«О, сколько нам открытий чудных...», или Роль случая в палеонтологии
Владимир Комаров, Екатерина Шпилевая, Юлия Вайтиева
«Природа» №12, 2020
 Владимир Николаевич Комаров — кандидат геолого-минералогических наук, доцент кафедры палеонтологии и региональной геологии Российского государственного геологоразведочного университета имени Серго Орджоникидзе (МГРИ). Область научных интересов — палеонтология и стратиграфия. Постоянный автор «Природы». |
 Екатерина Сергеевна Шпилевая — студентка первого курса геологоразведочного факультета того же университета. Специальность — прикладная геология. |
 Юлия Алексеевна Вайтиева — студентка второго курса того же факультета. Специальность — прикладная геохимия, петрология и минералогия. |
В начале известной в прошлом телевизионной передачи «Очевидное — невероятное» каждый раз звучали замечательные стихи А. С. Пушкина: «О, сколько нам открытий чудных / Готовят просвещенья дух / И опыт, сын ошибок трудных, / И гений, парадоксов друг». Но в этом стихотворении есть еще одна, заключительная, строчка, которую в передаче зрители не слышали: «И случай, Бог изобретатель». Роль случая огромна и часто имеет решающее значение в самых разных областях, в том числе и в палеонтологии.
В сентябре 2020 г. один из авторов данной статьи внимательно пересматривал учебные коллекции окаменелостей, из богатейшего запасного фонда кафедры палеонтологии и региональной геологии Российского государственного геологоразведочного университета имени Серго Орджоникидзе (МГРИ). Это было необходимо в связи с возможной подготовкой к дистанционному формату обучения из-за вредоносного коронавирусного поветрия. И вдруг взгляд остановился на уникальном по сохранности образце фрагмента раковины белемнита (Belemnoidea) (рис. 1), который до этого, скорее всего, никогда не использовался в учебном процессе. Находка в полной мере подтверждает, что для сокращения неполноты геологической летописи крайне важно повторно изучать объекты, в частности периодически пересматривать с современных позиций старые палеонтологические коллекции [1, 2]. Геологический возраст и автор находки, а также ее местонахождение, к сожалению, неизвестны. Однако даже без учета этих данных обнаруженный образец имеет огромное методическое и познавательное значение.

Строение белемнитов

Рис. 2. Ростры белемнитов
К белемнитам относятся вымершие колеоидеи с опорным внутренним скелетом, состоящим из трех частей — фрагмокона, проостракума и ростра. В ископаемом состоянии обычно сохраняется ростр, как наиболее прочный элемент скелета (рис. 2). Фрагмокон встречается значительно реже. Собственный опыт, основанный на изучении верхнеюрских белемнитов из зоны Virgatites virgatus в окрестностях пос. Нижние Мнёвники (Москва) показал, что на 30 ростров приходится одно ядро фрагмента фрагмокона [3]. Проостракум, как наиболее хрупкий элемент скелета, сохраняется в исключительных случаях.
Фрагмокон — камерная часть скелета белемнитов (рис. 3). Это арагонитовый прямой или слегка изогнутый к брюшной стороне конус. Состоит он из камер, которые разделены поперечными перегородками (септами), вогнутыми назад. В вершине альвеолярного конуса расположена первичная шарообразная или полусферическая камера, называемая эмбриональной. Фрагмокон окружен тонкой оболочкой — конотекой. Вдоль брюшного края сквозь перегородки проходит сифон, дудки которого направлены назад. Задняя часть фрагмокона располагается в альвеоле — коническом углублении на переднем конце ростра. Фрагмокон белемнитов соответствует сумме всех камер (за исключением жилой) остальных подклассов головоногих.

