царством называется высшая систематическая единица живой природы; в традиционной классификации царствами считались животные, растения, грибы. Правда, в современной биологии термин «царство», как и названия других традиционных систематических рангов, понемногу выходит из употребления.
Сейчас выделяется от 5 до 8 групп эукариот, соответствующих главным эволюционным ветвям — они называются супергруппами. В свою очередь, супергруппы объединяются в совсем уж небольшое число мегагрупп, которых в последнее время насчитывают три.
Мегагруппа, к которой относимся мы сами, называется Amorphea или Unikonta. Кроме многоклеточных животных, в нее входят воротничковые жгутиконосцы, грибы, многочисленные голые и раковинные амебы, слизевики (точнее, миксомицеты) и еще несколько групп одноклеточных и колониальных организмов, малоизвестных широкой публике (см., например: Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). Название Amorphea буквально означает «бесформенные». Наша мегагруппа названа так потому, что многие ее одноклеточные представители склонны использовать для движения или для питания временные клеточные выросты — ложноножки, а клетка, регулярно выпускающая и втягивающая ложноножки, по определению не может иметь постоянной формы. Второе название этой мегагруппы — Unikonta — значит «одножгутиковые». Дело в том, что у тех одноклеточных представителей этой группы, которые передвигаются с помощью жгутиков (такие существа называются жгутиконосцами), жгутик действительно чаще всего один, в то время как у членов других мегагрупп эукариот их обычно два. Но это не абсолютный закон: обладатели двух жгутиков среди униконтов, особенно примитивных, тоже встречаются.
Самая разнообразная из трех мегагрупп эукариот — безусловно, мегагруппа Diaphoretickes, название которой по-гречески и значит «разнообразные». В эту мегагруппу входят всевозможные водоросли (бурые, золотистые, желто-зеленые, диатомовые, красные, зеленые, харовые), высшие растения, ложные грибы (оомицеты), споровики, инфузории, радиолярии, фораминиферы, — и этот список более чем далек от полноты. Неудивительно, что внутренняя система этой мегагруппы сложна, запутана и до сих пор полна нерешенных вопросов (см., например: Новооткрытый жгутиконосец проливает свет на раннюю эволюцию эукариот, «Элементы», 17.01.2018). Подавляющее большинство эукариот, имеющих хлоропласты и способных к фотосинтезу, относится именно к мегагруппе Diaphoretickes. Другое ее название — Corticata, что означает «имеющие кортекс» (в данном случае этим словом называется укрепленный внешний слой клетки). Развитие прочного кортекса в общем случае мешает питанию животного типа, основанному на активном захвате пищевых объектов, и лучше сочетается с питанием растительного типа — фотосинтезом. Но, во-первых, тут встречаются исключения (например, инфузории обошли это ограничение и остались хищниками, создав постоянный клеточный рот), а во-вторых, само понятие «кортекс», честно говоря, довольно расплывчато. Структуры, укрепляющие внешний слой клетки, бывают устроены очень по-разному и сочетаются с различным образом жизни, так что далеко не все кортикаты фотосинтезируют; среди них есть не только одноклеточные хищники, но и паразиты, и сапротрофы, питающиеся мертвыми остатками других организмов.
Третья мегагруппа эукариот называется Excavata. Ее название происходит от латинских слов ex — внешний и cava — полость, борозда (имеется в виду характерная для представителей этой мегагруппы ротовая бороздка — продольное углубление на той стороне клетки, которая условно считается брюшной). Мегагруппа Excavata состоит преимущественно из жгутиконосцев; колониальных форм в ней мало, а обладателей многоклеточности в строгом смысле слова нет вовсе. Именно к экскаватам относятся жгутиконосцы, вошедшие в школьные учебники зоологии — фотосинтезирующая эвглена и хищный бодо.
Итак, эволюционное древо эукариот распадается на три мегаствола: Amorphea (= Unikonta), Diaphoretickes (= Corticata) и Excavata. Примем во внимание, что это предельно беглый обзор; для обсуждения более дробных подразделений — даже такого ранга, как многоклеточные животные или высшие растения — в короткой статье нет места, ибо их слишком много. Но даже столь общая картина позволяет сделать несколько важных выводов.
Во-первых, рассмотренная нами система не имеет ничего общего с той системой живой природы, которая господствовала в учебниках на протяжении XX века. Например, большинство тех организмов, которые в классической биологии назывались «одноклеточными животными», на самом деле относится или к мегагруппе Diaphoretickes, или к мегагруппе Excavata. Это означает, что они не более родственны животным, чем, скажем, какие-нибудь зеленые растения. С другой стороны, довольно близкими (по меркам мегасистемы) родственниками многоклеточных животных оказались грибы — раньше сближать эти группы было совершенно не принято.
Во-вторых, колоссальное место в современной системе эукариот занимают организмы, которые, в отличие от всем известных животных и растений, отнюдь не являются многоклеточными (во всяком случае, истинно многоклеточными). Еще с XIX века их принято собирательно называть протистами, то есть буквально «простейшими». К протистам в самом широком понимании этого термина можно при желании отнести всех эукариот, кроме многоклеточных животных и высших растений (J. O. Corliss, 1989. Protistan diversity and origins of multicellular/multitissued organisms). Гораздо чаще протистами называют все же только одноклеточных эукариот (или колониальных — при условии, что колонии состоят из однотипных клеток, способных в принципе жить отдельно). Но таких все равно очень много, и из них могут состоять целые супергруппы. По сути, все основание эволюционного древа эукариот слагается из протистов. Группы, состоящие целиком из многоклеточных организмов, образуют на этом древе лишь редкие «выбросы». Например, ветвей такого порядка, как многоклеточные животные, на эволюционном древе эукариот можно насчитать несколько сотен, и в большинстве из них истинная многоклеточность никогда не возникала (F. Burki, 2014. The eukaryotic tree of life from a global phylogenomic perspective).
