У предков билатерий уже была примитивная протохорда

Международная команда ученых взялась за изучение интересной анатомической структуры у кольчатых червей, на которую раньше внимания не обращали. Речь идет о брюшной продольной мышце кольчецов. Ученые присвоили этой структуре название аксохорд по аналогии с нотохордом (то есть хордой), характерным для хордовых (позвоночных). Это обязывающее название вполне оправдывает себя. Ведь, как выяснилось, аксохорд напоминает хорду по своей анатомической топографии, по своей генетической регуляции, по порядку развития из зародышевых тканей, а также функционально. По предположению ученых, подобная структура уже имелась у примитивных билатерий, а в ходе эволюции она видоизменилась, превратившись в аксохорд или в хорду.
Хорда (или нотохорд) — это осевой продольный стержень, имеющийся у всех (!) хордовых животных хотя бы на ранних стадиях развития. Это гибкое и прочное образование придает устойчивость форме животного, служит опорой при движении; хорда справляется с этой функцией надежнее, чем гидростатический мышечный мешок. У позвоночных животных хорда заменяется костным позвоночником, и тогда опора для тела становится еще основательнее. В небиологическом обиходе хорду знают по пирожкам с вязигой (вязига — хорда осетровых рыб; именно она и составляла основу начинки кулебяк и расстегаев). Для биологов хорда значит много больше, чем старинный кулинарный рецепт. Ясно, что ее формирование знаменует становление целого типа животных. Первые хордовые известны в палеонтологической летописи с начала кембрия (например, Haikouichthys). Как и откуда взялась хорда, этот наиважнейший элемент плана строения целого типа животных?
На этот вопрос ученые из Гейдельбергского университета, Европейского центра молекулярной биологии в Гейдельберге (Германия) и Исследовательского кампуса «Джанелия» в Ашберне (США) дали неожиданный ответ. Прообраз хорды (а точнее — топографически, генетически и функционально близкая к ней структура) имелся уже у общего предка всех двусторонне-симметричных животных Urbilateria (Ur — по-немецки означает исходный, bilateria — двустороннесимметричный, см. рис. 1). Это решение выглядит серьезно обоснованным: целый комплекс цитологических, эмбриологических и генетических признаков привлечен для его подтверждения.
Чтобы понять рассуждения ученых, нужно вспомнить две вещи. Во-первых, что билатерии разделяются на две группы: первичноротые и вторичноротые. К первичноротым относятся, в частности, членистоногие и кольчатые черви, а к вторичноротым, помимо прочих, хордовые. Во-вторых, как известно еще со времен Сент-Илера, план строения хордовых такой же, как и у первичноротых, только перевернутых с брюха на спину (см.: Д. А. Воронов. Старая гипотеза «перевернутости» хордовых подтверждается, «Природа» №11, 2000). У первичноротых на брюшной стороне расположены брюшная нервная цепочка, над ней — мезодермальные мышечные структуры, над ними — брюшной кровеносный сосуд и эктодермальная кишка; у хордовых все то же самое, только отсчитывая от спинной стороны: верхнее положение занимает нервный тяж, под ним хорда, под ним кровеносный сосуд и кишка. Это означает, что брюшная область мезодермы кольчатых червей гомологична спинной хондро-мезодермальной области хордовых, той, где расположена хорда. Поэтому, разобравшись в аналогиях развития осевых мезодермальных структур у представителей первичноротых и вторичноротых, можно реконструировать пути становления хорды.
Разобраться — значит выяснить, как формируются осевые мезодермальные клетки в раннем индивидуальном развитии первичноротых и какие гены участвуют в их трансформациях. Ученые обратились к анатомии многощетинкового червя Platynereis dumerilii (рис. 2).

