«Истинный творец всего». Отрывок из книги

История развития человеческого вида через призму Истинного творца всего — человеческого мозга, который на всем протяжении нашего длинного эволюционного пути придавал смысл окружающему нас миру, наделял вещи значениями и сам изменялся в зависимости от получаемой информации, позволяя нам адаптироваться к переменчивым условиям и выживать несмотря ни на что. …
Глава 4. Динамическая подпитка мозга. Биологические соленоиды и принципы функционирования (отрывок)
Уже 100 тысяч лет назад нервная система каждого человека могла похвастаться 86 миллиардами органических процессоров (нейронов), между которыми могло существовать от 100 триллионов до квадрильона прямых контактов (синапсов). В этой невероятной нейронной мастерской Истинный творец всего начал свою работу над человеческой вселенной в том виде, в котором мы знаем ее сегодня.
Масса коры мозга, об эволюции которой мы говорили выше, составляет около 82% массы всего человеческого мозга. Удивительно, но на нее приходится лишь 19% (около 16 миллиардов) нейронов мозга. Для сравнения в человеческом мозжечке — важнейшем скоплении серого вещества, отвечающем за контроль двигательной функции, на долю которого приходится лишь 10% массы мозга, — упаковано около 69 миллиардов нейронов, так что это очень плотный кластер нейронов. Однако мозжечок, насколько мы можем судить, не сочинял сонетов и пьес Шекспира и не проектировал космических кораблей, позволяющих нам осваивать космическое пространство (хотя он помогал их строить). Вот почему с этого момента, говоря о том, каким образом Истинный творец всего справляется со своими самыми сложными задачами, мы будем в основном обращать внимание на новую кору.
В оптимизации работы коры важнейшую роль играет сложная сеть белого вещества. Несколько плотных упаковок нервных волокон (рис. 1) белого вещества образуют петли, связывающие между собой скопления серого вещества. Я называю эти петли биологическими соленоидами — по аналогии с катушками в электромагнитах. Самой крупной из этих биологических катушек является мозолистое тело.

Мозолистое тело — это толстый слой ткани примерно из 200 миллионов нервов, расположенный вдоль продольной оси головного мозга, который обеспечивает обмен информацией между двумя полушариями мозга и координацию их активности. Между задней и передней частями мозолистого тела коры имеются значительные структурные различия, включающие в себя среди прочего плотность и диаметр аксонов, проводящих электрические импульсы (так называемые потенциалы действия), а также степень миелинизации аксонов. Особый тип поддерживающих клеток мозга образует вокруг нервных волокон слой миелина. Обертывание нервных волокон слоем миелина обеспечивает крайне высокую скорость передачи потенциала действия миелинизированными аксонами. Как следствие, миелинизированные нервы затрачивают на процесс передачи меньше энергии. Например, если немиелинизированное нервное волокно группы C диаметром 0,2–1,5 мкм проводит потенциал действия примерно со скоростью 1 м/с, в крупном миелинизированном волокне такой же электрический импульс перемещается со скоростью около 120 м/с, или более 400 км/ч. Таким образом, время передачи информации между полушариями по всей длине волокна весьма разнится в зависимости от того, из какого участка коры поступают сигналы. В целом эти различия в скорости передачи сигнала статистически описываются распределением в форме широкой колоколообразной кривой. Например, в соответствии с этим распределением обмен информацией между полушариями в моторных и сенсорных областях происходит очень быстро, поскольку они соединены толстыми миелинизированными аксонами мозолистого тела. Напротив, связь между так называемыми ассоциативными зонами в лобной и теменной долях осуществляется намного медленнее.
Мы точно не знаем, каким образом 200 миллионов волокон мозолистого тела координируют работу двух полушарий мозга. Однако мы знаем, что мозолистое тело действительно их синхронизирует, поскольку при его удалении полушария начинают действовать независимо. Глубокие исследования пациентов с так называемым расщеплением мозга начались много десятилетий назад, когда соответствующий тип хирургического вмешательства стали использовать для предотвращения распространения серьезных нарушений с одного полушария мозга на другое. В 1981 году американский нейробиолог Роджер Сперри был удостоен Нобелевской премии по медицине* за прорыв в изучении пациентов с расщепленным мозгом и функции мозолистого тела.
