Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Меловые муравьи Бирмы: что с ними не так?
Из бирманского янтаря возрастом около 99 млн лет описан 31 вид муравьев трех вымерших подсемейств (Haidomyrmecinae, Zigrasimeciinae, Sphecomyrminae). Эти муравьи имеют уникальные, не имеющие аналогов среди современных муравьев, особенности челюстей и наличника. Особенности головы и ротового «вооружения» приводили авторов описаний к выводу о своеобразном способе питания: специализированное хищничество, фильтрация жидкости, питание гемолимфой, капканный тип работы челюстей. Предлагаю гипотезу, согласно которой удивительное разнообразие и вымирание этих муравьев является следствием двух взаимосвязанных причин: особенностей морфологии, отличающих их от представителей современных подсемейств, и организации меловых сообществ муравьев, отличной от современного устройства мирмекокомплексов из-за отсутствия развитой внутрисемейной коммуникации. Представляется, что некоторые морфологические черты муравьев вымерших таксонов позволяют говорить об отсутствии у них эффективной коммуникации и коллективной охоты. Появление комплекса ключевых адаптаций у современных (кроновых) групп муравьев позволило им выйти за границы ниш специализированных хищников, поскольку при развитой коммуникации тип и размер жертвы не ограничен размерами муравья и его челюстей, использовать новые источники пищи (жидкие углеводы — нектар и падь) и обслуживать большие по численности семьи.
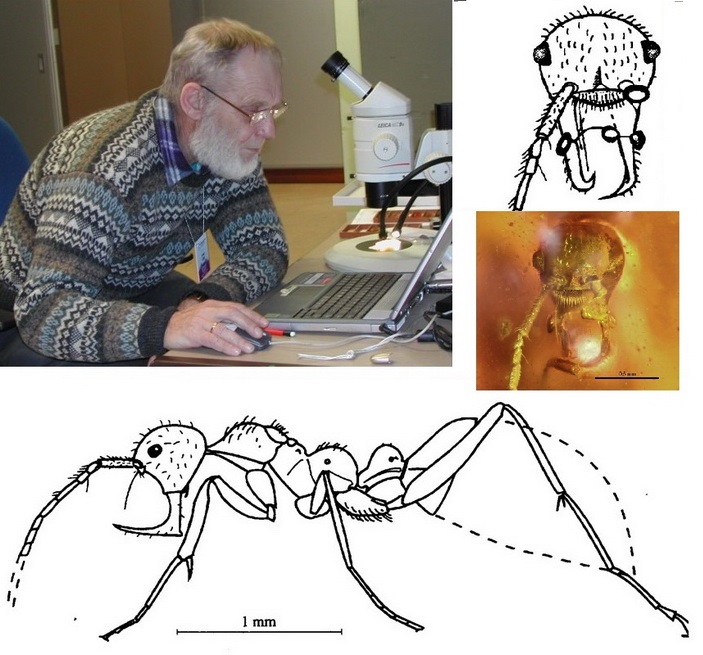
В 1995 году на кафедре теории эволюции и проблем дарвинизма (сейчас кафедра биологической эволюции) биологического факультета МГУ имени М. В. Ломоносова произошло, не поддамся ложной скромности, открытие мирового уровня. Профессор мирмеколог Геннадий Михайлович Длусский описал первого муравья из среднемелового бирманского янтаря (Мьянма, 99 млн лет; рис. 1). В то время всех меловых находок муравьев было всего-то несколько, среди них открытые самим же Длусским переходные формы между сколиоидами (от которых, как считается, произошли муравьи) и муравьями — самки арманииды (арманиины). Первого бирманского муравья заметил в коллекции Лондонского Музея естественной истории другой выдающийся российский (советский) ученый — Александр Павлович Расницын из лаборатории артропод Палеонтологического института имени А. А. Борисяка и обратил внимание Длусского на это примечательное насекомое. Муравей, помимо своей древности и отличной формы сохранности — янтарь, обладал совершенно замечательной внешностью: имел уникальные гигантские L-образные челюсти (мандибулы) и вытянутую в вертикальном направлении голову (см. рис. 1). Такие особенности отличали его от всех современных муравьев, хотя все другие части тела и весь облик насекомого однозначно говорили о том, что это муравей. Не чуждый иронии профессор назвал уникума «Муравей-цербер из Аида» — Haidomyrmex cerberus Dlussky, 1996 (Haidos греч. мифол. — Аид и myrmica греч. — муравей). Так, с легкой руки профессора, весь мир начал изучать «адских муравьев» (hell ants). Однако русскоязычной аудитории интересно будет узнать, что для самого профессора-первоописателя это был вовсе не адский муравей, а «Ай да муравей!» (Haido myrmex, айдамирмекс).