В-третьих, примитивными членами всех без исключения мегагрупп и супергрупп эукариот оказались одноклеточные протисты, передвигающиеся с помощью жгутиков, то есть уже не раз упоминавшиеся жгутиконосцы. Многие эволюционные ветви достаточно высокого ранга состоят из жгутиконосцев вообще целиком. Ни одна другая жизненная форма не имеет на эволюционном древе эукариот такого всеохватывающего распространения. Это — явное свидетельство того, что общий предок всех современных эукариот тоже был жгутиконосцем (скорее всего, имевшим 2 жгутика).
Но в любом случае система эукариот не исчерпывается этим простым описанием. Дело в том, что существует не столь уж малое количество эволюционных линий протистов, которые, судя по молекулярным данным, не входят ни в одну известную супергруппу, и даже их принадлежность к мегагруппам зачастую остается под вопросом. В последнее время их называют «линиями-сиротами» («orphan» lineages). В подавляющем большинстве это — конечно же — жгутиконосцы. К тому же многие из «линий-сирот» явно расположены очень близко к общему корню эукариотного древа. Именно поэтому уточнение их родственных связей может оказаться крайне важным для понимания эволюции эукариот в целом.
Новая супергруппа
Недавно вышла очередная статья, уточняющая структуру эволюционного древа эукариот на основе молекулярно-биологических данных. Список авторов этой статьи замыкают известные канадские биологи Аластер Симпсон (Alastair Simpson) и Эндрю Роджер (Andrew Roger), которые уже много лет решают молекулярными методами проблемы мегасистематики эукариот; их мнение служит хорошим показателем текущего положения дел в этой области (см., например: A. G. B. Simpson, A. J. Roger, 2004. The real ‘kingdoms’ of eukaryotes).
На этот раз в центре внимания Роджера, Симпсона и их коллег оказались четыре группы одноклеточных организмов, относящихся к «линиям-сиротам» (рис. 1).
- Коллодиктион (см. Collodictyon) — пресноводный жгутиконосец с 4 жгутиками и небольшими ложноножками, склонный к питанию крупной по своим меркам добычей. Вместе с родственным жгутиконосцем, имеющим два жгутика (см. Diphylleia), он выделяется в семейство коллодиктионид (см. Collodictyonidae).
- Анкиромонады (см. Ancyromonas) — жгутиконосцы, живущие в воде или почве. Для них характерно наличие двух жгутиков, один из которых направлен вперед, а другой назад.
- Мантамонады (см. Mantamonas) — морские жгутиконосцы с двумя жгутиками и небольшими ложноножками, живущие в прибрежных донных осадках.
- Ригифилиды (см. Rigifila) — пресноводные протисты без жгутиков, зато с ложноножками особого типа — длинными и тонкими (они называются филоподиями).
Роджер, Симпсон и их коллеги построили общее древо эукариот, включающее перечисленные «линии-сироты» вместе много с кем еще и основанное на солидном наборе данных — программа обработала выравненные последовательности 351 белка (97 000 аминокислотных остатков). Результаты получились интересные.
Прежде всего, оказалось, что три из четырех названных «линий-сирот» — коллодиктиониды, мантамонады и ригифилиды — образуют на древе единую компактную эволюционную ветвь, вполне заслуживающую статуса супергруппы. Гипотеза о единстве этой ветви обоснована молекулярными данными достаточно хорошо, чтобы новая супергруппа получила предварительное название — CRuM (сокращение от названий входящих в нее семейств). Это — совершенно новое объединение. Раньше коллодиктионид, мантамонад и ригифилид куда только не помещали, но сейчас авторы готовы настаивать, что решили задачу верно: их новая гипотеза имеет очень высокую поддержку.
На общем древе ветвь CRuM примыкает к основанию ветви Amorphea (= Unikonta), образуя сестринскую группу по отношению к ней (рис. 2).