Рис. 3. Схема строения скелета белемнитов [4]: а — продольный разрез, б — реконструкция скелета, в — продольный раскол; 1 — проостракум, 2 — ростр, 3 — сифон, 4 — камеры, 5 — начальная камера, 6 — перегородки, 7 — альвеола, 8 — апикальная линия
Проостракум представляет собой широкую языковидную тонкую пластинку. Традиционно под проостракумом понималось только продолжение спинной стороны конотеки фрагмокона. Но последние электронно-микроскопические исследования выявили наличие тончайшего слоя проостракума, перекрывающего весь фрагмокон.
Ростр — как правило, массивное образование, форма которого определяется соотношением спинно-брюшного и бокового диаметров на различных участках. Форма ростров может быть цилиндрической, конической, веретеновидной, ланцетовидной или сигаровидной. В передней (альвеолярной), расширенной, части ростра помещается альвеола. Угол, составленный образующими альвеолярного конуса, называется альвеолярным. Он отличается большим постоянством: не изменяется с возрастом и одинаковый у определенных родов и даже видов. Задняя часть (противоположный, обычно суженный, конец ростра) — это вершина, или вершинный конец. Острие располагается центрально или эксцентрично, т.е. приближается к спинной или брюшной стороне. Участок ростра от заднего конца до начала альвеолы называется постальвеолярной частью. Как уже отмечалось, у ростра различают брюшную сторону (к которой близко подходит сифон), спинную и две боковые. На брюшной стороне встречаются хорошо выраженные брюшная борозда и боковые бороздки, которые служат надежным систематическим признаком. Реже наблюдается спинная борозда. В передней части брюшная сторона иногда прорезана так называемой брюшной, или альвеолярной, щелью. На поверхности ростра наблюдаются отпечатки кровеносных сосудов. Форма поперечного сечения остается постоянной вдоль всей длины ростра или изменяется в различных его частях. Поперечное сечение — круглое, овальное, реже угловатое или многоугольное. Внутренние признаки ростра можно наблюдать в плоскости спинно-брюшного раскола, где очень хорошо видны линии нарастания, которые отражают изменение формы ростра в онтогенезе, а также осевая (апикальная) линия. На продольном расколе ростра (в плоскости симметрии вдоль брюшной стороны) рядом с альвеолой видна спайка — гладкая поверхность, отличающаяся от остальной части скола. Размеры ростров значительно варьируют.
На сегодняшний день единая точка зрения на первичный минеральный состав ростров не выработана. Одни исследователи [5] полагают, что они были первично арагонитовыми, а впоследствии частично или полностью раскристаллизовались в кальцит, другие же считают ростры первично кальцитовыми [6]. Судя по данным детального изучения мегатеутидид, ростры — первично массивные (непористые) образования, которые имеют смешанный кальцит-арагонитовый состав с закономерной локализацией обеих минеральных фаз [7].
Белемноидеи, по всей видимости, появились в девонском периоде и вначале были немногочисленны. Их бурное развитие начинается с триаса. В юрский и меловой периоды представители отряда Belemnitida имели широкое распространение в любых палеозоогеографических областях. В конце мела произошло их вымирание.
Уникальный образец