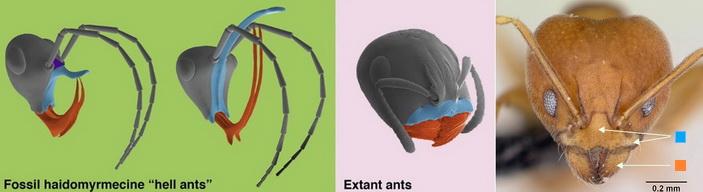
С момента описания первого представителя муравьев из бирманского янтаря прошло четверть века. За это время в бирмите обнаружили богатую фауну примитивных муравьев, представленную крылатыми и рабочими особями 31 вида трех вымерших подсемейств — Haidomyrmecinae, Zigrasimeciinae, Sphecomyrminae. Из бирмита этого возраста пока не описали муравьев из современных подсемейств, что говорит о преобладании, по крайней мере численном, в меловых биоценозах этого региона представителей стволовых групп муравьев, т.е. вымерших и не оставивших прямых потомков. Муравьи Sphecomyrminae, Haidomyrmecinae и Zigrasimeciinae имеют уникальные, не имеющие аналогов среди современных муравьев, особенности мандибул и, по крайней мере, для двух последних подсемейств наличника (клипеуса) и, по-видимому, не перешли мел-палеогеновую границу (рис. 2).
Условия возникновения специфического морфологического разнообразия и причины вымирания этих муравьев на данный момент не имеют удовлетворительного объяснения, несмотря на то, что обсуждаются почти в каждой публикации с описанием новых видов муравьев из бирманского янтаря. Целью моего исследования явился обзор и анализ причин специфической морфологической радиации стволовых таксонов бирманского янтаря и их вымирания на основании материалов, опубликованных за время с момента описания первого представителя.

Не вдаваясь в подробности, следует, наверное, пояснить, кто такие муравьи, как их выделить среди многообразия насекомых во всех исторических периодах. Внешне это бескрылые (кроме самок и самцов) насекомые, имеющие характерное расчленение тела — голову, грудь, брюшко и обязательно еще один (или два) «промежуточный» членик между грудью и брюшком, так называемый узелок (петиоль), узко причленяющийся к брюшку. Мелового представителя вымершей ветви, например Sphecomyrma freyi (американский меловой янтарь), не всякий энтомолог сможет отличить от современного муравья (рис. 3). Но уникальный внешний вид муравьев бирмита из других подсемейств, Haidomyrmecinae и Zigrasimeciinae, удивляет и привлекает внимание не только специалистов-энтомологов. У читателей популярных ресурсов по биологии и палеонтологии пользуются популярностью регулярные заметки об этих примечательных насекомых (читайте на страницах «Элементов» заметки Александра Храмова «Адские муравьи», «Охота адского муравья», Древний «тараканомуравей»). В бирманском янтаре кроме 14 видов айдамирмексов (адских муравьев) с гигантскими L-образными мандибулами, обнаружили 5 видов «муравьев — железных дев» (iron maiden ants, Zigrasimeciinae). Последние тоже весьма экзотической внешности: плоские мандибулы и наличник усеяны плотным частоколом хитиновых зубцов и щетинок (рис. 3).
Кажется удивительным тот факт, что сосуществовали очень похожие на современных муравьев представители стволовых групп типа Sphecomyrma freyi (в бирманском янтаре описаны 12 видов сфекомирмин из родов Gerontoformica и Myanmyrma) и в большом разнообразии экзотические адские муравьи и муравьи — железные девы (см. рис. 3). Морфологические исследования ротового вооружения айдамирмексов показали, что большие L-образно загнутые мандибулы и уникальный увеличенный наличник (названный рогом) возникали в группе адских муравьев неоднократно и параллельно (см. рис. 2). Вопрос, почему мир меловых муравьев был таким странным, очень волнует ученых уже четверть века и пока не имеет убедительного ответа. Энтомологи-исследователи мирмекофауны бирмита предположили, что это результат специализации муравьев по определенному типу добычи, а также экзотических способов охоты этих муравьев. Длусский еще при первом описании муравья-цербера предположил, что его мандибулы действовали по принципу капканных челюстей некоторых современных муравьев (например, Myrmoteras, Daceton; рис. 4). Основные аргументы в пользу этого это внушительные размеры мандибул и характерный атрибут челюстей-капканов современных муравьев — сигнальные волоски (trigger hairs, см. рис. 4).