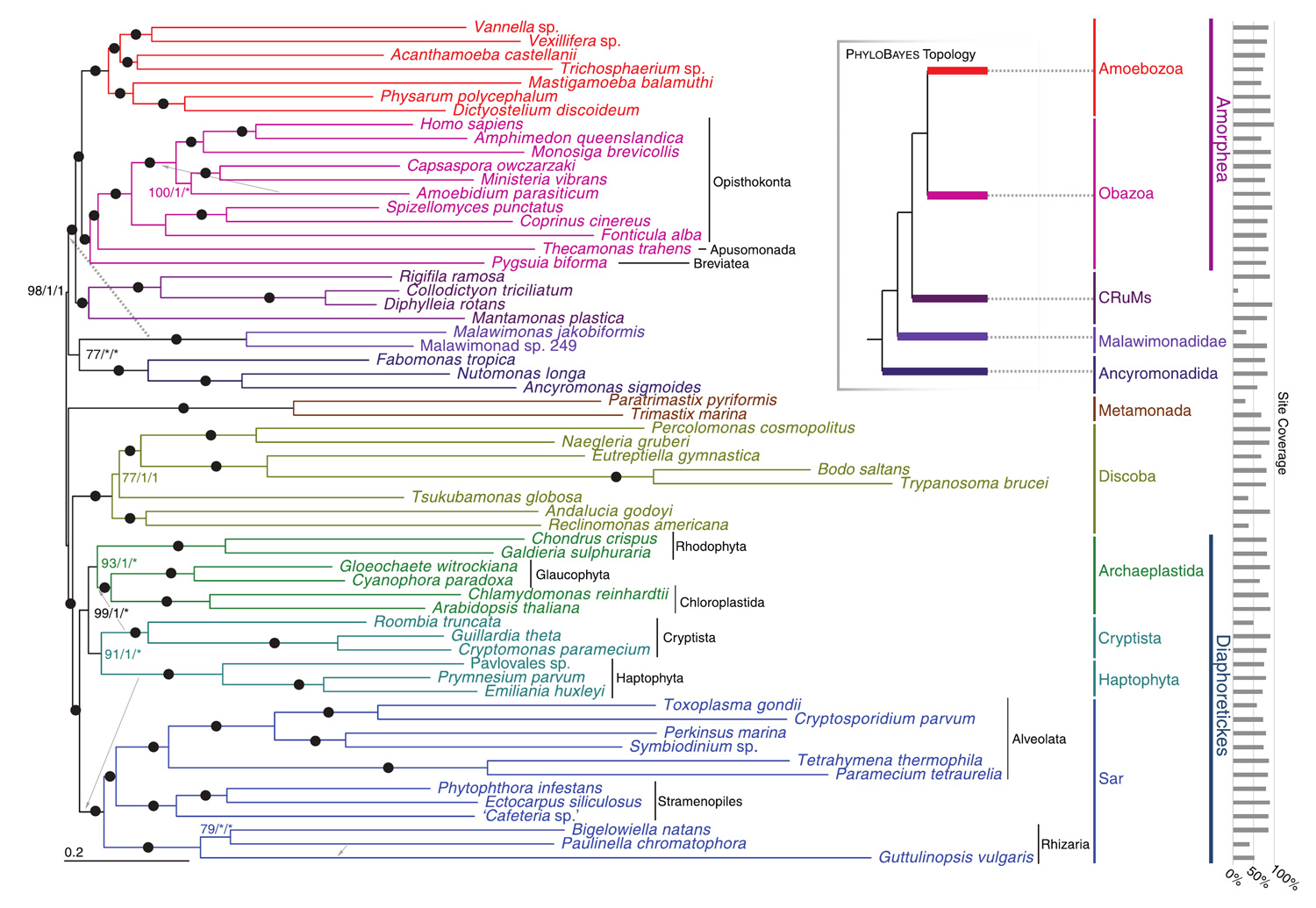
Рис. 2. Новая версия эволюционного древа эукариот. Иллюстрация из обсуждаемой статьи в Genome Biology and Evolution
Третья упомянутая нами «линия-сирота» — анкиромонады — пока осталась в одиночестве. На общем древе ветвь анкиромонад находится недалеко от основания ветви Amorphea (= Unikonta), но все же дальше от него, чем CRuM. Кстати, примерно такое же положение на общем древе занимает и еще одна маленькая группа пресноводных жгутиконосцев — малавимонады (см. Malawimonas). Однако в единую группу анкиромонады и малавимонады не сливаются (во всяком случае, данные на этот счет неоднозначны).
Получен и еще один результат, не вынесенный в заголовок обсуждаемой статьи, но тем не менее очень интересный. Симпсон, Роджер и их коллеги показали, что мегагруппа Excavata скорее всего на самом деле не является единой ветвью. Согласно полученным данным, эта группа, почти целиком состоящая из жгутиконосцев, распадается на две эволюционных линии, одна из которых несколько примитивнее другой. Первая эволюционная линия «экскаватных» жгутиконосцев называется метамонадами (Metamonada), вторая — дискобами (Discoba); именно к дискобам относятся, например, упомянутые выше эвглена и бодо. Если все это верно, то получается, что Excavata — не ветвь, а эволюционный уровень. Этот вывод нельзя назвать потрясающей сенсацией, потому что близкие к нему гипотезы высказывались и раньше, причем достаточно регулярно (см., например: T. Cavalier-Smith et al., 2014. Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa). Тем не менее, любые новые данные здесь достойны пристального внимания. Речь ведь идет о том, как устроено все древо эукариот целиком и с чего начиналась их эволюция — не больше и не меньше.
Почему сие важно?
Знаменитый генетик Николай Владимирович Тимофеев-Ресовский имел обыкновение на научных семинарах, выслушав докладчика, громко спрашивать: «Почему сие важно в-пятых?» Этот заимствованный из русской классики вопрос призывал участников как бы отступить на шаг, чтобы увидеть проблему в целом и понять точно — какое место в общей картине занимают обнаруженные факты.
При обсуждении научных новостей эта постановка вопроса тоже бывает полезна. Действительно, какое значение для нашего мировоззрения может иметь очередная перетасовка запутанных родственных связей между мало кому известными эукариотными микроорганизмами, живущими где-нибудь в далеких морях или болотах? И на какие детали тут стоит обращать внимание, а на какие нет?