Рис. 4. Скелет белемнита во вмещающей породе. Продольная пришлифовка. Здесь и далее фото В. Н. Комарова
Обнаруженный скелет белемнита мы изучали на продольном сечении (рис. 4). Он находится в пелитоморфном известняке черного цвета с характерным раковистым изломом. Окаменелость представляет собой довольно крупный фрагмент фрагмокона, в передней части местами разрушенный и деформированный, с фрагментом ростра. С учетом того, что начало альвеолы располагается по центру ростра, можно считать, что мы имеем дело не со спинно-брюшным сечением, а с боковым (латеральным), на котором невозможно увидеть сифон. Сохранившаяся часть ростра длиной 3,7 см характеризуется выдержанной цилиндрической формой. Спинно-брюшной диаметр в сечении у начала альвеолы определить не удалось, так как здесь ростр скрыт породой. На расстоянии 1,2 см от начала альвеолы в сторону вершины (что отвечает длине сохранившейся постальвеолярной части) его радиус составляет 0,9 см (диаметр, соответственно, 1,8 см). Альвеолярный угол в вентральном ракурсе равен 32°.
В пределах альвеолы длиной 2,5 см насчитывается 13 септ. В неразрушенной (1,4 см) части, выходящей за пределы альвеолы, — девять септ (рис. 5). Всего во фрагмоконе (общая сохранившаяся длина которого около 10,0 см) не менее 41 септы (точно подсчитать трудно, поскольку в деформированной части септы сближены, поломаны и искривлены). С возрастом животного септы более разряжены. В основании фрагмокона длина камер равна 1,5 мм, у внешней границы альвеолы — 2,4 мм (относительная длина камер здесь составляет 0,19 диаметра фрагмокона), а у внешней части целого фрагмента фрагмокона — 4,5 мм (относительная длина камер достигает 0,21 диаметра фрагмокона). Толщина септ чуть менее 0,1 мм, она немного увеличивается к краям. Начальная часть фрагмокона шарообразная, диаметром (насколько позволяет судить сохранность) 0,7 мм. Стенка местами хорошо сохранилась вне ростра и отчетливо обнажена в породе.

Рис. 5. Фрагмокон белемнита: а — задняя, находящаяся в альвеоле часть, б — неразрушенная часть, располагающаяся сразу за пределами альвеолы. Камеры фрагмокона заполнены пестроцветным вторичным кальцитом. Бинокулярный микроскоп МБС-1
Недеформированная часть фрагмокона заполнена кальцитом разных генераций (см. рис. 5). На стенках септ наблюдается инкрустация, выполненная крупнозернистым кальцитом пестрой окраски — белой, желтоватой, коричневатой, черной. Обычно он полностью заполняет камеры фрагмокона. Иногда в центральной части камер встречается вторичный мелкокристаллический кальцит светло-серого цвета. На его долю может приходиться до трети их объема.
Проведенное исследование показало, что фрагмокон и порода, заполняющая его пустотное пространство, пиритизированы, причем интенсивность пиритизации резко различна. Ею совершенно не затронута целая, недеформированная, часть фрагмокона, в которой камеры заполнены гетерогенным кальцитом. В пределах поломанного сектора порода интенсивно, местами тотально, пиритизирована, а фрагменты скелета с сохранившимся первичным карбонатным составом чередуются с полностью пиритизированными фрагментами септ и стенками фрагмокона (рис. 6). В породе пирит формирует мелкозернистую, иногда сплошную массу, в которой, используя бинокулярный микроскоп МБС-1, невозможно даже различить отдельные грани кристаллов. На фоне мелкозернистого пирита в виде одного небольшого локального очага мы отметили агрегат крупнокристаллического (до 2,5 мм) пирита (см. рис. 6).

Рис. 6. Пиритизация фрагмокона: а — агрегат крупнокристаллического пирита в поломанной части, б — поломанные, частично пиритизированные перегородки на фоне сплошной массы мелкозернистого пирита. Бинокулярный микроскоп МБС-1
Полное отсутствие пирита во вмещающей породе и наличие его внутри фрагмокона (рис. 7), на наш взгляд, безусловно, свидетельствует о его возникновении благодаря биологически индуцированному процессу. Доказательством этому служат многочисленные находки ростров с характерной пиритизацией в области фрагмокона. Вероятно, пирит образовался в осадке на ранних стадиях диагенетического перераспределения вещества в восстановительных геохимических условиях, ниже границы раздела осадка с поверхностными водами, в местах разложения органического вещества. Сульфатредуцирующие бактерии, восстанавливая сульфаты морской воды, используют кислород молекулы CaSO4 при окислении органического вещества. Для извлечения кислорода из этой химически очень прочной молекулы необходимо затратить значительную энергию, которую бактерии получают, усваивая органическое вещество. Вовлечение сульфатов в жизненный цикл бактерий сопровождается изменением валентности серы. Положительная шестивалентная сера в молекуле сульфата переходит в отрицательную двухвалентную. Такие реакции называют окислительно-восстановительными: S6+ → S2−. Двухвалентный ион серы взаимодействует с ионом водорода (H+), который в результате реакции диссоциации всегда присутствует в природных водах, образуя биогенный сероводород (H2S). Последний реагирует с ионами растворенного двухвалентного железа (Fe2+), связанного с поровыми водами окружающего осадка. Это приводит к возникновению моносульфида железа (FeS · nH2O) — коллоидного минерального образования черного цвета, кристаллизация которого через ряд промежуточных стадий завершается формированием пирита*. Считается, что органические соединения, растворенные в воде, увеличивают вязкость среды и обеспечивают осаждение более значительных концентраций тонких сульфидов, чем это возможно в чистой воде.