У большинства людей некоторые ключевые функции мозга, такие как речь, латерализованы в коре, т. е. осуществляются в основном в одном из полушарий (так, у правшей за речь отвечает левое). В результате латерализации пациенты с расщепленным мозгом не всегда могут описать словами то, что они видят. Например, если какое-то изображение находится в левой части их поля зрения или если их просят держать левой рукой предмет, который они не видят, они просто не могут назвать или описать этот предмет или изображение. И дело не в том, что они не знают ответа на вопрос. Знают. Проблема в том, что стимулы, поступающие с левой стороны, обрабатываются правой частью мозга. Поскольку мозолистое тело отсутствует, правое полушарие не способно сообщаться с речевой зоной левого. На самом деле пациенты с расщеплением мозга могут левой рукой выбрать из набора предметов один предмет, который идентичен тому, что они держали за минуту до этого; они осознают, что видят и до чего дотрагиваются. Но они не могут об этом рассказать.
Внутри каждого полушария находится множество других крупных петель и пучков белого вещества, связывающих между собой разные области коры. Одна из таких систем, обеспечивающая важную связь между лобной, теменной и височной долями, образована тремя плотно упакованными нервными пучками с очень высокой проводимостью. Первый из них называется крайней (наружной) капсулой и обеспечивает связь между ключевыми участками височной доли (например, находящимися в верхней височной борозде, sulcus temporalis superior, STS, и нижней височной зоне) и нижней частью префронтальной коры. Вторая система, связывающая STS с участком теменной коры, образована так называемыми медиальными и задними продольными пучками. Наконец, существует верхний продольный пучок, осуществляющий связь между теменной и лобной долями. Вместе эти три пути вовлечены в обеспечение таких ключевых функций, как речь, изготовление орудий и мимикрия движений.
Еще один важнейший скоростной коммуникационный путь мозга — кортико-таламо-кортикальная петля, которая обеспечивает взаимодействие между корой и таламусом — важнейшей субкортикальной структурой, получающей основной поток сенсорных данных от периферических нервов. Поэтому этот мультимодальный сенсорный путь является важнейшим элементом в механизме непрерывного сравнения данных между уже имеющимися у мозга данными и набором сырой информации, поступающей из внешнего мира. Эта петля также играет важную роль в синхронизации электрической активности коры и таламуса.
Еще одна важная особенность человеческого белого вещества заключается в его развитии. По сравнению с мозгом наших родственников шимпанзе человеческий мозг на момент рождения организма еще сравнительно неразвит и достигает зрелости только через два десятилетия. Кроме того, хоть мы и рождаемся примерно с тем количеством нейронов, которое будем иметь на протяжении всей жизни, на пик своего функционирования белое вещество выходит лишь через тридцать или даже сорок лет. В частности, в префронтальной зоне лобной доли связи между нейронами (как синапсы, проводящие потенциалы действия между нейронами, так и дендриты, принимающие эти сообщения) достигают полной зрелости только на третьем десятке жизни. Все это означает, что процесс увеличения объема мозга после рождения связан с разрастанием и усложнением белого вещества. Этот длительный процесс созревания (и возможность его нарушения) объясняет подверженность человека ментальным нарушениям, таким как шизофрения и аутизм, в детские годы и в подростковом возрасте. Отсроченное созревание белого вещества также помогает объяснить изменения поведения и мыслительных функций, которые все мы переживаем в первые десятилетия жизни. Поэтому, когда в следующий раз будете иметь «дружескую» дискуссию со своим бунтующим ребенком-подростком, просто сделайте глубокий вдох и вините вместо него во всем медленное созревание белого вещества!
Одно из самых замечательных открытий в области изучения мозга за последние пятьдесят лет было сделано группой нейробиологов под руководством Джона Кааса из Университета Вандербильта и Майкла Мерзенича из Университета Калифорнии в Сан-Франциско, которые в начале 1980-х годов убедительно показали, что сложные сети нейронов, определяющие функцию мозга млекопитающих и приматов, находятся в постоянном динамическом развитии на протяжении всей жизни. Наш мозг изменяет сам себя, как в анатомическом, так и в физиологическом плане, в ответ на все и на всех, с кем мы взаимодействуем по мере получения новых навыков, и даже когда в нашем теле или вокруг нас происходят значительные изменения. Нейробиологи называют это свойство пластичностью мозга, и оно является важнейшим ключом в раскрытии глубочайших тайн Истинного творца всего.
Изменение нейропластичности на уровне синапсов происходит несколькими путями. Например, количество и распределение синапсов может значительно изменяться при обучении новому навыку или в ходе восстановления после повреждения периферических частей тела или самого мозга. Даже у взрослых животных некоторые нейроны могут создавать новые синапсы, что способствует укреплению связи с отдельными или со всеми целевыми нейронами. Возможен и обратный процесс, когда одни нейроны ликвидируют синапсы, тем самым ослабляя связь с другими. Степень влияния каждого синапса на конкретный нейрон также может значительно изменяться в зависимости от того, что происходит с нашим мозгом. Фактически любой стимул может изменить тонкую микроструктуру и функцию сотен триллионов синаптических связей, посредством которых осуществляется контакт между десятками миллиардов нейронов коры.