Чтобы разобраться в этом, начнем с фактов, которые мы знаем более-менее твердо. Все современные эукариоты имеют единое происхождение: они произошли от одного предка, а не от нескольких эволюционных линий. Никаких эпизодов «параллельной эукариотизации», насколько можно судить, не было — по крайней мере, на тех эволюционных уровнях, которые доступны сейчас для изучения (см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). Не вызывает сомнений, что общий предок всех современных эукариот был «полностью оперившимся», типичным по нашим меркам эукариотом, у которого уже было полностью сформированное ядро, цитоскелет, эндоплазматическая сеть, а также митохондрии и жгутики. Облик этого предполагаемого предка в большой степени сохранили коллодиктиониды, мантамонады, анкиромонады, малавимонады (и некоторые другие жгутиконосцы тоже). Не случайно все они, в общем, достаточно сильно похожи друг на друга. Итак, есть серьезные основания считать, что общий предок современных эукариот был гетеротрофным жгутиконосцем, который вел придонный образ жизни, питался в основном бактериями, имел два жгутика и в придачу к ним небольшие ложноножки, служившие на том этапе больше для питания, чем для движения.
В каком же порядке от этого общего предка расходились эволюционные ветви? Тут начинаются сложности. Почти все основные супергруппы эукариот молекулярная систематика (стоило ей только возникнуть) выделила довольно легко, но вот установить их родственные отношения между собой оказалось очень трудно. Современная систематика, как правило, соблюдает методические ограничения кладистики, где по умолчанию предполагается, что эволюционное древо может ветвиться только дихотомически: в каждом узле каждая ветвь делится строго надвое. Поэтому молекулярные систематики видят свою задачу в сведении эволюционного древа к последовательности развилок надвое — дихотомий. И внутри эукариотных супергрупп это в целом удается. Проблемы начинаются при попытке раскрыть самую основу эукариотного древа — ту его часть, где «действующими лицами» становятся объединения супергрупп, мегагруппы и, конечно, «линии-сироты». Вот здесь сведение к набору дихотомий или не удается вовсе, или возникающие при этом гипотезы имеют недостаточную поддержку и легко ставятся под сомнение другими группами исследователей (см., например: D. He et al., 2014. An alternative root for the eukaryote tree of life). Почему?
Еще на рубеже XX и XXI веков французский биолог Эрве Филипп (Herve Philippe) предположил, что эта неопределенность имеет фундаментальную причину, связанную с особенностями ранней эволюции эукариот (см., например: H. Philippe et al., 2000. The new phylogeny of eukaryotes). Когда в истории жизни на Земле возникает что-то принципиально новое, это может сопровождаться кладогенезом — распадом маленькой предковой группы сразу на несколько эволюционных ветвей довольно высокого порядка. Кладогенез — вполне реальный способ эволюции. При этом свести его к последовательности дихотомий в общем случае принципиально невозможно, сколько ни старайся: что поделать, если группа-предок за короткое по эволюционным меркам время распалась не на две ветви, а на 3, 4, 5 или 6? Здесь кладистический подход просто обязан дать сбой. Эрве Филипп считал, что именно такая ситуация и наблюдается в начале эволюции эукариот (во всяком случае, их современных групп).
Не будем спешить с выводами. Даже если Филипп совершенно прав (что не доказано), это ни в коем случае не значит, что исследования, подобные тем, которые сейчас ведут Симпсон и Роджер, бесполезны. Совсем нет — эти исследования в любом случае дают новую, очень ценную информацию. Но вот помещение той или иной «линии-сироты» в ту или иную точку общего древа теперь может и не иметь решающего значения. Потому что биологический смысл складывающейся картины в любом случае уже приблизительно понятен — перестановки отдельных линий (которые вполне могут еще случиться) вряд ли его изменят. Вероятно, все современные эукариоты произошли от очень компактной группы предков, которые были жгутиконосцами, в целом похожими на современных анкиромонад или коллодиктионид. После серии быстрых расхождений (порядок которых, честно говоря, не столь важен) оформилось несколько эволюционных линий — например, на рис. 3 их семь, и порядок их расположения относительно друг друга там соответствует тому, что установили Симпсон и Роджер в обсуждаемой работе. Вероятно, это и есть картина кладогенеза.

Рис. 3. Сильно упрощенная версия предыдущего древа. Красный треугольник обозначает предполагаемый общий корень и указывает на общего предка всех современных эукариот
Остается выяснить, где же находится «корень» эукариотного древа? Это — очень непростой вопрос. Симпсон и Роджер принимают версию, согласно которой «корень» общего древа располагается между анкиромонадами и метамонадами, то есть между двумя группами жгутиконосцев (R. Derelle et al., 2015. Bacterial proteins pinpoint a single eukaryotic root). В некотором смысле можно сказать, что все эукариоты — жгутиконосцы. Просто некоторые из них сохранили эту жизненную форму до сих пор, а другие преобразовали ее, создав причудливые организмы амеб, инфузорий, грибов, растений, животных.
Источник: Matthew W. Brown, Aaron A. Heiss, Ryoma Kamikawa, Yuji Inagaki, Akinori Yabuki, Alexander K. Tice, Takashi Shiratori, Ken-Ichiro Ishida, Tetsuo Hashimoto, Alastair G. B. Simpson and Andrew J. Roger. Phylogenomics places orphan protistan lineages in a novel eukaryotic supergroup // Genome Biology and Evolution. 2018. V. 10. № 2. P. 427–433. DOI: 10.1093/gbe/evy014.