Рис. 7. Ростры белемнитов, пиритизированные в области фрагмокона
Историю формирования нашей окаменелости, с некоторой долей условности, можно представить следующим образом. Погибший белемнит оказался на дне достаточно глубоководного морского бассейна. В пользу относительной глубоководности свидетельствуют, на наш взгляд, полное отсутствие в породе ископаемых остатков донных организмов, а также пиритизация, которая указывает на дефицит свободного кислорода. Со временем, в процессе или после уничтожения почти всего мягкого тела, проостракум и внешняя часть фрагмокона разрушились. Оставшийся фрагмент хрупкого фрагмокона с внешней стороны подвергся деструкции. Поломка некоторых скелетных структур позволила осадку отложиться внутри фрагмокона. В заднюю же, не затронутую деформацией часть осадок попасть не смог. Органическое вещество, сохранившееся в открытом поломанном и заполненном (возможно, частично) осадком фрагменте, способствовало образованию здесь в восстановительных геохимических условиях пирита. Значительно позднее (вероятно, уже на стадии диагенеза) задняя, законсервированная часть фрагмокона в два этапа вся заполнилась вторичным кальцитом.
Реконструкция размеров раковины белемнита
Перед тем как привести данные о выполненных реконструкциях размера важнейших элементов раковины нашей окаменелости, хотелось бы отметить, что на большинстве изученных нами моделей скелета белемнитов фрагмокон показан выходящим за пределы ростра. Тем не менее на отдельных реконструкциях (в том числе, приведенных в учебной литературе) размеры фрагмокона ограничиваются размерами альвеолы, что может создать у читателей упрощенное представление о строении раковины белемнитов. Следует также подчеркнуть, что при определении размера отдельных частей мягкого тела мы использовали некоторые пропорции, характерные для современных кальмаров (рис. 8): форма тела белемнитов (об этом можно судить по немногочисленным отпечаткам их мягкого тела) и их образ жизни, видимо, приближались к таковым у кальмаров.