Посвятив более десяти лет изучению пластичности мозга, летом 2005 года я предложил нейробиологу Эрику Томсону, работавшему в моей лаборатории в Университете Дьюка, весьма неортодоксальный способ попытаться понять, насколько далеко распространяется данное явление. Мы разработали эксперимент со взрослыми крысами, с помощью которого хотели проверить, поможет ли максимальная степень пластичности приобрести совершенно новые ощущения в дополнение к традиционным, с которыми животные родились (тактильным, зрительным, слуховым, вкусовым, обонятельным, вестибулярным). Мы решили попробовать научить крыс «дотрагиваться» до невидимого инфракрасного света. Для этого нам потребовалось создать устройство, превращающее инфракрасное излучение от внешнего источника в поток электрических импульсов (именно этот язык мозг использует для передачи сообщений), которые дальше могут передаваться в первичную соматосенсорную кору животного — основную зону, ответственную за создание тактильных ощущений у млекопитающих. Направляя эти новые электрические стимулы в первичную соматосенсорную кору, мы хотели понять, смогут ли наши «киберкрысы» научиться обрабатывать инфракрасное излучение в качестве дополнительного элемента тактильного восприятия.
С этой целью Эрик сконструировал устройства, состоящие из нескольких инфракрасных сенсоров (от одного до четырех), и разместил их на голове у крыс (рис. 2). Каждый сенсор фиксировал инфракрасное излучение в пространственном секторе охватом около 90 градусов, что означает, что устройство с четырьмя сенсорами обеспечивало крысе полный обзор окружающего пространства в инфракрасном спектре. В соматосенсорной коре нашей целью был участок, называемый бочонком, который обрабатывает поступающие тактильные сигналы, возникающие при стимуляции усов крысы. Усы крыс, подобно кончикам пальцев у приматов, являются самыми чувствительными тактильными органами, а потому значительная площадь соматосенсорной коры мозга крыс вовлечена в обработку тактильных сигналов, создаваемых этими волосками на мордочке.

Рис. 2. Первая конфигурация инфракрасного нейропротеза, использованного Эриком Томсоном в экспериментах в нашей лаборатории. А: Схема камеры, в которой фиксировалось поведение крыс при решении задач на дискриминацию инфракрасного (IR) света. На внутренней поверхности большого (60 см) цилиндра симметрично установлены четыре порта с отверстиями для носа, источником IR и видимого света. B: Топографическое расположение четырех имплантатов в первичной соматосенсорной коре мозга крысы (S1), проводящих электрические сигналы от четырех детекторов IR. Сенсоры IR расположены под прямым углом по отношению друг к другу, каждый совмещен со своей парой стимулирующих электродов в S1. C: Частота стимуляции зависит от интенсивности IR в каждом сенсоре. Интенсивность каждого IR луча конвертируется в реальном времени в сигнал с разной частотой стимуляции в соответствующем канале. D: График в полярных координатах, отражающий ответ каждого сенсора IR в зависимости от угла, когда сенсорная антенна находится в фиксированной позиции по отношению к единственному активированному источнику IR. Точка на окружности (вверху справа) указывает относительное расположение источника IR. E: Профиль ответа, выраженного как полная ширина на половине высоты (FWHM), в зависимости от положения в камере. Черная точка соответствует положению активного источника IR, а FWHM — среднее значение FWHM для всех четырех сенсоров в данном положении (см. D). Если удаляться от источника или перемещаться вбок, профиль ответа сужается. Черная точка обозначает ситуацию, представленную на рисунке D. Hartmann K. et al. Embedding a Novel Representation of Infrared Light in the Adult Rat Somatosensory Cortex through a Sensory Neuroprosthesis. Journal of Neuroscience 36, no. 8, February 2016: 2406–24.