Сергей Ястребов
-
А не пора ли, ввиду многообразия характеристик и сложного родства, перейти от самой примитивной организации сети - иерархического дерева к нормальной сети - многосвязному графу?
-
С чего бы? Древо эукариот - нормальное древо, а никакой не "многосвязный граф". Слияние ветвей (в том масштабе, который мы сейчас обсуждаем; гибридогенное видообразование сейчас не считается, потому что оно бывает только между близкими видами и в этом масштабе незаметно), так вот, слияние ветвей на древе эукариот бывает только в одном известном мне случае: при возникновении вторичных хлоропластов (это показано вот здесь: http://2.bp.blogspot.com/_FeaU01D-3wI/TCR9N5ZgX2I/AAAAAAAABs
g/opmEec5giRQ/s1600/Keeling+2004+Am+J+Bot.jpeg). В остальном - древо и древо. В данном же обзоре вторичные хлоропласты вообще не фигурируют - они все приобретаются уже внутри супергрупп, в небольших ветвях, которые в обсуждение не попали. Так что нет причин переходить от древа к какой-то там сети. -
А как же метахондрии? А как же идея симбиотического организма? А как же горизонтальный генный перенос, если строить связи по геному? А сходные органеллы в разных царствах?
Более того, генетический ландшафт вида (множества особей с генетическим разнообразием) вовсе не линия, а полоса. Дискретизация дерева связана с давлением отбора. Если добавить виртуальные оборванные линии, то это будет не сеть, а ткань. Как это в постмодернизме ... ризома.-
Приобретение митохондрий (если Вы их имеете в виду ;)) было единичным событием, и оно случилось до того, как началось "действие", охваченное данным обзором. У обшего предка современных эукариот митохондрии уже были.
О симбиотическом организме я уже сказал. Единственная ситуация, когда в результате симбиоза между эукариотами действительно возникает новый организм - это приобретение вторичных хлоропластов. Но и оно находится за рамками данного обзора.
Что касается горизонтального переноса генов, то он тут просто ни при чем - мы ведь обсуждаем древо организмов.
Отсылки к ГПГ, к ризоме и к постмодернизму здесь подобны вот чему. Представим, что мы решаем задачу о том, чьим сыном был... ну, чтобы никого не обидеть, скажем - царь Дмитрий 1-й. Пусть у нас есть несколько версий, между которыми историки как-то выбирают. И тут является некий постмодернист и говорит: "А не пора ли, ввиду многообразия характеристик и сложного родства, перейти от иерархического дерева к ризоме? Это же так современно!"
Действительно, почему бы не считать, что Лжедмитрий получил, ну скажем, 40% генов от Гришки Отрепьева, еще 40% - от Ивана Грозного, а остальные 20% от какого-нибудь неведомого боярского ставленника? И все довольны. Великолепное постмодернистское решение.
Вот и применительно к древу эукариот оно выглядит так же.-
Птицы с коровами и Гришка с Иваном торчат далеко на кончиках ветвей. А суп царств одноклеточных стремительно начал разрастаться по мере того, как стали внимательно приглядываться к лужам под ногами. Ещё недавно отнесение грибов к третьему царству было смелым, а сейчас царства и группы в царствах появляются раз в пятилетку. Перекидывание видов на разные ветви в связи с появляющимися новыми подробностями говорит мне о сомнительности принципов классификации. Ex: хромисты весьма преуспели в симбиозе и гулянии по систематике.
-
Так вот я склонен думать, что _современные_ эукариоты (включая их общего предка) находятся по ту сторону водораздела, что и птицы с коровами и Гришка с Иваном - а не по ту, где бактерии, археи, прогеноты и прочий первичный суп.
Про симбиоз у хромистов я уже сказал дважды, третий раз, извините, не буду. А что я думаю о "сомнительности принципов классификации" - сказано, собственно, в статье, в последней части. Некая сомнительность действительно налицо, но вот никакая ризома тут ни при чем. Причины "сомнительности" совсем другие.
-
-
>А суп царств одноклеточных
Прежде чем рассуждать о ГПГ и ризомах, нужно перестать употреблять давно в этом контексте устаревшие понятия "одноклеточных/многоклеточных", а начинать прежде всего с определения о ком идёт речь - прокариотах или эукариотах - ибо эта дихотомия - вторая после клеточных/неклеточных форм в современной биологии.
Сетевой подход действительно имеет смысл в классификации прокариот - но и там далеко не тотальный солярисный суп, ибо репродуктивная изоляция - хотя и относительная, но планомерно возрастающая с таксономической удаленностью - там таки вполне имеется: в грампозитивную бактерию, например, плазмидный вектор грамнегативных просто так не пересадишь. Поэтому там тоже никакая не ризома, а сеть или, по выражению Е.Кунина, "лес" деревьев, где стволы можно отнести к типам (хотя их, пожалуй, тоже следовало бы именовать царствами/супергруппами - даже с большим основанием).
Что же касается эукариот, то там распространенность ГПГ резко идёт на убыль (по-видимому этому способствует компартментализация клетки) настолько, что никакой существенной помехи древовидной таксономии он не представляет. Все эти случаи - от хромист до насекомых с карсонеллами и вольбахиями и бделлоидных коловраток можно просто перечислять отдельными примечаниями.