Рис. 8. Современный кальмар. Фото с сайта behance.net
Изучив многочисленные реконструкции, касающиеся соотношения длины отдельных частей ростра, и отбросив крайние цифры, мы вычислили среднее значение, которое показывает, во сколько раз постальвеолярная часть ростра длиннее альвеолярной. Оно оказалось равным 2,07. У нашего образца длина альвеолы составляет 2,5 см. Используя полученный коэффициент, мы определили гипотетическую длину постальвеолярной части ростра в 5,2 см, а его общую длину — в 7,7 см.
Соотношение длины выступающей над ростром части фрагмокона и длины самого ростра также приводится в литературе по-разному, причем примерно в равных пропорциях это касается как данных, отраженных просто на реконструкциях скелета белемнитов, так и данных, полученных на реконструкциях раковины, которая располагалась внутри мягкого тела. Средние значения этого показателя меняются от 0,66 (ростр длиннее выступающей над ним части фрагмокона) до 2,08 (ростр короче выступающей над ним части фрагмокона). У нашего образца данное соотношение составляет 0,97. Однако, учитывая то, что мы, безусловно, имеем дело далеко не с полным фрагмоконом, а лишь с его фрагментом, в дальнейших расчетах использовался коэффициент 2,08. При таких условиях длина выступающей над ростром части фрагмокона оказалась равной 16,01 см.
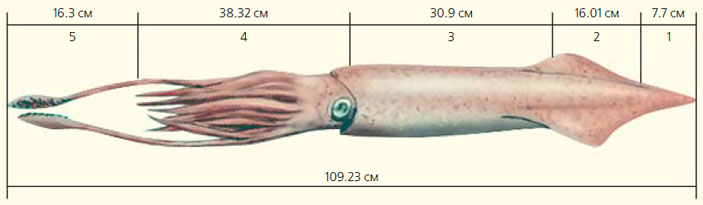
Рис. 9. Реконструкция размера белемнита на основе изученной окаменелости. 1 — ростр, 2 — выступающая из ростра часть фрагмокона, 3 — проостракум, 4 — голова и щупальцы, 5 — ловчие руки
Анализ литературных данных показал, что в среднем длина проостракума превышает длину выступающей над ростром части фрагмокона в 1,93 раза. Обычно на реконструкциях, отображающих положение скелета внутри мягкого тела головоногого моллюска, проостракум достигает головы животного. Если исходить из этого, мы получаем длину проостракума, равную 30,9 см, а длину от вершины ростра до головы белемнита — 54,61 см.
Отношение длины головы моллюска вместе со щупальцами к длине проостракума на подавляющем количестве реконструкций примерно одинаковое. В среднем этот показатель составляет 1,24. Путем несложных вычислений получаем длину головы вместе с основными щупальцами равную 38,32 см. Общая длина реконструируемого моллюска, таким образом, составит 92,93 см. А если учесть и длину пары ловчих рук, то размер окажется еще больше. Массовые замеры, проведенные нами с использованием опубликованных источников, показали, что длина головы современных кальмаров вместе с основными щупальцами превышает длину выступающих за их пределы ловчих рук в среднем в 2,35 раза. Таким образом, с учетом добавления еще 16,3 см (выступающая часть ловчих рук) общая длина нашего моллюска составит 109,23 см. Безусловно, все проведенные выше расчеты выполнены с известной долей условности.
Литература
1. Комаров В. Н. О неполноте геологической летописи. Статья 2. Об успехах в сокращении неполноты и не только // Известия вузов. Геология и разведка. 2010; 4: 3–9.
2. Шпилевая Е. С., Вайтиева Ю. А., Комаров В. Н. Об уникальном образце (Mollusca, Cephalopoda) в учебной коллекции кафедры палеонтологии и региональной геологии МГРИ // Инновации. Наука. Образование. Электронное периодическое издание. 2020; 20: 728–742.
3. Лукашенко С. В., Наянова Т. Ф., Комаров В. Н. Местонахождение ископаемых остатков зоны Virgatites virgatus в Нижних Мнёвниках (г. Москва) — уходящая натура // Известия вузов. Геология и разведка. 2009; 1: 3–7.
4. Бондаренко О. Б., Михайлова И. А. Краткий определитель ископаемых беспозвоночных. М., 1984; 354.
5. Dauphin Y., Williams C. T., Barskov I. S. Aragonitic rostra of the Turonian belemnitid Goniocamax: Arguments from diagenesis // Acta Palaeontol. Polon. 2007; 52(1): 85–97.
6. Stevens K., Griesshaber E., Schmahl W. et al. Belemnite biomineralization, development, and geochemistry: The complex rostrum of Neohibolites minimus // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017; 468: 388–402.
7. Ипполитов А. П., Барсков И. С., Косоруков В. Л., Десаи Бх. Микроструктурная и минералогическая архитектура ростров некоторых батских мегатеутидид (Belemnitida) // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция, экология и биостратиграфия. Материалы совещания. ПИН РАН. 2018; 5: 94–99.
* Кизильштейн Л. Я. Сульфатвосстанавливающие бактерии: великие дела маленьких существ // Природа. 2016. № 2. С. 51–53.

















Рис. 1. Реконструкция белемнита