Барден с коллегами полагает, что муравей-вампир Linguamyrmex vladi питался гемолимфой беспозвоночных, пронзая длинными челюстями тело жертвы и направляя при помощи желоба, образованного сомкнутыми мандибулами, гемолимфу к максиллам и затем в рот (отсюда название — Дракула Влад Цепеш). Удивительное устройство околоротового пространства довольно мелких муравьев — железных дев, длина тела которых 2-4 мм, приводило авторов к заключению о возможном использовании своеобразной «хитиновой щетки» для фильтрации неких жидкостей для питания, однако другие авторы описания видов муравьев — железных дев предполагали, что все-таки они охотились, как простые муравьи, на мелких беспозвоночных, не объясняя, зачем им понадобилась эта щетина. В этом случае исследователи верно (на мой взгляд) подметили, что мандибулы и наличник работают наподобие верхней и нижней челюстей позвоночных.

Находка адского муравья с добычей, зажатой между рогом и мандибулами, показала (и подтвердила мои догадки) еще один способ использования мандибул айдамирмексами, а также возможность движений их челюстей в вертикальной плоскости (см. заметку на «Элементах» «Охота адского муравья»). Совсем недавно обнаружены современные перепончатокрылые со сходной способностью к вертикально-латеральным движениям челюстей (Tyrannoscelio, Stentorceps, Nanocthulhu, Colotrechninae sp.) и внешним сходством формы головы и мандибул с адскими муравьями, подтвердившие реализуемость таких перестроек головной капсулы и кранио-мандибулярной системы (рис. 5).

Строение антенн — другая важная черта меловых муравьев, отличающая их от современных «настоящих» муравьев. Дело в том, что антенны современных муравьев имеют ряд особенностей (рис. 6): во-первых, они, так называемые, коленчатые: первый членик антенн (скапус) является самым длинным — его длина составляет не менее трети от всей длины антенны, за ним второй членик (педицель) и цепочка из маленьких члеников (жгутик). Таким образом, антенна имеет «колено». Во-вторых, второй членик антенн (педицель) относительно длинный и изогнут, так что позволяет жгутику плотно прилегать к скапусу. В-третьих, вершинные членики антенны крупнее срединных. Четвертая особенность — так называемый латеральный поворот, когда усиковые желобки ограничивают движения антенн преимущественно боковой плоскостью. Антенны же вымерших меловых муравьев совсем другие: скапус бывает разной длинны и у подавляющего большинства видов не достигает и четверти длины всей антенны, педицель прямой и короткий, а жгутик антенны «нитчатый» — длинный с заостренным вершинным члеником (см. рис. 6).
В свое время Длусский и Федосеева (1988) предположили, что строение антенн (короткий скапус) и двузубые мандибулы вымерших меловых муравьев свидетельствуют об отсутствии «истинной социальности, для которой характерны тонкие манипуляции с расплодом» и непригодны для построения гнезда. Тем не менее за прошедшее время вполне убедительно показано, что меловые муравьи вели эусоциальный образ жизни. Барден и Грималди (2016) в своем исследовании резюмировали веские морфологические доказательства эусоциальности меловых муравьев: наличие по крайней мере у некоторых из них бескрылой и крылатой каст, при этом имеются репродуктивные самки со следами отброшенных крыльев. Также они предположили, что косвенным свидетельством семейственности и наличия группового поведения является наличие нескольких довольно редких в меловых ориктоценозах муравьев в одном куске смолы. Совместное нахождение крылатых самок и рабочих особей Zigrasimecia ferox разного размера дали повод другим исследователям говорить о начальных этапах формирования у меловых муравьев примитивного полиморфизма — размерного — на основе изометрического роста. Однако связь строения мандибул и антенн с социальностью, которую предположили Длусский и Федосеева, другими исследователями не рассматривалась, за исключением работы Борисенко (Borysenko, 2017), в которой он статистически подтвердил предположения этих авторов о морфометрических особенностях антенн современных муравьев в сравнении с другими социальными и одиночными перепончатокрылыми, а также с вымершими меловыми муравьями.
Таким образом, к настоящему моменту накопилось довольно много фактического материала и его исследований другими авторами, чтобы сделать анализ и обоснованные заключения об отличиях муравьев стволовых групп от современных и их причинах.