А если озаботится ещё и вирусным генетическим наследием, то тогда нужно и к формуле "человек произошел от обезьяны" всегда добавлять "...и десятка ретровирусов". Оно кому надо кроме генетиков-ретровирусологов? Ну вот и. -
Нужно "родственников" среди всех Hominoidea секвентировать по вопросу о ретровирусов, найти на все ветвления оценку времени расхождения.
Конечно вплоть для гоминидов это давно сделали, но что-то там возраст общего предка с гориллой может расходиться с более поздними оценками (в пользу того, что старые оценки были 6-8 My для обоих видов гоминин, а сейчас кажется 7 для шимпанзе и 10 для гориллы).
-
-
-
-


А вот в технологической эволюции такое сплошь и рядом.
-
>Млекопитающие с птицами вряд ли произведут гибрид.
А те и другие с (ретро)вирусами - запросто.
А насекомые - ещё и с бактериями. А бделлоидные коловратки - вообще с кем угодно.
http://www.evolbiol.ru/lgt2008/lgt2008.htm
http://elementy.ru/novosti_nauki/430449/Gorizontalnyy_perenos_genov_privodit_k_novomu_rezhimu_evolyutsii
По поводу Excavata - вполне резонно и раньше отделяли Discoba от Metamonada, с Malawimonadidae возможно обособляющихся в Loukozoa. Кто-то из этих же соображений вовсе идет на таран, ломая Excavata, Malawimonadidae, что само по себе обычно и делается, вместе с Metamonada кладя рядом с Podiata или в них же, а Discoba не без оснований отправляя ближе к Diaphoretickes. Но верно отмечено, что разбиение Excavata на две вполне различные ветви это не новинка.
Новые же данные молекулярной биологии, будучи объективными, сами по себе и заключают основной смысл подобных работ, когда они верно получены, так что никаких претензий, вне зависимости от новизны точек зрения, быть к их авторам не может.
-
Ну во-первых, как Вы сами отмечаете, в группу Varisulca входят анкиромонады, и в таком контексте это довольно существенно. Во-вторых, эту группу выделил Кавалье-Смит, а он выделил в разное время столько таксонов, что современное научное сообщество, по-моему, уже приучилось их игнорировать, мол, на каждый чих Кавалье-Смита не наздравствуешься. В целом - тут я с Вами совершенно согласен - мне самому не нравится тенденция по каждому малейшему поводу придумывать новые названия, вместо того, чтобы "отредактировать" имеющиеся таксоны (например, я совсем не уверен, что таксон Chromalveolata действительно надо было так поспешно выкидывать на помойку). Но - таково положение дел.
Честно говоря, лично я думаю, что изначально ВСЕ супергруппы эукариот были "микроцарствами", и было их 15-20. Потом некоторые из них разрослись, а остальные так до сих пор "микроцарствами" и остались.-
Логично. Просто благодаря тому, что основные названия все же обычно сохраняют, а меняются преимущественно группировки, и придумываются новые наименования для новых объединений, типа SAR с Hacrobia вместо Chromalveolata, трудно видеть клады как нечто конкретное, то есть включающее строго определенные супергруппы. Хотя бы примерный вид древа вырисовался наконец в последние пару лет, за счет работ по всем эукариотам вплоть до приматов, так что почему бы не заниматься теперь как раз частными группировками да палеонтологической систематикой. Без резких открытий, хотя бы порядка этого (https://elementy.ru/novosti_nauki/433183/Novootkrytyy_zhgut
ikonosets_prolivaet_svet_na_rannyuyu_evolyutsiyu_eukariot) и горы прочих неклассифицированных организмов, складированных на том же NCBI, а также обнаружения ошибок в неверном восприятии данных, все к тому и сводится теперь: к "а я бы поставил этих сюда, а тех - туда". Вполне нормально, ведь, в конце концов, систематика подобна гиперболе, и может показывать картину лишь примерно, из-за относительности понятий "родственность", "отличие", "характерная черта", и отсутствия конкретных предков. Мешает только то, что некоторые действительно делают поспешные выводы на фоне огромного числа работ, растущего уже полвека по экспоненте. -
Ну, а что такого резкого в открытии анкорацисты? То есть она, конечно, интересна сама по себе (о чем и говорится в статье по ссылке), но с точки зрения мегасистемы - всего лишь очередное микроцарство, каких известно уже штук пятнадцать. Как я понимаю, она даже не является базальным эукариотом, а примыкает к группе SAR (ох, как же мне не нравится это название!).
И... А почему Вы считаете понятие "родство" относительным?-
Недавнее обнаружение, потому вспомнилась; "хотя бы порядка" - поскольку филогенетически - хотя бы что-то. Действительно резких вещей не помню за последние года два. В систематическом плане - да, один из кучи неклассифицированных эукариот, где она и сидит; в тех же сотнях у них ныне находятся Telonemia.