Рис. 2. Полихета Platynereis dumerilii. Ее нетрудно выращивать в лабораторных условиях, поэтому это удобный объект для исследований. Например, как выяснилось не так давно, развитие и специализация продольных нервных стволов у этого червя организованы теми же генными регуляторами и по той же топографической схеме, что и в центральной нервной системе у позвоночных животных. Следовательно, у их общего предка могла быть центральная нервная система, которая могла развиваться по общей базовой схеме (A. S. Denes et al., 2007. Molecular architecture of annelid nerve cord supports common origin of nervous system centralization in bilateria). Фото с сайта dailymail.co.uk
У этого червя даже на ранних стадиях развития превосходно видны все требуемые осевые структуры (их можно увидеть на компьютерной модели). С помощью различных методов визуализации удалось различить особую группу клеток, которые формируют тонкий продольный мускульный тяж на брюшной стороне (продольная брюшная мышца, как ее называют). Эти клетки изначально появляются парами по бокам от средней линии зародыша, а по мере развития продвигаются друг к другу и сливаются в единую центральную структуру (рис. 3). Они располагаются над брюшной нервной цепочкой.

Рис. 3. Схематическое изображение развития позвоночных (рыбка данио) и аннелид (P. dumerilii). Зародыш Danio rerio изображен через 9 (стадия 90% эпиболии), 14 (стадия нейрального киля) и 30 (стадия органогенеза) часов после оплодотворения; зародыш P. dumerilii — через 32 часа, 72 часа и 2 месяца после оплодотворения. Толстая пунктирная линия показывает слияние нейроэктодермальных клеток с мезодермальными, у беспозвоночных это брюшная сторона (v), у позвоночных — спинная (d). Голубые стрелки — экспрессия Bmp — топографического организатора формы тела. Тонкие черные стрелки — направление соединения и растяжения клеток, тонкий пунктир — сечение зародыша, изображенное внизу (показаны белыми стрелками). Показаны красным — хорда (nch) и аксохорд (ach), оранжевым — мезодерма (mes), фиолетовым, серым и желтым — нейроэктодермальные структуры (neuroect), синим — эпидермис, зеленым — энтодерма (end) или кишка. Coe — целом, vao — брюшной кровеносный сосуд, vom — брюшные косые мышцы, vlm — брюшные боковые мышцы. Общие для двух групп транскрипционные факторы показаны соответствующими цветами. Схема из обсуждаемой статьи в Science
Затем были выявлены генетические маркеры, характерные для этих особых клеток. В них экспрессируются, как оказалось, гены, которые подключаются и при формировании хорды у хордовых (конкретно вот эта группа: brachyury, foxA, foxD, twist, not, soxD, soxE, noggin, hedgehog, netrin, slit). Все эти гены многофункциональны и работают в разных сочетаниях или по отдельности и в других клетках полихеты, но только в клетках центрального мускульного тяжа они экспрессируются все вместе — собираются, чтобы совместными усилиями организовать эту структуру.
Какова функция этой осевой структуры? Опять же, тонкое анатомирование показало, что сам тяж способен сокращаться; кроме того, к нему прикрепляются поперечные мускульные волокна (брюшных косых мышц, идущих к параподиям). Если с помощью лазера перерезать его, не задев сами мышечные волокна, то червяк теряет способность ползать, только неловко изгибается и ёрзает (видео с результатом операции). Это означает, что тяж участвует в координированном перемещении червя: служит опорой для мышечных волокон, участвующих в движении конечностей.
Авторы работы назвали этот осевой мышечный тяж аксохордом (от греч. άξονας ‘ось’ и χορδή ‘струна’). Название отражает сходство с нотохордом (от греч. νώτα ‘спина’ и χορδή ‘струна’): это сходство проявляется в топографии аксохорда, в порядке его формирования, в генетической регуляции его развития и, в известной степени, это функциональное сходство. Также это название указывает на различие — расположение на брюхе, а не на спине. Чтобы оправдать столь обязывающее название, ученые выявили подобные мезодермальные тяжи — аксохорды — и у других первичноротых (рис. 4). Как выяснилось, аксохорд имеется у других видов полихет, в том числе и у сидячих, а у личинок брахиопод и моллюсков имеется аналогичная по расположению осевая продольная мускульная структура. У щетинкочелюстных он присутствует в виде двух сомкнутых продольных мускульных тяжей, которые раздваиваются впереди; известен он и у полухордовых.