Мы начали эксперимент с того, что стали обучать крыс следить за пучком видимого света, подводившего их к лакомству. Когда они научились решать эту базовую задачу, мы подключили инфракрасные сенсоры Эрика, чтобы понять, смогут ли они находить угощение, фиксируя и отслеживая пучок инфракрасного излучения путем прикосновения. Для этого Эрик установил на внутренней поверхности круглой камеры, где во время эксперимента находились «киберкрысы», источники инфракрасного излучения в позициях 0, 90, 180 и 270°. Положение излучателей позволяло нам выборочным образом изменять источник луча во время эксперимента, чтобы мы могли быть уверены в том, что крысы находят угощение не с помощью обычных органов чувств. Поначалу мы встраивали крысам лишь один сенсор инфракрасного излучения. Животным потребовалось около четырех недель, чтобы научиться успешно «дотрагиваться» до инфракрасного луча и следовать вдоль него в поисках лакомства более чем в 90% экспериментов.
В первых экспериментах наши «киберкрысы» демонстрировали очень интересные особенности поведения: поначалу они крутили головой в горизонтальной плоскости, как будто сканировали пространство вокруг себя в поисках сигнала; при появлении инфракрасного луча крысы всегда терли мордочку передними лапами, прежде чем начинали следовать за лучом в сторону конкретного излучателя. Хотя первое наблюдение показало, что крысы разработали собственную стратегию для обнаружения первых признаков появления инфракрасного луча, второе говорило скорее о том, что они чувствовали инфракрасное излучение, как будто их усики касались чего-то во внешнем пространстве. Однако на деле они ни до чего не дотрагивались. Это мозг крыс обучался обрабатывать поступающий сигнал инфракрасного света в качестве некоего действующего на усики тактильного стимула!
Хотя эти результаты уже были весьма обнадеживающими, самый большой сюрприз ждал нас чуть позже, когда Эрик начал анализировать записи электрической активности отдельных нейронов в соматосенсорной коре мозга наших крыс, следивших за инфракрасным излучением. Значительная доля их нейронов, которые раньше возбуждались только тогда, когда животные касались чего-либо усиками, теперь приобрели способность реагировать на присутствие в окружающей среде инфракрасного излучения (рис. 3).
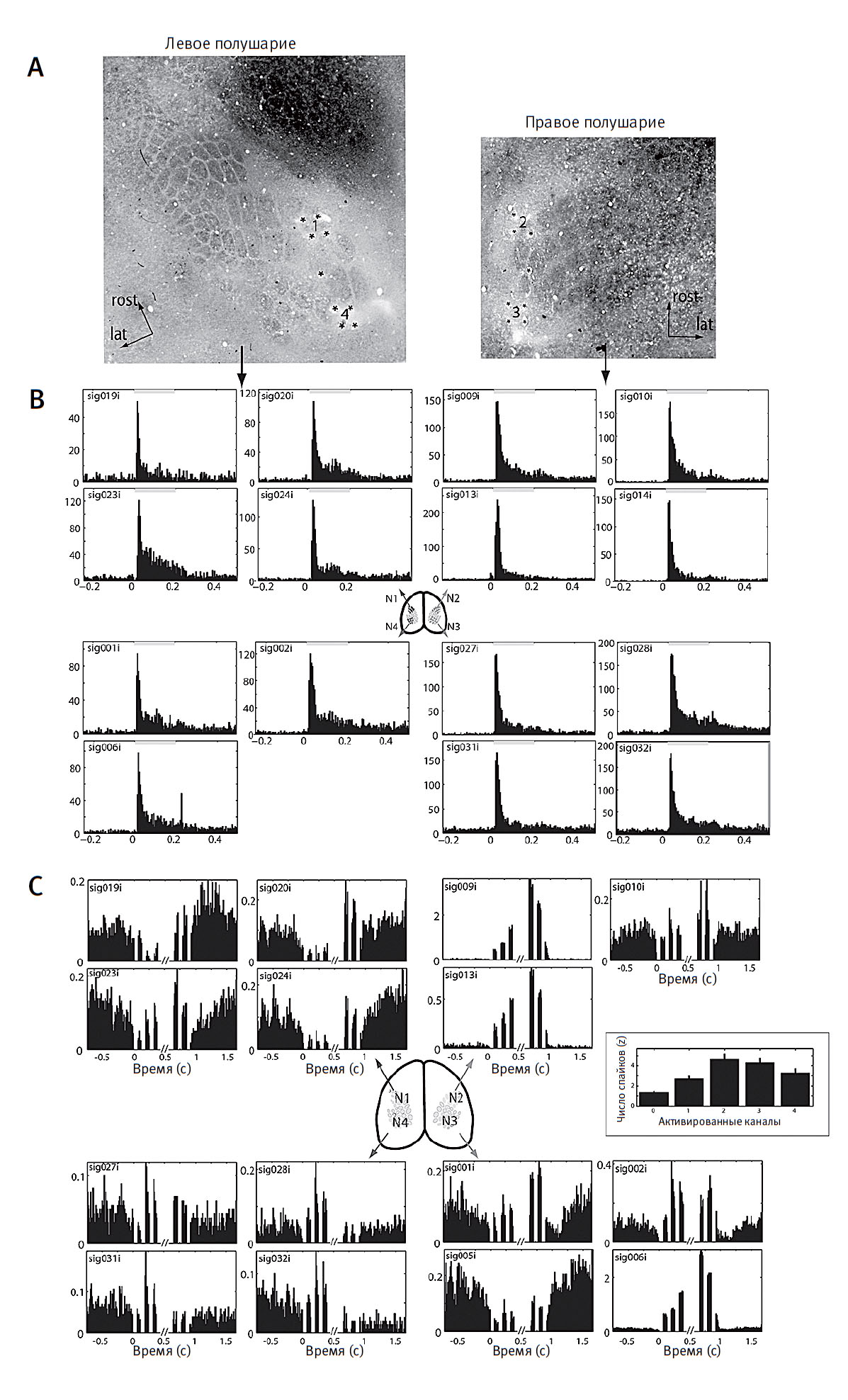
Рис. 3. Отдельные нейроны соматосенсорной коры (S1, A) отвечают и на механическую стимуляцию усиков на мордочке животного (верхняя полоса, B), и на IR-излучение в случае крыс с имплантированным нейропротезом (нижняя полоса, C), передающим электрический стимул на первичную соматосенсорную кору (S1). Верхняя полоса, A: Уплощенные срезы коры в S1 полушариях мозга одного животного демонстрируют расположение электродов. Звездочки указывают места вживления электродов. B: Очень стойкие ответы для 15 нейронов S1 у того же животного, вызванные сенсорным