Потому что есть объективное родство, а есть - субъективно воспринятое. Все базируется на субъективности, ведь из любых данных (не факт, что верно добытых) нужно сделать вывод; на нее и смотрим. В ее рамках - из-за, во-первых, суждения без возможности опереться на конкретного общего предка, лишь на нечто максимально близкое к обоим представителям из того, что удалось найти; во-вторых, сравнительный молекулярный анализ можно проводить от полной идентичности (= одна и та же особь) до бесконечности, и все сводится к тому, насколько дальними родственниками будут восприняты особи, стоит объединить каких-нибудь руконожек с лемурами, или они из-за максимальной удаленности от других лемуров будут удостоены чести обособиться в свой инфраотряд. Особо "плавная" дискретность, обусловленная постепенным расхождением организмов с каждым новым жизненным циклом на протяжении сотен миллионов лет, ну никак не позволяет сказать: "Эта особь принадлежит к этой группе, а та - к другой, и это объективный вывод, а не наша выдумка, призванная упорядочить море информации в соответствии с естественной потребностью человека". До каких пределов проводить разграничения? Они ведь призваны снизить "плавность", сделать разницу между особями более заметной. Классифицировать все от первичного эукариота до каждой отдельной особи не представляется возможным из-за умерших и необнаруженных особей, методологических ошибок, огромного объема данных; в то же время чем меньше ветвление - тем хуже показана "родственность". Кто-то в итоге выделяет SAR, кто-то - еще группы в рамках SAR.
Технически, для природы, родственность - вполне четкое понятие, но в рамках систематики, которая основана на человеческих возможностях, это до сих пор остается "пальцем в небо". Данные есть, но как их (статей), так и всей данной природой информации для изучения такие объемы, что в выводах, вместо объективных вещей, получается "мое видение" и всяческие предположения. На них систематика и базируется. Можно с пафосом говорить о существовании истины, к которой и стремимся, но приходится признать, что она просто недостижима. Кто-то оттого идет легким путем, и ударяется в мистику, кто-то - более благородным, но это, как ни ужасно, Ахиллес и черепаха. И опираться приходится на свои возможности, а не само наличие черепахи. Это все околофилософская ерунда, но куда без нее в данном вопросе.-
А что, Ахиллес действительно не может догнать черепаху? :)
Я понимаю, о чем Вы говорите, но в целом я сторонник реалистической позиции: "любая ветвь эволюционного древа так же реальна, как и любая ветвю любого дерева, ветвящегося в пространстве". Это древо может быть видно лучше или хуже, какие-то его участки от нас вообще пока закрыты, но так или иначе оно существует и оно единственное.-
Не стал указывать, что на практике - может, и вот подловили.
О том и речь, что древо реально, родство для природы реально, это легко разумеется, никто не говорит, будто объективной реальности вовсе нет, но какая разница, если по крайней мере n лет оно не будет доступно, лишь контур, как и сейчас. А до этих n лет человечество само себя разрушит, может; не фактически - так научно-культурно. Это не повод сдаваться, но не лучше ли отталкиваться от своих нынешних и ближайших возможностей вместо, абсолютизируя, допустимого "когда-нибудь переместимся и сами на два миллиарда лет назад - да на все взглянем".-
1. Не только "на практике", но и в том случае, если его движение описывается интегральным уравнением, а не аппроксимируется суммой конечных отрезков. По-моему, смысл парадокса с Ахиллом и черепахой в этом и был: автор смутно догадывался, что пора бы переходить к интегральному исчислению, но не знал как.
2. Есть разные мнения, в том числе и такое:
https://www.zin.ru/journals/trudyzin/doc/vol_317_s1/TZ_317_1_Supplement_Aleoshin.pdf
В любом случае, даже если филогенетическое древо будет установлено полностью, это будет только начальной ступенью изучения эволюции.-
Интегралы перпендикулярны Ахиллу. Тут конечное значение суммы бесконечного ряда. Попросту сумма убывающей геометрической прогрессии.
-
Насколько я понимаю, теория рядов появилась позже, чем исчисление бесконечно малых, и на его основе. Я в том смысле, что для понимания того, что парадокс на самом деле не парадокс, был нужен матан.
-
Нас учили такой идее, что производные придумали для того, чтобы считать интегралы. Правда на Вики написано, что обозначение опр. интегралла придумал только Фурье.
Методы с рядами придумал какие-то Коши, он же и с пределами работал (хотя как до него определяли производную - не знаю).
На Вики такое есть:
"школа Лейбница предпочитала в качестве базового понятия дифференциал".
-
-
-
-
-
-
-
-
Я когда то в интернете писал о том как после войны Германия стала великолепной страной и как там устроено. Довоенная социалдемократическая Германия было ужастиком. С нацизмом всё понятно. А вот после 1945-ого все остальные собрались и решили, как жить дальше. Получился синтез капитализма и христянской этики - социальная рыночная экономика. По сути экономисты изучали как лучше зарабатывать, а теологи, как - тратить. В народе тогда Гитлер все еще был поопулярен, и воровали много.
Но это не всё. В ГДР преподовалась теология. Недавно при Меркель президентом был Йоагим Глаук, который в ГДР из за плохого происхождение мог поступить только на теолога. А по сути то же управление людми.
У них там самыми велики немцами считается Аденауер, Лютер (религия) и Маркс (!). Вот и власти открыли дисертации по теологии. Наверное мои тексты на это повлияли;)
Я тут только любитель, но многие западные практики, как демократия или вожди харизматы, сначало были в религии. Сначало только реформатские церкви управлялись институциональной демократиией. А потом их приверженцы поплыли в Америке и устроили себе такую же власть. Наверное по этой природе вне запада демократия так плохо приживается. Не тратить деньги на дешёвые понты, а на благотворительность идёт с религиозных взглядов Северо Запада Европы.