В концепции, предлагаемой мной, развиваются идеи Длусского и Федосеевой. Полагаю, что комплекс морфологических и поведенческих признаков современных муравьев сформировался в результате двух основных конструктивных требований: (I) увеличение эффективности ориентации наземного бегающего бескрылого социального насекомого (герпетобионта), (II) контроль за околоротовым пространством. К комплексу ключевых морфологических признаков я отношу: 1. латеральный поворот антенн и коленчатые антенны с длинным скапусом (более трети от длины антенны); 2. относительно длинный изогнутый педицель и увеличение размеров вершинных члеников жгутика; 3. мандибулы относительно небольшие, имеющие не менее трех зубцов. Особенности же социальности «настоящих» муравьев, по моему мнению, заключаются в: 1. коммуникации, основанной на обонятельных стимулах; 2. уходе за расплодом — облизывание и транспортировка личинок и куколок; 3. возможности трофаллаксиса (орального обмена жидкостью между членами семьи, рис. 7) (тенденция); 4. замене ядовитого жала распылительной железой (тенденция); 5. постройке и обустройстве сложного гнезда.
Как уже было сказано, Г. М. Длусский связывал коленчатость антенн с эусоциальностью из-за возможности эффективного манипулирования мелкими предметами. Однако в рамках этой концепции сложно объяснить дальнейшее удлинение скапуса у муравьев (у Formicinae, Myrmicinae, Dolichoderinae длина скапуса составляет более 0,4 длины всей антенны) и латеральный поворот антенн, поскольку пчелы и осы успешно справляются с задачами манипуляции мелкими предметами со скапусом меньшей длины и без поворота антенн, а с другой стороны, коленчатые антенны встречаются и у несоциальных наездников, например у Anastatus sp. (Eupelmidae), паразитирующих на яйцах насекомых.
Полагаю, что дальнейшее увеличение относительной длины скапуса и поворот антенн потребовались для повышения эффективности ориентирования в пространстве быстрому нелетающему социальному хищнику. Одной из причин этого мог стать находящийся в педицеле Джонстонов орган, являющийся мультисенсорным организатором — у муравьев осуществляет функции ветрового компаса, восприятия гравитации, шагового интегратора, частично отобрав функции ориентирования у зрительного органа. С инженерной точки зрения, анализировать данные с таких рецепторов тем удобнее, чем дальше друг от друга они находятся. Компромисс между необходимостью разместить анализаторы как можно дальше друг от друга, но при этом полностью контролировать пространство непосредственно около ротового отверстия и мандибул привел к 1) латеральному повороту антенн, 2) удлинению скапуса и 3) изменению формы педицеля — удлиненный изогнутый в основании членик позволяет жгутику максимально приблизиться к скапусу и, следовательно, к мандибулам и ротовому отверстию.
Четвертая особенность антенн муравьев в том, что вершинные членики жгутика увеличиваются в относительных размерах, так что вершинный членик самый большой (и по толщине, и по длине), а иногда последние членики жгутика могут формировать булаву. На мой взгляд, это строение объясняется теми же причинами — повышение эффективности пространственной ориентации, в данном случае за счет увеличения роли обонятельных анализаторов с меньшим по отношению к летающим перепончатокрылым вкладом зрения, уход за потомством, а впоследствии и коммуникация посредством обоняния. В связи с удлинением скапуса это приводит к увеличению числа и концентрации обонятельных рецепторов на вершинных члениках жгутика. О решающем значении обонятельных рецепторов для социального образа жизни в филогенетической ветви муравьев в сравнении с другими насекомыми, в том числе перепончатокрылыми, говорят результаты исследований мозга и геномов муравьев. Эволюция коммуникативных возможностей способствует появлению высших форм трофаллаксиса (в широком смысле это не только собственно обмен жидкой пищей, но и ее сбор, «содержание» выделяющих падь насекомых; см. рис. 7), что расширяет экологические потенции видов и способствует поддержанию высокой численности семей.

Уникальное разнообразие челюстей муравьев из бирманского янтаря, как было неоднократно показано, выходит за рамки существующего морфопространства рецентных муравьев и вызывает интерес не только со стороны морфологии, но и с точки зрения экологических и этологических предпосылок его возникновения. Исследователи меловых муравьев предположили, что пищевая специализация этих хищных насекомых явилась причиной подобного морфологического разнообразия, однако не ответили на вопрос: по какой причине именно такое устройство головы и челюстей «предпочитают» адские муравьи, неоднократно его воспроизводя?