стимулом и сопровождающиеся механическими отклонениями усиков, проявляются в виде четких пиков электрической активности нейронов на перистимулярных временных гистограммах (PSTH). Такой тактильный ответ нейронов был получен после обучения животных распознанию IR-сигналов. Ширина столбика гистограммы PSTH — 1 мс. C: PSTH описывает электрический ответ нейронов S1 на сигналы IR-стимуляции. Стрелки указывают на расположение нейронов в коре S1. Правый график отражает z-значение для потенциалов действия как функцию числа активированных стимулирующих каналов. Это типичная картина, при которой максимальный ответ наблюдается при одновременной активации двух каналов. C модификациями из работы: Hartmann K. et al. Embedding a Novel Representation of Infrared Light in the Adult Rat Somatosensory Cortex through a Sensory Neuroprosthesis. Journal of Neuroscience 36, no. 8, February 2016: 2406–24.
В следующем эксперименте мы использовали четыре сенсора инфракрасного излучения, позволявшие получать панорамное изображение цилиндра. В этой серии экспериментов для освоения такой же задачи крысам потребовалось всего три дня, а не четыре недели. Контрольные эксперименты показали, что даже при изменении картины пространственного взаимодействия между выходами инфракрасных сенсоров и различными подобластями соматосенсорной коры мозга крысы быстро переучивались отслеживать инфракрасные лучи и успешно находить с их помощью лакомство более чем в 90% случаев.
В целом эти две группы экспериментов четко подтвердили возможность приобретения крысами нового тактильного ощущения. Примечательно, что это происходило не за счет уже существовавшего репертуара возможностей: к лету 2016 года Эрик показал, что ни одна из крыс, способных распознавать инфракрасное излучение, не утратила способности использовать усики для выполнения стандартных задач по тактильной дискриминации, которые они выполняют с таким мастерством. Иными словами, участок коры, который ранее занимался исключительно обработкой одного важнейшего типа сигнала (в данном случае тактильной информации), превратился в мультифункциональный участок мозга, хотя за долгую эволюционную историю этого живучего вида ни одна крыса никогда не ориентировалась на сигналы такого рода. В целом можно сказать, что благодаря использованию сенсорных нейропротезов головной мозг наших модифицированных крыс получил способность создавать новые изображения окружающего мира на основании инфракрасных сигналов в дополнение к уже существующему тактильному представлению.
* * *
* Одновременно с Роджером Сперри Нобелевскую премию по медицине «за открытия, касающиеся принципов переработки информации в зрительной системе», получили Дэвид Хьюбел и Торнтон Визель.






















Рис. 1. Типичные петли белого вещества коры, наблюдаемые при помощи диффузионно-тензорной томографии (изображения любезно предоставлены Алленом Сонгом).