-
Можно и ГМО анализировать таким же образом, теологически и социально.
ГМО страшно. Но например, весь инсулин произведён ГМО. И сколько жизней уже спас этот ГМО?
Я бы предпочёл "Бычее Сердце" а не фиолетовые ГМО помидоры со вкусом сахарина. Но вот если бы в СССР выращивали ГМО хлопок, которому нужно было бы меньше пестицидов, то сколько бы родилось больше здоровых детей?
И кажется, что то место, где можно увидеть ГМО, это ближайший цветочный магазин. А то в нём как то необычайно ярко всё сияет. К тому цветы на ГМО не проверяют и не запрещают, их то не жрут.
п.с. полный оффтоп. -
Отвечу на вопрос, комментировать не-вопрос не буду. Считай, наука это бренд. Т.е. есть метод (он хорошо описан, я не буду повторяться), и что методу не соответствует - это не наука. Наукой не являются теология, астрология, хиромантия и прочее - т.к. не проходят проверки по методу. Так вот теологи лезут в школу и навязывают свои теории параллельно науке и имитируя науку. Это подделка бренда. Хотите преподавать - делайте воскресные школы, отдельные от общеобразовательных. Иначе это сбивает с толку и нарушает брендирование. Это всё равно что прийти в храм и кричать что наука в теории существования бога не нуждается и начать спорить там в храме во время проповеди, приводя аргументы что землю создали не 5000 лет назад и указывая на подобные ляпы библии - согласитесь, верующие никакой терпимости тут на "своей" территории не проявят и самое меньшее, выставят такого проповедника от науки прочь. Так вот наука против того что кто-то лезет на её территорию.
PS: сама фраза "теория Дарвина" вместо "эволюционная биология" чаще всего выражает уничижительное отношение к науке, какбэ подразумевающее что наука находится на уровне Дарвина (19-й век) и что всё излагаемое в эволюционной биологии есть мнение Дарвина, а не научное направление, которое за 100 лет окрепло и обросло огромным количеством проверок и фактов, а также - успешных предсказаний и практически применимых технологий.-
-
Строго говоря, история будет не понятна не изучив историю религии. У них вроде не совсем что религия а как бы социология религии. Самый большой конфликт с биологами. У физиков проще, так не касается предмета изучения.
Методика эффективная. ИМХО я думаю, что про ГМО надо говорить в таком ключе. Народ верит в теории заговора, вредность не доказана, но восприятие тут скорее сердцем а не разумом и нужен эмоциональный аргумент. Но ГМО лекарство и спасает жизни. Как инсулин.
Дарвин на западе, особо среди англосаксов считается величайшим гением. Что так. Но в основном популяризирует как раз него, мало рассказывая про более новые открытия. Западная капиталистическая экономика основана на дарвинизме, зделали среду, разделили поставщиков и проивзодителей газа и конкурируйте. Но есть и менее жёсткие теории, где говорится о роли сотрудничества, кооперации, помощи в эволюции.
Устойчивые к антибиотикам микробы - бич больниц и рак всё эволюция, с которой практически сталкиваются, отрицать это глупо. Офф церковные вожды умные люди, так что не отрицают. Это скорее народная вера и секты.
-
-
-
11. Неправедный пусть еще делает неправду, нечистый пусть еще сквернится; праведный да творит правду еще, и святой да освящается еще.
12. Се, гряду скоро, и возмездие Мое со Мною, чтобы воздать каждому по делам его.
13. Я есмь Альфа и Омега, начало и конец, Первый и Последний.
14. Блаженны те, которые соблюдают заповеди Его, чтобы иметь им право на древо жизни и войти в город воротами.
15. А вне — псы и чародеи, и любодеи, и убийцы, и идолослужители, и всякий любящий и делающий неправду.
Апок. 22;11:15
2. Не дадите ли ссылку на побег пищи из тел Collodictyon? (не столько научный интерес, просто потрясно)
3. Публикации в инете, как и на бумаге, имеет много смыслов метить датой - по ссылке я ее так и не нашел :( хорошо, что тут Вы ее помянули.
-
1. Пробовали, конечно. Покамест основной подозреваемый на далёкого предка LECA, вроде, Локи. У него, в частности, уже началась характерная для эукариот дупликация многих генов, и, например, паралоги субъединиц протеасомы явно находятся на пути к эукариотным.
2. https://en.wikipedia.org/wiki/Collodictyon
Последние новости











Рис. 1. Представители «линий-сирот»: A — Ancyromonas (иллюстрация из Википедии), Б — Rigifila (иллюстрация из статьи A. Yabuki et al., 2013. Rigifila ramosa n. gen., n. sp., a filose apusozoan with a distinctive pellicle, is related to Micronuclearia), В — Mantamonas (иллюстрация из статьи E. Gluksman et al., 2011. The novel marine gliding zooflagellate genus Mantamonas (Mantamonadida ord. n.: Apusozoa)), Г — Collodictyon (иллюстрация с сайта dailymail.co.uk). Изображения в разных масштабах! На фото Б стрелки указывают на бактерий, захваченных длинными тонкими ветвящимися ложноножками ригифилы. На фото В стрелка указывает на передний жгутик