Предрасположенность к эволюции в этом направлении у меловых муравьев стволовых таксонов заложена, как мне представляется, несколькими морфологическими особенностями, а именно двузубыми мандибулами и иной кранио-мандибулярной системой. Двузубые мандибулы перепончатокрылых фиксируют жертву и не позволяют ей вертеться вокруг оси в момент нанесения удара жалом. Этого достаточно для летающих хищников, даже охотящихся на крупную добычу: жертва после ужаления неподвижна и во время транспортировки по воздуху зафиксирована (поведение Ammophila, например; рис. 8).
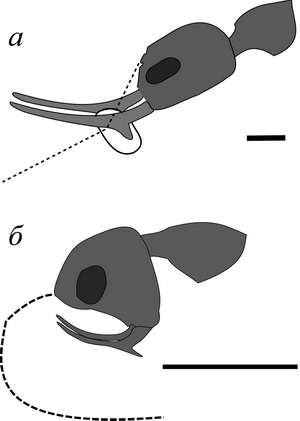
Но нелетающему социальному хищнику требуется донести до гнезда по субстрату добычу (т.е. если добыча крупная, то фактически волоком), которую в пути еще могут отобрать конкуренты. Решение, которое реализуют сфекомирмины и другие стволовые таксоны — это прижать добычу к головной капсуле (лоб, наличник), в отличие от рецентных муравьев, которые зажимают предметы между мандибулами. Таким образом, у айдамирмексов, например, с увеличением размеров добычи вершинный зубец мандибул удлиняется, лобное пространство увеличивается и армируется — добыча помещается не между челюстями, а между головой (наличником) и челюстями (см. заметку «Охота адского муравья»). Вершинный зубец мандибул увеличивается, а базальный остается ближе к основанию мандибул и ротовому отверстию и используется для более тонких манипуляций с расплодом, разделки добычи в гнезде или при обустройстве гнезда (рис. 9). Муравьи — железные девы организуют частокол зубцов, наподобие пасти некоторых рыб, на наличнике и мандибулах, которые действуют аналогично верхней и нижней челюстям рыб (см. рис. 3). Таким образом, большие или необычные «зубастые» челюсти нужны в большей степени не как орудия убийства, а в качестве средства зафиксировать жертву для удара жалом и затем донести добычу до гнезда.

Кроновые таксоны пошли по другому пути — манипуляция предметами только между мандибулами, не привлекая наличник, — и добились успеха, варьируя размеры, форму, число зубцов мандибул с одной стороны и объем и группы мышц кранио-мандибулярной системы с другой. О большом значении контроля за движениями мандибул у современных муравьев говорит исключительное развитие у них подглоточного нервного узла, осуществляющего этот контроль. Существенное преимущество современных муравьев — это коллективная охота, когда и защиту добычи, и ее транспортировку могут осуществлять очень маленькие особи с небольшими челюстями вне зависимости от размера добычи (рис. 10).
Габитус (внешний вид) эффективного наземного быстрого нелетающего хищника-герпетобионта (дендробионта) и социальное устройство, основанное на запаховых стимулах, способствовали процессу совершенствования новой жизненной формы. В этот момент, видимо, происходит радиация современных таксонов по Г. М. Длусскому, т.е. приспособление новой (конструктивной) формы к разным ярусам с вытеснением стволовых таксонов. Представители стволовых таксонов муравьев обладали менее совершенной коммуникативной способностью, менее эффективной (просто другой) кранио-мандибулярной системой и зрительной пространственной ориентацией в большей степени, чем обонятельной, что блокировало развитие социальности, основанной на запаховых стимулах.
Таким образом, экологическая роль и у вымерших стволовых таксонов, и у появившихся приблизительно в то же время современных муравьев была одна и та же, но конструктивные особенности позволили «настоящим» муравьям вытеснить меловую мирмекофауну. Несмотря на специализацию, муравьи стволовых таксонов не могут удержать позиций, поскольку совершенные социальные навыки кроновых групп муравьев способствуют более эффективному удержанию пищевых ресурсов. В созданном стволовыми таксонами муравьев адаптивном пространстве поделенные специалистами по размерам и типам добычи экологические ниши по-новому «форматируются» представителями современных групп муравьев — возникает система соподчинения, понимаемая у современных муравьев под доминированием (виды-доминанты, субдоминанты, инфлюенты), где тип и размеры добычи на видовом уровне определяются не размером челюстей, а численностью семьи.
Популярные синопсисы