Это исследование показало, что осевая мезодермальная структура, так или иначе выполняющая функцию организатора движений, присутствует и у первичноротых, и у вторичноротых. Авторы предполагают, что общий предок всех билатерий Urbilateria уже был снабжен подобной структурой. О том, обладала ли она способностью к сокращению в исходном состоянии или нет, можно только гадать. Так или иначе, в ходе эволюции первичноротые преобразовали ее в аксохорд, членистоногие его впоследствии утеряли; у вторичноротых она трансформировалась в хорду; в группе вторичноротых от него избавились иглокожие. Прежние кандидаты на роль про-нотохорда — стомохорд или пигохорд полухордовых — явно проигрывают аксохорду. Для этих двух, помимо прочего, не соблюдается общая топография общего плана строения и работают другие генетические регуляторы, чем в хорде и аксохорде.
Аксохорд — «многообещающий» орган у первичноротых. Во-первых, имея его в виду, можно объяснить «внезапное» появление совершенно нового органа у хордовых животных: ведь хорда не объявилась вдруг из ниоткуда, для ее построения уже имелась солидная база. Преобразование первичной мезодермальной осевой структуры по ходу эволюции первичноротых и вторичноротых, и хордовых в их числе, станет новой и содержательной темой для дальнейших исследований. Во-вторых, вполне вероятно, что аксохорд имелся у ранних проблематичных животных кембрия и докембрия. Теперь стало понятно, на какие структуры древнейших билатерий стоит обратить более пристальное внимание.
Источник: A. Lauri et al. Development of the annelid axochord: Insights into notochord evolution // Science. 2014. V. 345, P. 1365–1368.
Елена Наймарк
-
"У первичноротых на брюшной стороне расположены брюшная нервная цепочка, над ней — мезодермальные мышечные структуры, над ними — брюшной кровеносный сосуд и эктодермальная кишка" - конечно, эктодермальная (передняя) кишка бывает, но все-таки под хордой кишка, наверное, энтодермальная (судя по подписям к рисункам, авторы статьи пишут о ней).
И вот еще что интересно - классическая эмбриология учит нас, что хорда у ланцетника образуется не из мезодермы, а из энтодермы - из крыши первичной кишки. Причем ее зачаток явно не составляет единого целого с зачатком мезодермы - хорда образуется спереди назад, а закладывается на переднем конце там, где сомитов нет. Обсуждают ли это авторы статьи?
Интересно также, пишут ли они что-нибудь про нематод и других экдизозоев (кроме насекомых)? Жалею, что сейчас у меня нет открытого доступа, и без обходных путей не могу посмотреть оригинал статьи...-
Не соглашусь. По-моему, классическая эмбриологии учит нас, что хорда - из мезодермы (так в большинстве учебников). Например, у лягушки, насколько я понимаю, зачаток хорды на начальных стадиях вполне составляет одно целое с зачатками парной мезодермы. Ну не может же быть, чтобы у ланцетника хорда развивалась из одного листка, а у лягушки из другого! Впрочем, мне кажется, что этот спор - стопроцентно схоластический, поскольку и сами понятия зародышевых листков - в общем, сугубо договорные.
-
"Ну не может же быть, чтобы у ланцетника хорда развивалась из одного листка, а у лягушки из другого!" - Еще как может! Вот у насекомых, например, так: "Однако у более примитивных щетинохвосток эктодерма не принимает участия в построении средней кишки, она образуется целиком за счет потомков вителлофагов. У многих, более совершенных насекомых, напротив, весь кишечник развивается из эктодермы. У паразитического перепончатокрылого Prestwichia он формируется из разросшегося стомодеума, а функции энтодермы ограничиваются присутствием в массе желтка постепенно дегенерирующих вителлофагов. Таким образом, в эволюционных преобразованиях эмбриогенезов насекомых происходит постепенное замещение энтодермы эктодермальными зачатками кишечника."
Что касается "схоластичности" - тут уже я не соглашусь. Речь идет не о зародышевых листках, а о том, откуда берется и как возникает хорда ланцетника - так, как описано авторами статьи для якобы всех хордовых, или иначе (из другой области зародыша и за счет других клеточных процессов).
-
-
-
Да, я тоже обратила на это внимание: на расхождение с тем, как нас учили. Но нигде не нашла упоминаний о другом варианте,энтодермальном, закладки хорды.
-
Иванова-Казас описывает развитие ланцетника со ссылками на работы Conklin и других авторов. Хорда отшнуровывается от крыши первичной кишки (отдельно от мезодермы), причем примерно так же, как нервная трубка от эктодермы. Клетки ее сначала похожи на энтодермальные, так как содержат желток. Сходно с другими хордовыми то, что происходит "вклинивание" рядов клеток при их продвижении к средней линии.
Любопытно также, что у асцидий зачатки хорды и мезодермы практически полностью изолированы друг от друга (на карте зачатков) и в яйце , и на стадии 8 клеток. Те бластомеры, что содержат локальные детерминанты хорды, дают начало энтодерме, но не мезодерме. Для индукции хорды у асцидий, насколько я понимаю, необходимо ее взаимодействие с энтодермой и нейральной эктодермой, но не с мезодермой...
-


Эволюция хордовых
-
 16.09.2024Открыто новое кембрийское позвоночноеСергей Ястребов • Новости науки
16.09.2024Открыто новое кембрийское позвоночноеСергей Ястребов • Новости науки -
 05.11.2015Гомологи хорды широко распространены у первичноротыхСергей Ястребов • Новости науки
05.11.2015Гомологи хорды широко распространены у первичноротыхСергей Ястребов • Новости науки
-
 18.03.2015Общий предок вторичноротых мог быть похож на хордовоеСергей Ястребов • Новости науки
18.03.2015Общий предок вторичноротых мог быть похож на хордовоеСергей Ястребов • Новости науки
-
 04.03.2015У ланцетника тоже есть хрящЕлена Наймарк • Новости науки
04.03.2015У ланцетника тоже есть хрящЕлена Наймарк • Новости науки
-
 04.02.2015«Четвертый зародышевый листок» позвоночных зародился у низших хордовыхСергей Ястребов • Новости науки
04.02.2015«Четвертый зародышевый листок» позвоночных зародился у низших хордовыхСергей Ястребов • Новости науки
-
 18.09.2014У предков билатерий уже была примитивная протохордаЕлена Наймарк • Новости науки
18.09.2014У предков билатерий уже была примитивная протохордаЕлена Наймарк • Новости науки
-
 25.10.2013У одноклеточных организмов есть ген, способный управлять развитием хордыСергей Ястребов • Новости науки
25.10.2013У одноклеточных организмов есть ген, способный управлять развитием хордыСергей Ястребов • Новости науки
-
 17.06.2013Своим сердцем позвоночные обязаны полногеномной дупликацииСергей Ястребов • Новости науки
17.06.2013Своим сердцем позвоночные обязаны полногеномной дупликацииСергей Ястребов • Новости науки
-
 16.03.2012Пикайя — самое примитивное хордовое?Александр Марков • Новости науки
16.03.2012Пикайя — самое примитивное хордовое?Александр Марков • Новости науки
-
 23.06.2008Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночныхАлександр Марков • Новости науки
23.06.2008Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночныхАлександр Марков • Новости науки
Последние новости











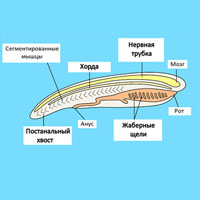
Рис. 1. Так мог выглядеть общий предок двусторонне-симметричных животных Urbilateria. Его сегментированное тело развивалось из трех зародышевых слоев, в нем различались перед и зад, на переднем конце располагался рот и чувствительные фоторецепторы и антенны, что предполагало наличие небольшого нервного центра, также имелись осевая кишка и сердце с кровеносной системой. Выясняется, что у них, возможно, имелась и осевая структура на брюхе, гомологичная хорде. Рисунок с сайта palaeos.com